
Functional Role of AGAP2/PIKE-A in Fcγ Receptor-Mediated Phagocytosis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Antibodies
2.3. Isolation of Human PMNs
2.4. Electrophoresis and Immunoblotting
2.5. AGAP2 Immunoprecipitation
2.6. Metabolic Labelling with 32P
2.7. Plasmid Constructs and PCR Mutagenesis
2.8. Expression and Purification of AGAP2
2.9. Cell Culture and Transfection
2.10. RT-PCR
2.11. Confocal Microscopy
2.12. Phagocytosis Assays
2.13. FACS Analysis of Phagocytosis
2.14. Statistical Analysis
3. Results
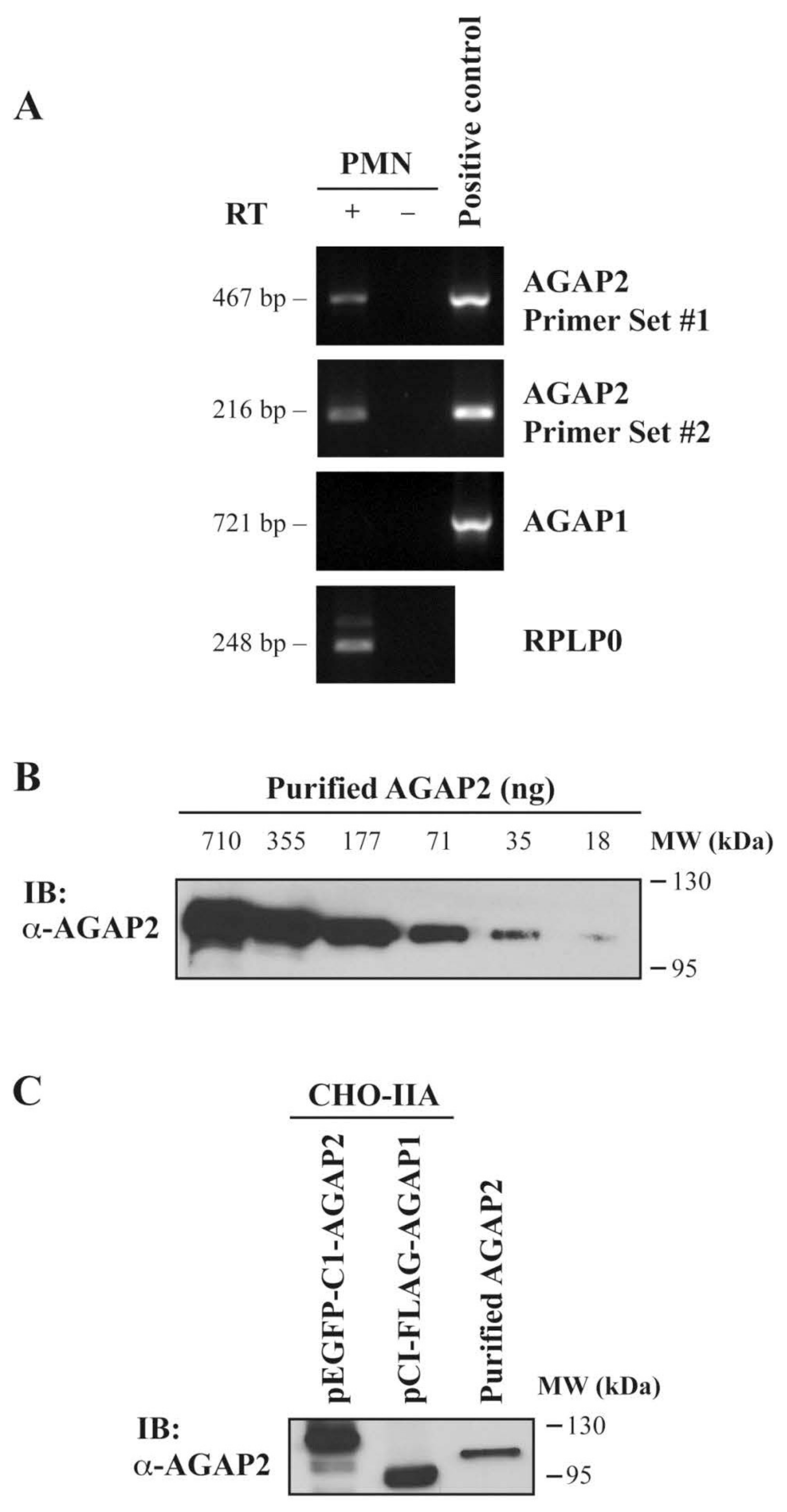
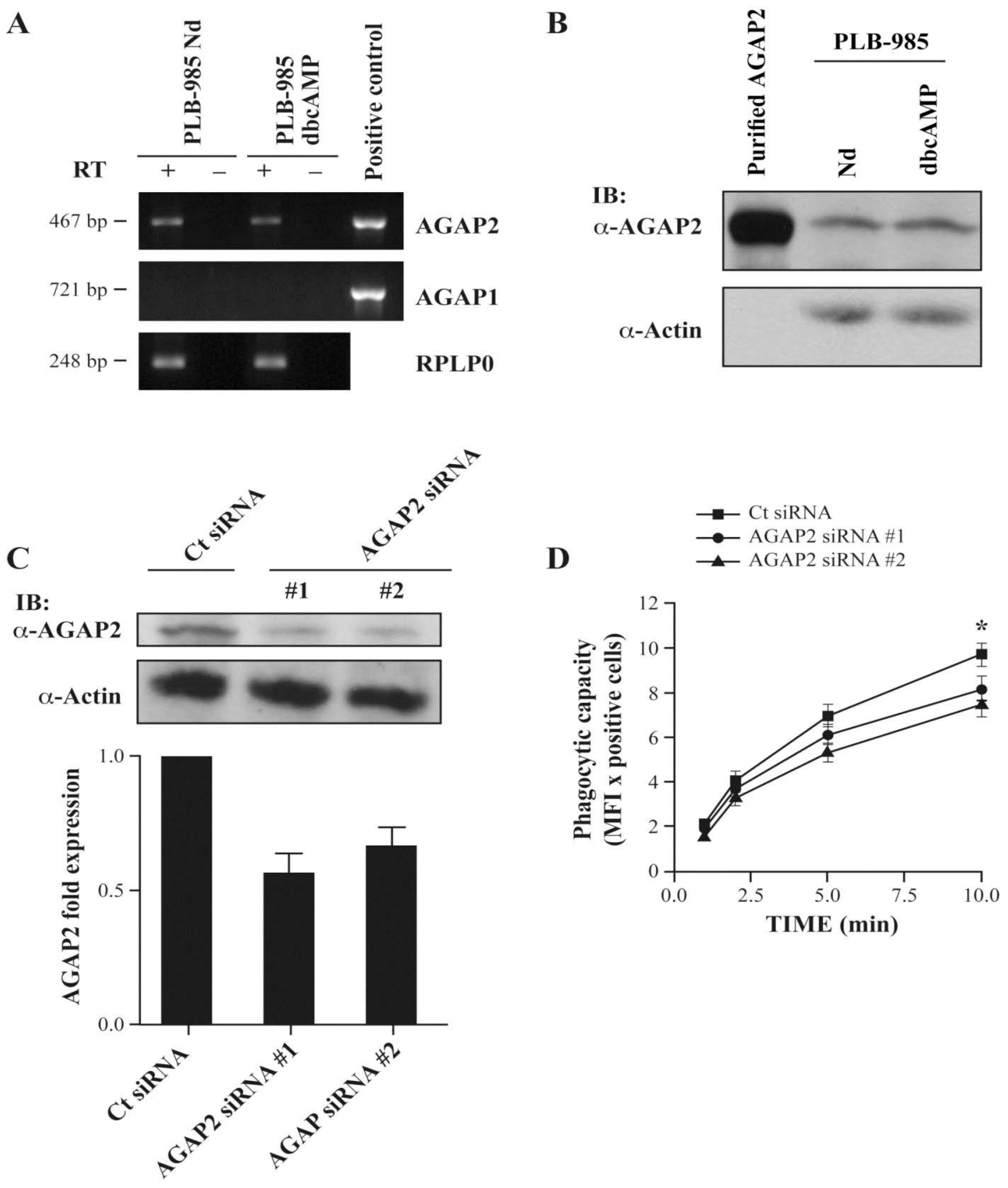
3.1. AGAP2, but Not AGAP1 or PIKE-L, Is Expressed in Human Neutrophils
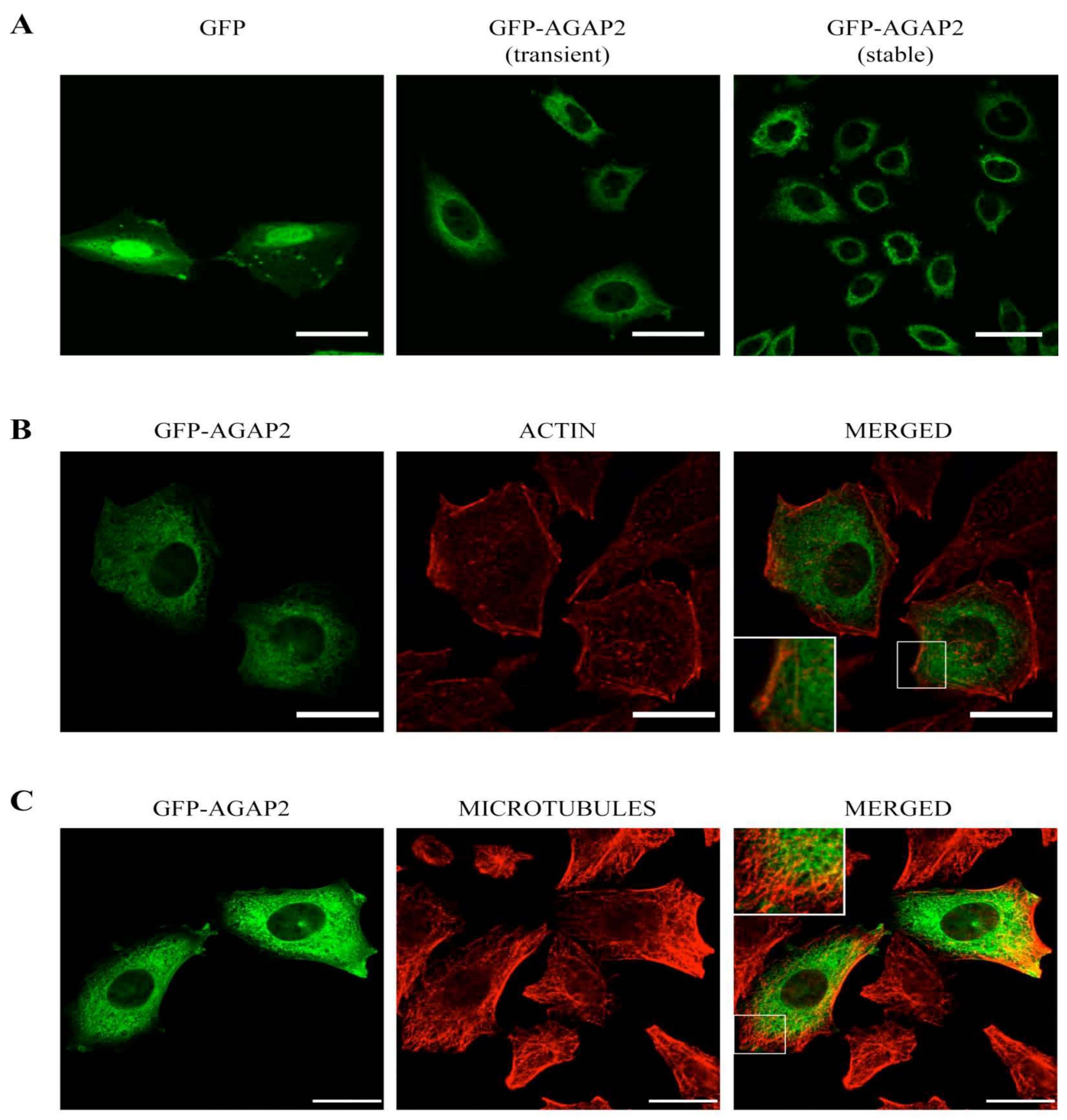
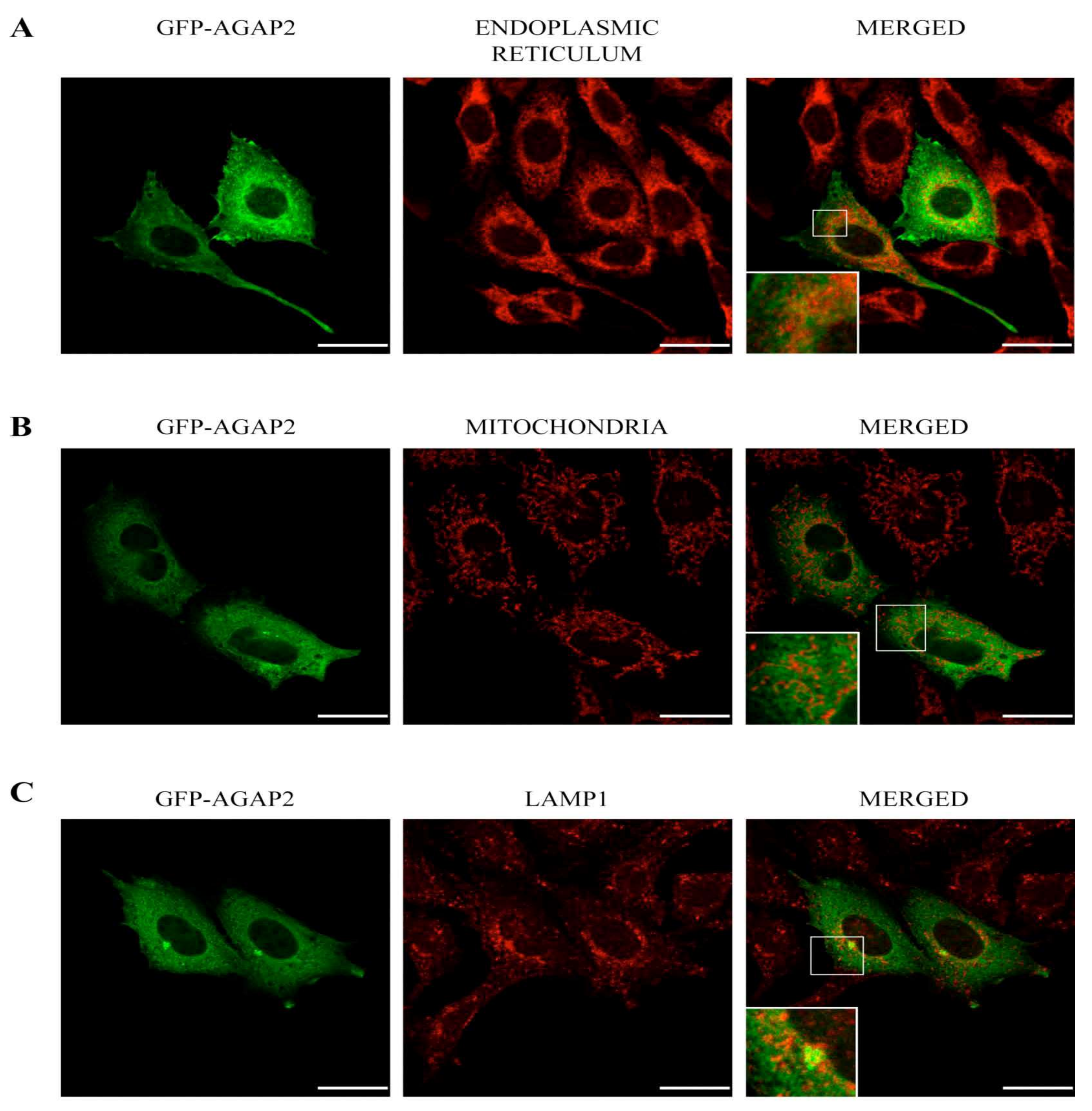
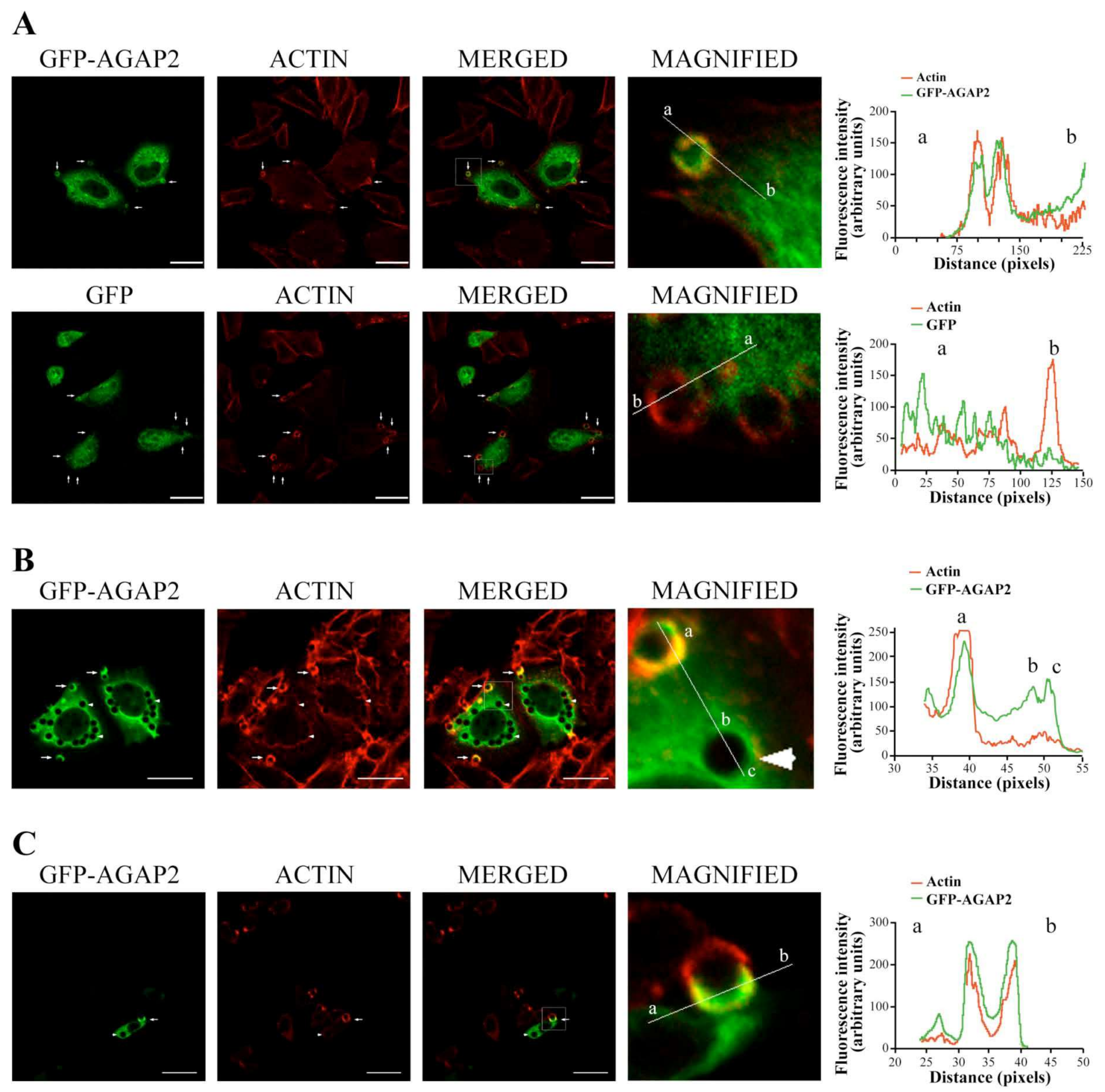
3.2. Sub-Cellular Localization of AGAP2 in CHO-IIA Cells
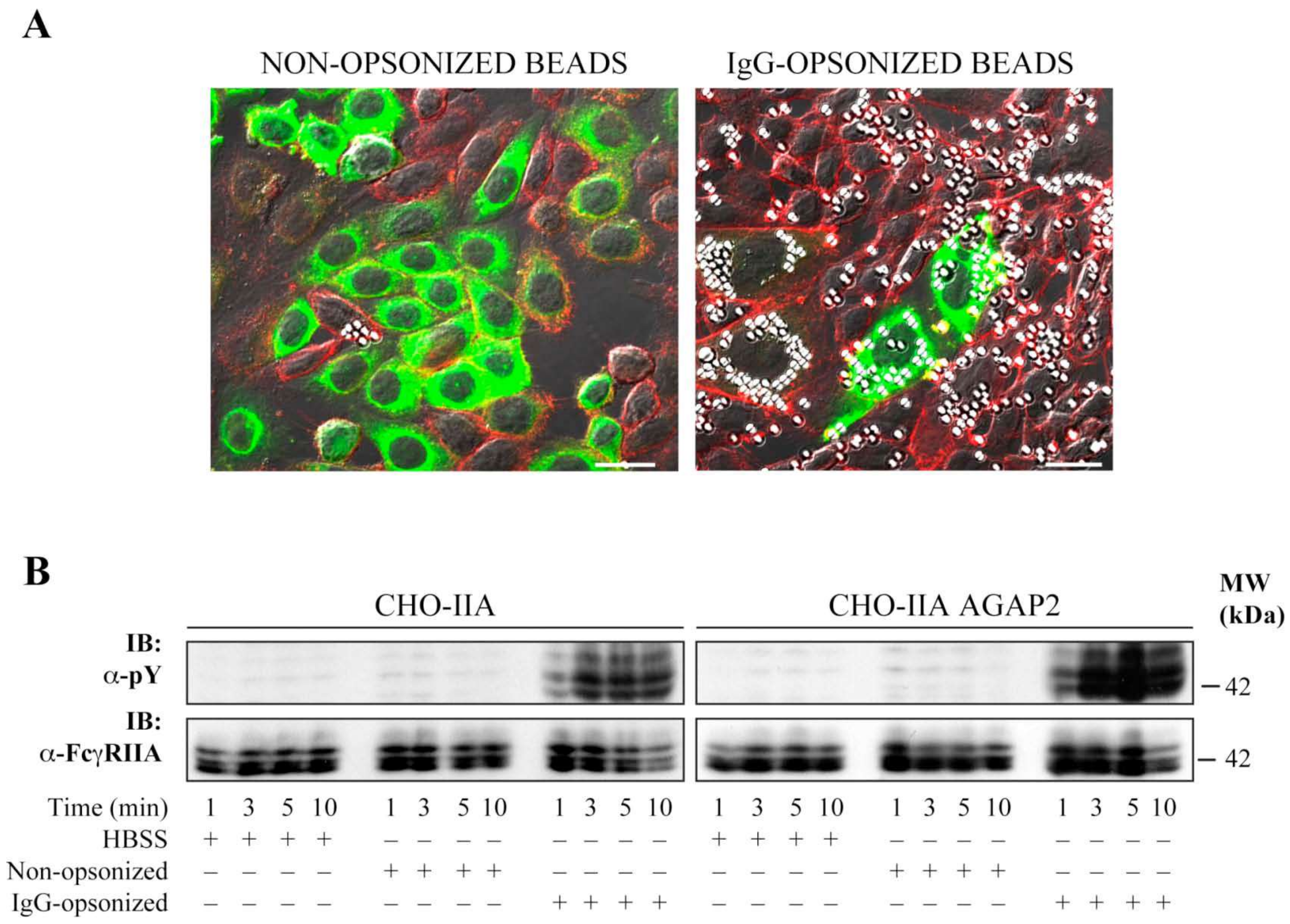
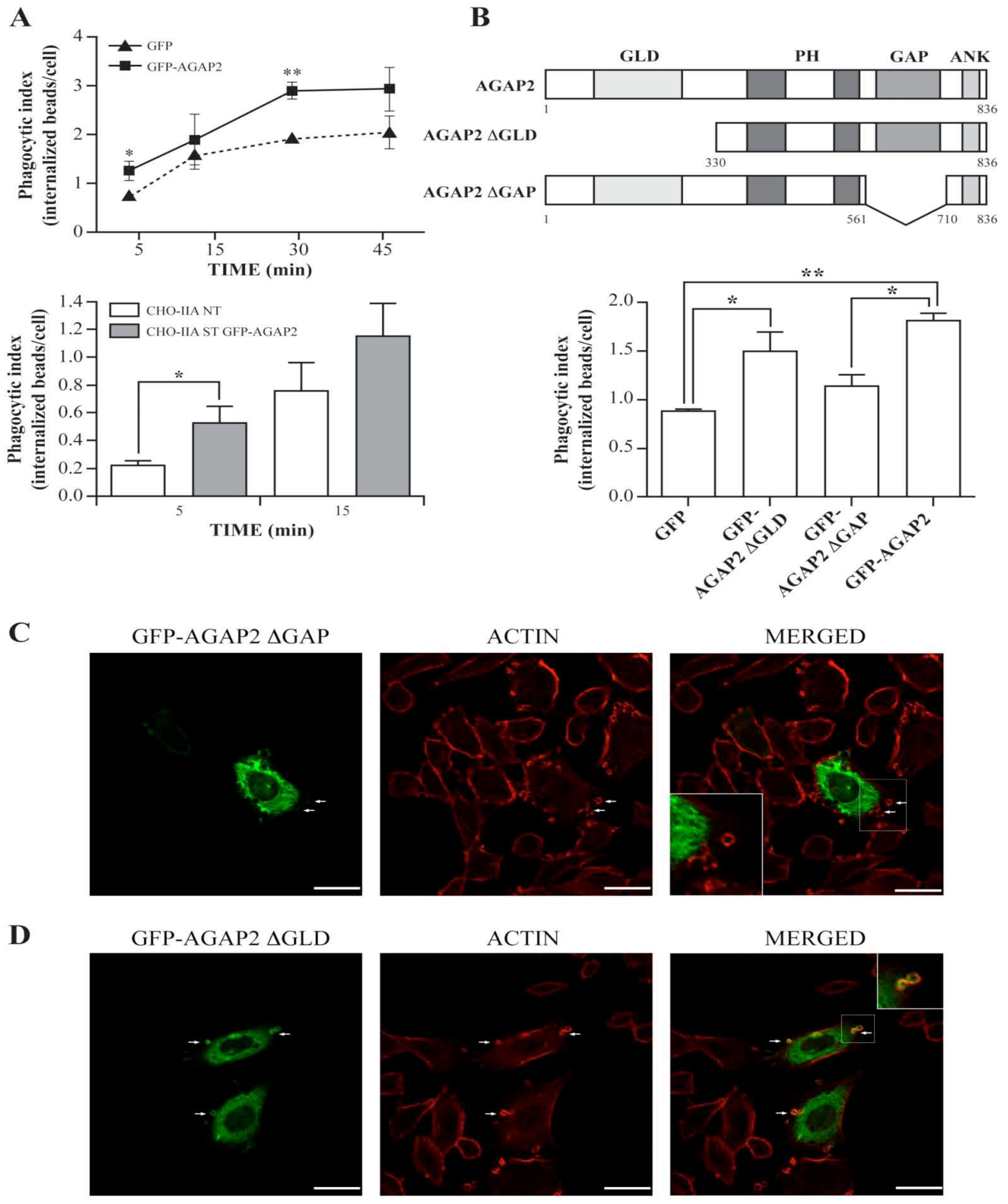
3.3. AGAP2 Is Transiently Recruited to the Phagocytic Cup during FcγR-Mediated Phagocytosis
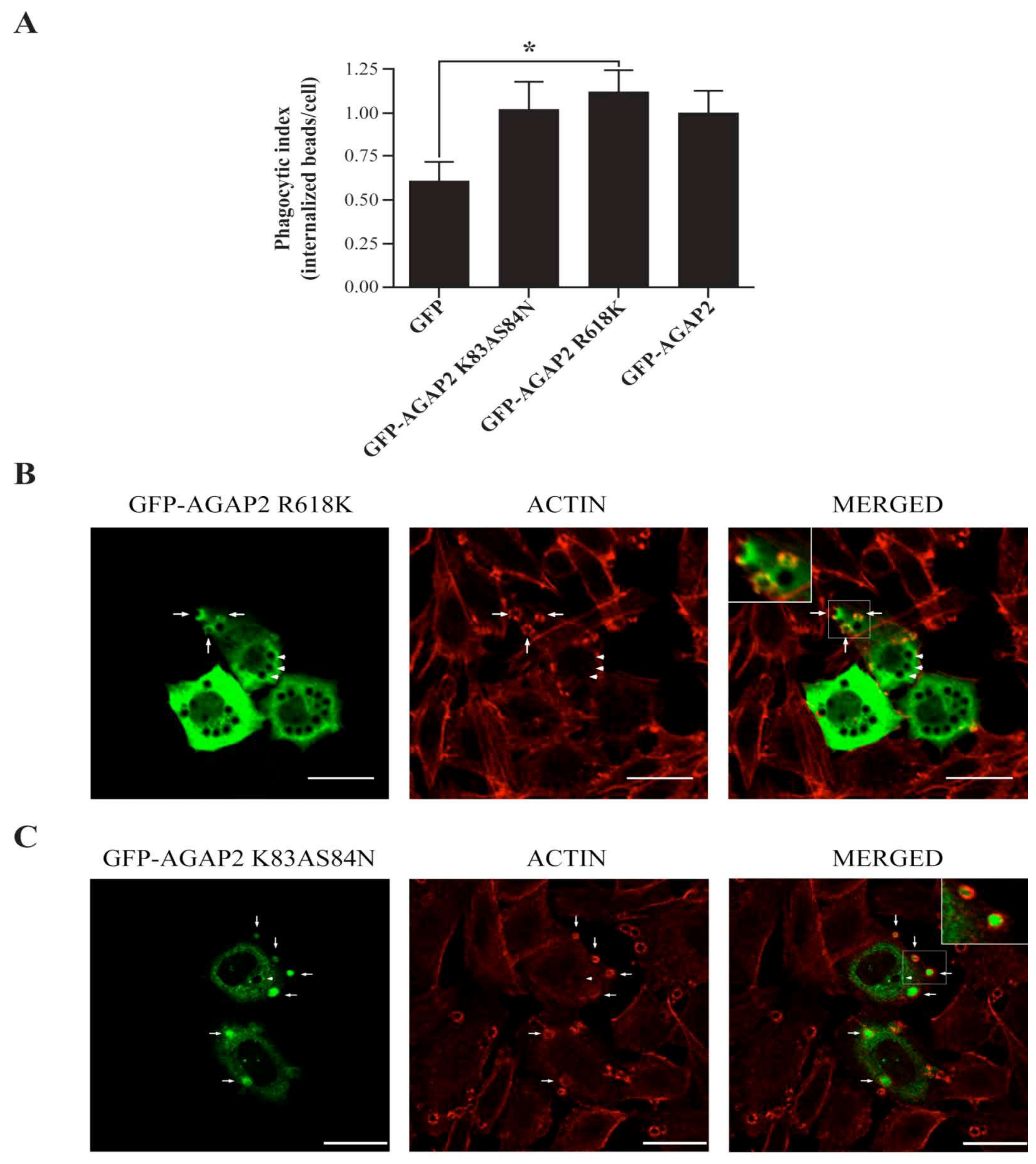
3.4. AGAP2 Increases the Phagocytic Efficiency of CHO-IIA Cells
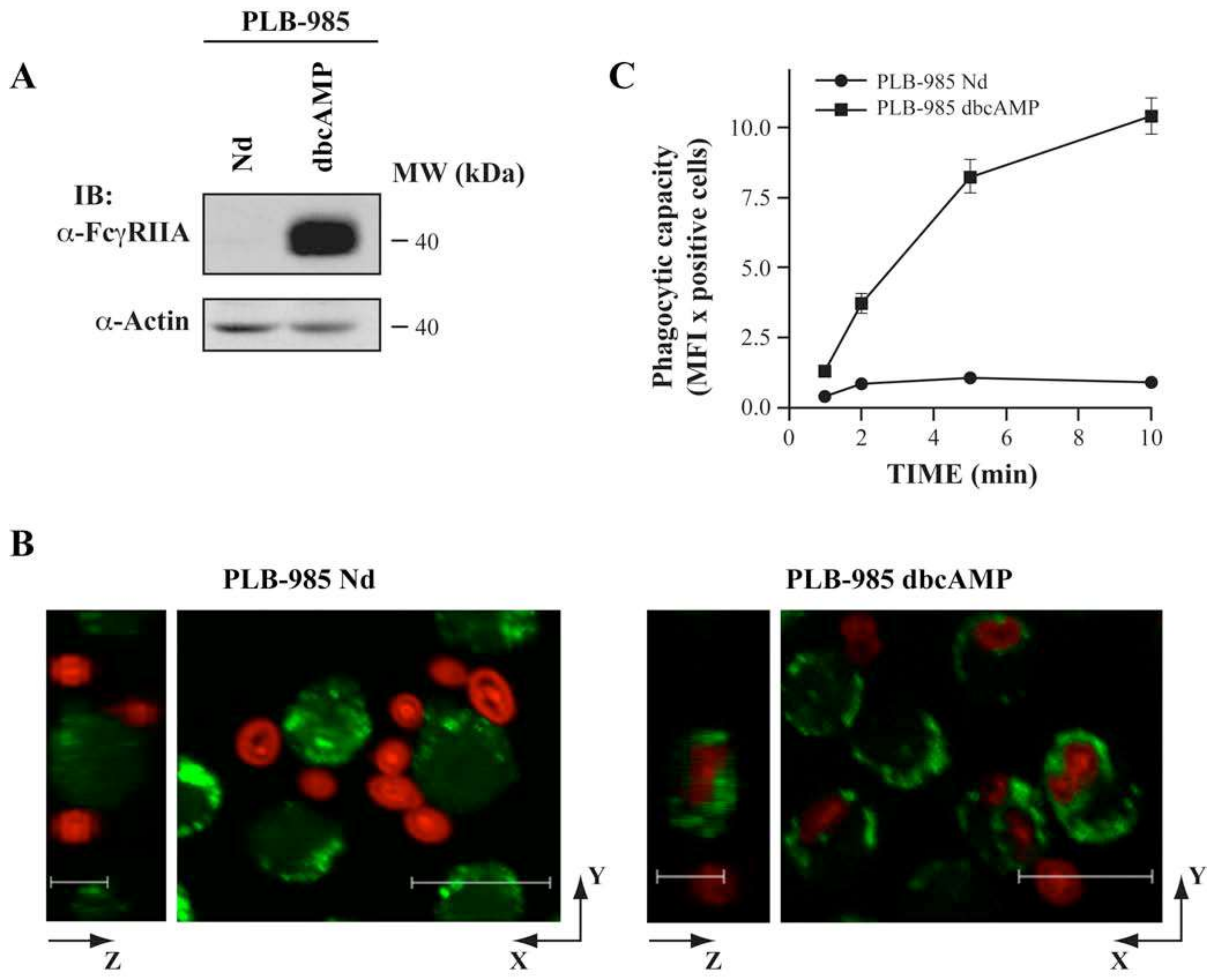
3.5. AGAP2 Silencing in the Myeloid Cell Line PLB-985 Decreases Phagocytosis
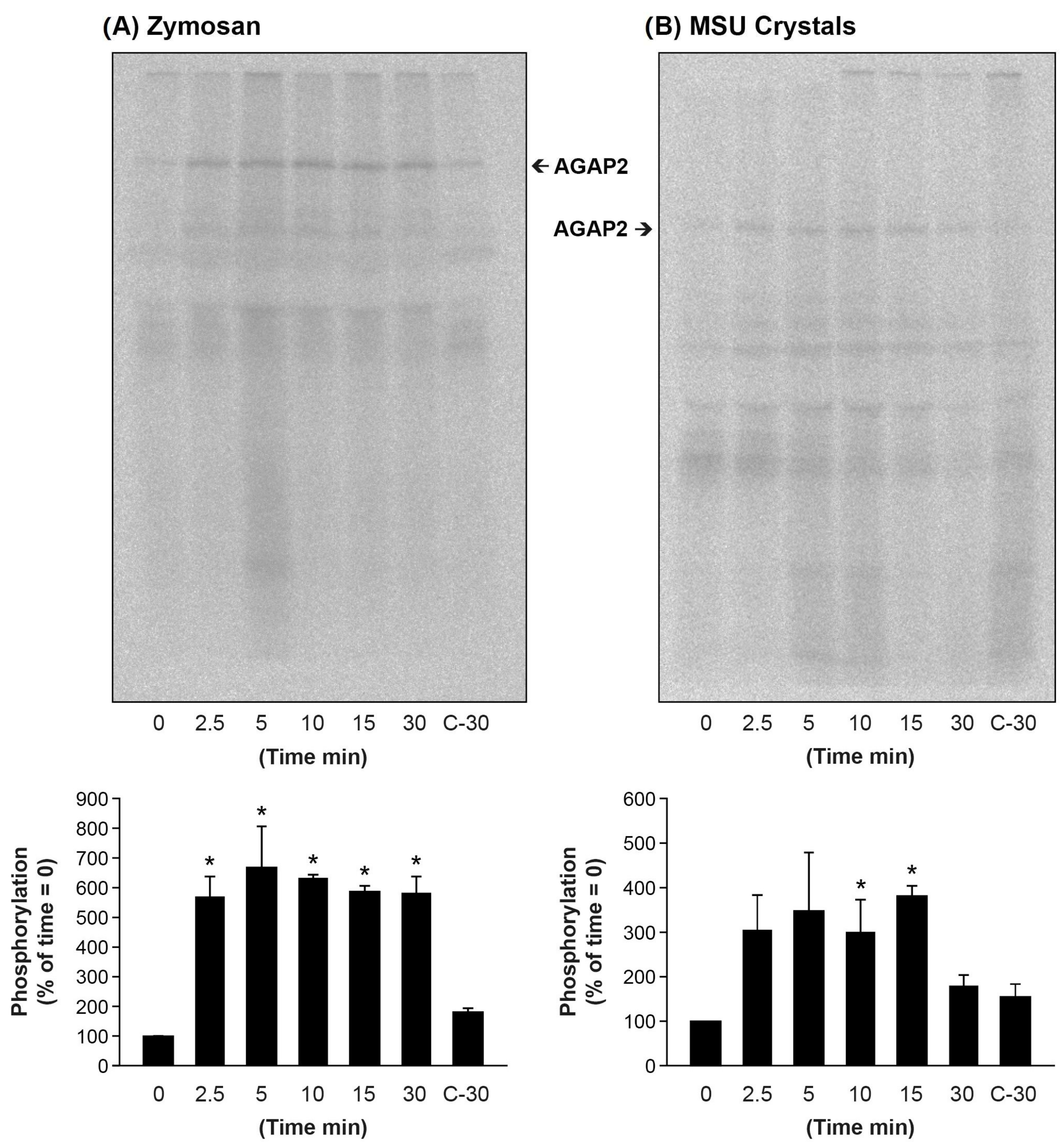
3.6. Particulate Agonists Induce AGAP2 Phosphorylation in Human PMNs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gillingham, A.K.; Munro, S. The small G proteins of the Arf family and their regulators. Annu. Rev. Cell Dev. Biol. 2007, 23, 579–611. [Google Scholar] [CrossRef]
- Sztul, E.; Chen, P.W.; Casanova, J.E.; Cherfils, J.; Decks, J.B.; Lambright, D.G.; Lee, F.J.S.; Randazzo, P.A.; Santy, L.C.; Schurmann, A.; et al. ARF GTPases and their GEFs and GAPs: Concepts and challenges. Mol. Biol. Cell 2019, 30, 1249–1271. [Google Scholar] [CrossRef] [PubMed]
- Kahn, R.A.; Bruford, E.; Inoue, H.; Logsdon, J.M.; Nie, Z.; Premont, R.T.; Randazzo, P.A.; Satake, M.; Theibert, A.B.; Zapp, M.L.; et al. Consensus nomenclature for the human ArfGAP domain-containing proteins. J. Cell Biol. 2008, 182, 1039–1044. [Google Scholar] [CrossRef]
- Randazzo, P.A.; Hirsch, D.S. Arf GAPs: Multifunctional proteins that regulate membrane traffic and actin remodelling. Cell Signal. 2004, 16, 401–413. [Google Scholar] [CrossRef]
- Nie, Z.; Stanley, K.T.; Stauffer, S.; Jacques, K.M.; Hirsch, D.S.; Takei, J.; Randazzo, P.A. AGAP1, an endosome-associated, phosphoinositide-dependent ADP-ribosylation factor GTPase-activating protein that affects actin cytoskeleton. J. Biol. Chem. 2002, 277, 48965–48975. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.Y.; Rong, R.; Kroll, T.G.; Van Meir, E.G.; Snyder, S.H.; Ye, K. PIKE (phosphatidylinositol 3-kinase enhancer)-A GTPase stimulates Akt activity and mediates cellular invasion. J. Biol. Chem. 2004, 279, 16441–16451. [Google Scholar] [CrossRef] [Green Version]
- Xia, C.; Ma, W.; Stafford, L.J.; Liu, C.; Gong, L.; Martin, J.F.; Liu, M. GGAPs, a new family of bifunctional GTP-binding and GTPase-activating proteins. Mol. Cell Biol. 2003, 23, 2476–2488. [Google Scholar] [CrossRef] [Green Version]
- Nagase, T.; Seki, N.; Ishikawa, K.; Tanaka, A.; Nomura, N. Prediction of the coding sequences of unidentified human genes. V. The coding sequences of 40 new genes (KIAA0161-KIAA0200) deduced by analysis of cDNA clones from human cell line KG-1. DNA Res. 1996, 3, 17–24. [Google Scholar] [CrossRef]
- Rong, R.; Ahn, J.Y.; Huang, H.; Nagata, E.; Kalman, D.; Kapp, J.A.; Tu, J.; Worley, P.F.; Snyder, S.H.; Ye, K. PI3 kinase enhancer-Homer complex couples mGluRI to PI3 kinase, preventing neuronal apoptosis. Nat. Neurosci. 2003, 6, 1153–1161. [Google Scholar] [CrossRef]
- Ye, K.; Hurt, K.J.; Wu, F.Y.; Fang, M.; Luo, H.R.; Hong, J.J.; Blackshaw, S.; Ferris, C.D.; Snyder, S.H. Pike. A nuclear gtpase that enhances PI3kinase activity and is regulated by protein 4.1N. Cell 2000, 103, 919–930. [Google Scholar] [CrossRef]
- Elkahloun, A.G.; Krizman, D.B.; Wang, Z.; Hofmann, T.A.; Roe, B.; Meltzer, P.S. Transcript mapping in a 46-kb sequenced region at the core of 12q13.3 amplification in human cancers. Genomics 1997, 42, 295–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, Z.; Fei, J.; Premont, R.T.; Randazzo, P.A. The Arf GAPs AGAP1 and AGAP2 distinguish between the adaptor protein complexes AP-1 and AP-3. J. Cell Sci. 2005, 118, 3555–3566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Hu, Y.; Hao, C.; Rempel, S.A.; Ye, K. PIKE-A is a proto-oncogene promoting cell growth, transformation and invasion. Oncogene 2007, 26, 4918–4927. [Google Scholar] [CrossRef] [PubMed]
- Rong, R.; Tang, X.; Gutmann, D.H.; Ye, K. Neurofibromatosis 2 (NF2) tumor suppressor merlin inhibits phosphatidylinositol 3-kinase through binding to PIKE-L. Proc. Natl. Acad. Sci. USA 2004, 101, 18200–18205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, J.Y.; Rong, R.; Liu, X.; Ye, K. PIKE/nuclear PI 3-kinase signaling mediates the antiapoptotic actions of NGF in the nucleus. EMBO J. 2004, 23, 3995–4006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Wang, J.; Li, R.; Ayala, G.; Ittmann, M.; Liu, M. GGAP2/PIKE-a directly activates both the Akt and nuclear factor-kappaB pathways and promotes prostate cancer progression. Cancer Res. 2009, 69, 819–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, W.; Feng, Y.I.; Sanders, A.J.; Davies, E.L.; Jiang, W.G. Phosphoinositide-3-Kinase Enhancers, PIKEs: Their Biological Functions and Roles in Cancer. Anticancer Res. 2016, 36, 1103–1109. [Google Scholar]
- Liu, R.; Tian, B.; Gearing, M.; Hunter, S.; Ye, K.; Mao, Z. Cdk5-mediated regulation of the PIKE-A-Akt pathway and glioblastoma cell invasion. Proc. Natl. Acad. Sci. USA 2008, 105, 7570–7575. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Qi, Q.; Chan, C.B.; Zhou, W.; Chen, J.; Luo, H.R.; Appin, C.; Brat, D.J.; Ye, K. Fyn-phosphorylated PIKE-A binds and inhibits AMPK signaling, blocking its tumor suppressive activity. Cell Death Differ. 2016, 23, 52–63. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Sheng, H.; Zhang, X.; Qi, Q.; Chan, C.B.; Li, L.; Shan, C.; Ye, K. Cellular energy stress induces AMPK-mediated regulation of glioblastoma cell proliferation by PIKE-A phosphorylation. Cell Death Dis. 2019, 10, 222. [Google Scholar] [CrossRef] [Green Version]
- Bajno, L.; Peng, X.R.; Schreiber, A.D.; Moore, H.P.; Trimble, W.S.; Grinstein, S. Focal exocytosis of VAMP3-containing vesicles at sites of phagosome formation. J. Cell Biol. 2000, 149, 697–706. [Google Scholar] [CrossRef]
- Allen, L.A.; Yang, C.; Pessin, J.E. Rate and extent of phagocytosis in macrophages lacking vamp3. J. Leukoc. Biol. 2002, 72, 217–221. [Google Scholar] [CrossRef]
- Niedergang, F.; Colucci-Guyon, E.; Dubois, T.; Raposo, G.; Chavrier, P. ADP ribosylation factor 6 is activated and controls membrane delivery during phagocytosis in macrophages. J. Cell Biol. 2003, 161, 1143–1150. [Google Scholar] [CrossRef] [Green Version]
- Murray, R.; Kay, J.G.; Sangermani, D.G.; Stow, J.L. A role for the phagosome in cytokine secretion. Science 2005, 310, 1492–1495. [Google Scholar] [CrossRef] [Green Version]
- Braun, V.; Fraisier, V.; Raposo, G.; Hurbain, I.; Sibarita, J.B.; Chavrier, P.; Galli, T.; Niedergang, F. TI-VAMP/VAMP7 is required for optimal phagocytosis of opsonised particles in macrophages. EMBO J. 2004, 23, 4166–4176. [Google Scholar] [CrossRef] [Green Version]
- Czibener, C.; Sherer, N.M.; Becker, S.M.; Pypaert, M.; Hui, E.; Chapman, E.R.; Mothes, W.; Andrews, N.W. Ca2+ and synaptotagmin VII-dependent delivery of lysosomal membrane to nascent phagosomes. J. Cell Biol. 2006, 174, 997–1007. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Cox, D.; Tseng, C.C.; Donaldson, J.G.; Greenberg, S. A requirement for ARF6 in Fcgamma receptor-mediated phagocytosis in macrophages. J. Biol. Chem. 1998, 273, 19977–19981. [Google Scholar] [CrossRef] [Green Version]
- Uchida, H.; Kondo, A.; Yoshimura, Y.; Mazaki, Y.; Sabe, H. PAG3/Papalpha/KIAA0400, a GTPase-activating protein for ADP-ribosylation factor (ARF), regulates ARF6 in Fcgamma receptor-mediated phagocytosis of macrophages. J. Exp. Med. 2001, 193, 955–966. [Google Scholar] [CrossRef] [Green Version]
- Balañá, M.E.; Niedergang, F.; Subtil, A.; Alcover, A.; Chavrier, P.; Dautry-Varsat, A. ARF6 GTPase controls bacterial invasion by actin remodelling. J. Cell Sci. 2005, 118, 2201–2210. [Google Scholar] [CrossRef] [Green Version]
- Berón, W.; Mayorga, L.S.; Colombo, M.I.; Stahl, P.D. Recruitment of coat-protein-complex proteins on to phagosomal membranes is regulated by a brefeldin A-sensitive ADP-ribosylation factor. Biochem. J. 2001, 355, 409–415. [Google Scholar] [CrossRef]
- Mansfield, P.J.; Hinkovska-Galcheva, V.; Borofsky, M.S.; Shayman, J.A.; Boxer, L.A. Phagocytic signaling molecules in lipid rafts of COS-1 cells transfected with FcgammaRIIA. Biochem. Biophys. Res. Commun. 2005, 331, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Braun, V.; Deschamps, C.; Raposo, G.; Benaroch, P.; Benmerah, A.; Chavrier, P.; Niedergang, F. AP-1 and ARF1 Control Endosomal Dynamics at Sites of FcR mediated Phagocytosis. Mol. Biol. Cell 2007, 18, 4921–4931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naccache, P.H.; Grimard, M.; Roberge, C.J.; Gilbert, C.; Lussier, A.; de Medicis, R.; Poubelle, P.E. Crystal-induced neutrophil activation. I. Initiation and modulation of calcium mobilization and superoxide production by microcrystals. Arthritis Rheum. 1991, 34, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Rollet-Labelle, E.; Marois, S.; Barbeau, K.; Malawista, S.E.; Naccache, P.H. Recruitment of the cross-linked opsonic receptor CD32A (FcgammaRIIA) to high-density detergent-resistant membrane domains in human neutrophils. Biochem. J. 2004, 381, 919–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcil, J.; Harbour, D.; Houle, M.G.; Naccache, P.H.; Bourgoin, S. Monosodium urate-crystal-stimulated phospholipase D in human neutrophils. Biochem. J. 1999, 337 (Pt 2), 185–192. [Google Scholar] [CrossRef] [PubMed]
- Gamara, J.; Chouinard, F.; Davis, L.; Aoudjit, F.; Bourgoin, S.G. Regulators and Effectors of Arf GTPases in Neutrophils. J. Immunol. Res. 2015, 2015, 235170. [Google Scholar] [CrossRef] [Green Version]
- Faugaret, D.; Chouinard, F.C.; Harbour, D.; El azreq, M.A.; Bourgoin, S.G. An essential role for phospholipase D in the recruitment of vesicle amine transport protein-1 to membranes in human neutrophils. Biochem. Pharmacol. 2011, 81, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Bolte, S.; Cordelières, F.P. A guided tour into subcellular colocalization analysis in light microscopy. J. Microsc. 2006, 224, 213–232. [Google Scholar] [CrossRef]
- Ahn, J.Y.; Hu, Y.; Kroll, T.G.; Allard, P.; Ye, K. PIKE-A is amplified in human cancers and prevents apoptosis by up-regulating Akt. Proc. Natl. Acad. Sci. USA 2004, 101, 6993–6998. [Google Scholar] [CrossRef] [Green Version]
- Meurer, S.; Pioch, S.; Wagner, K.; Müller-Esterl, W.; Gross, S. AGAP1, a novel binding partner of nitric oxide-sensitive guanylyl cyclase. J. Biol. Chem. 2004, 279, 49346–49354. [Google Scholar] [CrossRef] [Green Version]
- Chouinard, F.; Centre de Recherche du CHU de Québec—Université Laval, Québec City, QC, Canada. Idendification and characterization of the ArfGAPs expressed by human neutrophils. Unpublished work. 2011. [Google Scholar]
- Randazzo, P.A.; Inoue, H.; Bharti, S. Arf GAPs as regulators of the actin cytoskeleton. Biol. Cell 2007, 99, 583–600. [Google Scholar] [CrossRef] [PubMed]
- Dunphy, J.L.; Ye, K.; Casanova, J.E. Nuclear functions of the Arf guanine nucleotide exchange factor BRAG2. Traffic 2007, 8, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Barbe, L.; Lundberg, E.; Oksvold, P.; Stenius, A.; Lewin, E.; Björling, E.; Asplund, A.; Pontén, F.; Brismar, H.; Uhlén, M.; et al. Toward a confocal subcellular atlas of the human proteome. Mol. Cell Proteomics 2008, 7, 499–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Wu, Y.; Kim, J.; Wang, Z.; Daaka, Y.; Nie, Z. The Arf GTPase-activating protein AGAP2 regulates focal adhesion kinase activity and focal adhesion remodeling. J. Biol. Chem. 2009, 284, 13489–13496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- May, R.C.; Machesky, L.M. Phagocytosis and the actin cytoskeleton. J. Cell Sci. 2001, 114, 1061–1077. [Google Scholar] [CrossRef] [PubMed]
- Indik, Z.K.; Park, J.G.; Hunter, S.; Schreiber, A.D. The molecular dissection of Fc gamma receptor mediated phagocytosis. Blood 1995, 86, 4389–4399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Downey, G.P.; Botelho, R.J.; Butler, J.R.; Moltyaner, Y.; Chien, P.; Schreiber, A.D.; Grinstein, S. Phagosomal maturation, acidification, and inhibition of bacterial growth in nonphagocytic cells transfected with FcgammaRIIA receptors. J. Biol. Chem. 1999, 274, 28436–28444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibarrola, I.; Vossebeld, P.J.; Homburg, C.H.; Thelen, M.; Roos, D.; Verhoeven, A.J. Influence of tyrosine phosphorylation on protein interaction with FcgammaRIIa. Biochim. Biophys. Acta. 1997, 1357, 348–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolotila, M.P.; Diamond, R.D. Stimulation of neutrophil actin polymerization and degranulation by opsonized and unopsonized Candida albicans hyphae and zymosan. Infect. Immun. 1988, 56, 2016–2022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Liu, Z.; Ye, K. Phosphoinositol lipids bind to phosphatidylinositol 3 (PI3)-kinase enhancer GTPase and mediate its stimulatory effect on PI3-kinase and Akt signalings. Proc. Natl. Acad. Sci. USA 2005, 102, 16853–16858. [Google Scholar] [CrossRef] [Green Version]
- Pivot-Pajot, C.; Chouinard, F.; Amine El Azreq, M.; Harbour, D.; Bourgoin, S. Characterisation of degranulation and phagocytic capacity of a human neutrophilic cellular model, PLB-985 cells. Immunobiology 2010, 215, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Corcuera, A.; Ansorena, E.; Montiel-Duarte, C.; Iraburu, M.J. AGAP2: Modulating TGF beta 1-Signaling in the Regulation of Liver Fibrosis. Int. J. Mol. Sci. 2020, 21, 1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, K.; Jang, S.W.; Joshi, J.; Yoo, M.H.; Ye, K. Akt-phosphorylated PIKE-A inhibits UNC5B-induced apoptosis in cancer cell lines in a p53-dependent manner. Mol. Biol. Cell 2011, 22, 1943–1954. [Google Scholar] [CrossRef] [PubMed]
- Liebl, D.; Griffiths, G. Transient assembly of F-actin by phagosomes delays phagosome fusion with lysosomes in cargo-overloaded macrophages. J. Cell Sci. 2009, 122, 2935–2945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, D.; Chang, P.; Zhang, Q.; Reddy, P.G.; Bokoch, G.M.; Greenberg, S. Requirements for both Rac1 and Cdc42 in membrane ruffling and phagocytosis in leukocytes. J. Exp. Med. 1997, 186, 1487–1494. [Google Scholar] [CrossRef]
- Caron, E.; Hall, A. Identification of two distinct mechanisms of phagocytosis controlled by different Rho GTPases. Science 1998, 282, 1717–1721. [Google Scholar] [CrossRef] [Green Version]
- Massol, P.; Montcourrier, P.; Guillemot, J.C.; Chavrier, P. Fc receptor-mediated phagocytosis requires CDC42 and Rac1. EMBO J. 1998, 17, 6219–6229. [Google Scholar] [CrossRef] [Green Version]
- Castellano, F.; Montcourrier, P.; Chavrier, P. Membrane recruitment of Rac1 triggers phagocytosis. J. Cell Sci. 2000, 113 (Pt 17), 2955–2961. [Google Scholar] [CrossRef]
- Chimini, G.; Chavrier, P. Function of Rho family proteins in actin dynamics during phagocytosis and engulfment. Nat. Cell Biol. 2000, 2, E191–E196. [Google Scholar] [CrossRef]
- Hoppe, A.D.; Swanson, J.A. Cdc42, Rac1, and Rac2 display distinct patterns of activation during phagocytosis. Mol. Biol. Cell 2004, 15, 3509–3519. [Google Scholar] [CrossRef] [Green Version]
- Tanguy, E.; Tran Nguyen, A.P.; Kassas, N.; Bader, M.F.; Grant, N.J.; Vitale, N. Regulation of Phospholipase D by Arf6 during FcgammaR-Mediated Phagocytosis. J. Immunol. 2019, 202, 2971–2981. [Google Scholar] [CrossRef] [PubMed]
- Gamara, J.; Davis, L.; Leong, A.Z.; Page, N.; Rollet-Labelle, E.; Zhao, C.; Hongu, T.; Funakoshi, Y.; Kanaho, Y.; Aoudji, F.; et al. Arf6 regulates energy metabolism in neutrophils. Free Radic. Biol. Med. 2021, 172, 550–561. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, J.G.; Honda, A. Localization and function of Arf family GTPases. Biochem. Soc. Trans. 2005, 33, 639–642. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, J.G.; Honda, A.; Weigert, R. Multiple activities for Arf1 at the Golgi complex. Biochim. Biophys. Acta. 2005, 1744, 364–373. [Google Scholar] [CrossRef] [Green Version]
- Mulloy, J.C.; Cancelas, J.A.; Filippi, M.D.; Kalfa, T.A.; Guo, F.; Zheng, Y. Rho GTPases in hematopoiesis and hemopathies. Blood 2010, 115, 936–947. [Google Scholar] [CrossRef]
- Mazaki, Y.; Hashimoto, S.; Tsujimura, T.; Morishige, M.; Hashimoto, A.; Aritake, K.; Yamada, A.; Nam, J.M.; Kiyonari, H.; Nakao, K.; et al. Neutrophil direction sensing and superoxide production linked by the GTPase-activating protein GIT2. Nat. Immunol. 2006, 7, 724–731. [Google Scholar] [CrossRef]
- Gambardella, L.; Anderson, K.E.; Nussbaum, C.; Segonds-Pichon, A.; Margarido, T.; Norton, L.; Ludwig, T.; Sperandio, M.; Hawkins, P.T.; Stephens, L.; et al. The GTPase activating protein ARAP3 regulates chemotaxis and adhesion-dependent processes in neutrophils. Blood 2011, 118, 1087–1098. [Google Scholar] [CrossRef]
- Chung, J.; Serezani, C.H.; Huang, S.K.; Stern, J.N.; Keskin, D.B.; Jagirdar, R.; Brock, T.G.; Aronoff, D.M.; Peters-Golden, M. Rap1 activation is required for Fc gamma receptor-dependent phagocytosis. J. Immunol. 2008, 181, 5501–5509. [Google Scholar]
- Zhang, J.; Guo, J.; Dzhagalov, I.; He, Y.W. An essential function for the calcium-promoted Ras inactivator in Fcgamma receptor-mediated phagocytosis. Nat. Immunol. 2005, 6, 911–919. [Google Scholar] [CrossRef] [Green Version]
- Cuthbert, E.J.; Davis, K.K.; Casanova, J.E. Substrate specificities and activities of AZAP family Arf GAPs in vivo. Am. J. Physiol. Cell Physiol. 2008, 294, C263–C270. [Google Scholar] [CrossRef] [Green Version]
- Beemiller, P.; Hoppe, A.D.; Swanson, J.A. A phosphatidylinositol-3-kinase-dependent signal transition regulates ARF1 and ARF6 during Fcgamma receptor-mediated phagocytosis. PLoS Biol. 2006, 4, e162. [Google Scholar] [CrossRef]
- Thacker, E.; Kearns, B.; Chapman, C.; Hammond, J.; Howell, A.; Theibert, A. The arf6 GAP centaurin alpha-1 is a neuronal actin-binding protein which also functions via GAP-independent activity to regulate the actin cytoskeleton. Eur. J. Cell Biol. 2004, 83, 541–554. [Google Scholar] [CrossRef]
- Venkateswarlu, K.; Hanada, T.; Chishti, A.H. Centaurin-alpha1 interacts directly with kinesin motor protein KIF13B. J. Cell Sci. 2005, 118, 2471–2484. [Google Scholar] [CrossRef] [Green Version]
- Horiguchi, K.; Hanada, T.; Fukui, Y.; Chishti, A.H. Transport of PIP3 by GAKIN, a kinesin-3 family protein, regulates neuronal cell polarity. J. Cell Biol. 2006, 174, 425–436. [Google Scholar] [CrossRef] [Green Version]
- Tong, Y.; Tempel, W.; Wang, H.; Yamada, K.; Shen, L.; Senisterra, G.A.; MacKenzie, F.; Chishti, A.H.; Park, H.W. Phosphorylation-independent dual-site binding of the FHA domain of KIF13 mediates phosphoinositide transport via centaurin alpha1. Proc. Natl. Acad. Sci. USA 2010, 107, 20346–20351. [Google Scholar] [CrossRef] [Green Version]
- Khandani, A.; Eng, E.; Jongstra-Bilen, J.; Schreiber, A.D.; Douda, D.; Samavarchi-Tehrani, P.; Harrison, R.E. Microtubules regulate PI-3K activity and recruitment to the phagocytic cup during Fcgamma receptor-mediated phagocytosis in nonelicited macrophages. J. Leukoc. Biol. 2007, 82, 417–428. [Google Scholar] [CrossRef] [Green Version]
- Silver, K.E.; Harrison, R.E. Kinesin 5B is necessary for delivery of membrane and receptors during FcγR-mediated phagocytosis. J. Immunol. 2011, 186, 816–825. [Google Scholar] [CrossRef] [Green Version]
- Soundararajan, M.; Yang, X.; Elkins, J.M.; Sobott, F.; Doyle, D.A. The centaurin gamma-1 GTPase-like domain functions as an NTPase. Biochem. J. 2007, 401, 679–688. [Google Scholar] [CrossRef] [Green Version]
- Cheng, N.; Zhang, H.; Zhang, S.Y.; Ma, X.D.; Meng, G.Y. Crystal structure of the GTP-binding protein-like domain of AGAP1. Acta Crystallogr. F 2021, 77, 105–112. [Google Scholar] [CrossRef]
- Luo, R.B.; Akpan, I.O.; Hayashi, R.; Sramko, M.; Barr, V.; Shiba, Y.; Randazzo, P.A. GTP-binding Protein-like Domain of AGAP1 Is Protein Binding Site That Allosterically Regulates ArfGAP Protein Catalytic Activity. J. Biol. Chem. 2012, 287, 17176–17185. [Google Scholar] [CrossRef] [Green Version]
- Braun, V.; Niedergang, F. Linking exocytosis and endocytosis during phagocytosis. Biol. Cell 2006, 98, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Huynh, K.K.; Kay, J.G.; Stow, J.L.; Grinstein, S. Fusion, fission, and secretion during phagocytosis. Physiology 2007, 22, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Vashi, N.; Andrabi, S.B.A.; Ghanwat, S.; Suar, M.; Kumar, D. Ca2+-dependent Focal Exocytosis of Golgi-derived Vesicles Helps Phagocytic Uptake in Macrophages. J. Biol. Chem. 2017, 292, 5144–5165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiba, Y.; Römer, W.; Mardones, G.A.; Burgos, P.V.; Lamaze, C.; Johannes, L. AGAP2 regulates retrograde transport between early endosomes and the TGN. J. Cell Sci. 2010, 123, 2381–2390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botelho, R.J.; Teruel, M.; Dierckman, R.; Anderson, R.; Wells, A.; York, J.D.; Meyer, T.; Grinstein, S. Localized biphasic changes in phosphatidylinositol-4,5-bisphosphate at sites of phagocytosis. J. Cell Biol. 2000, 151, 1353–1368. [Google Scholar] [CrossRef]
- Marshall, J.G.; Booth, J.W.; Stambolic, V.; Mak, T.; Balla, T.; Schreiber, A.D.; Meyer, T.; Grinstein, S. Restricted accumulation of phosphatidylinositol 3-kinase products in a plasmalemmal subdomain during Fc gamma receptor-mediated phagocytosis. J. Cell Biol. 2001, 153, 1369–1380. [Google Scholar] [CrossRef] [Green Version]
- Ostrowski, P.P.; Freeman, S.A.; Fairn, G.; Grinstein, S. Dynamic Podosome-Like Structures in Nascent Phagosomes Are Coordinated by Phosphoinositides. Dev. Cell 2019, 50, 397–410.e393. [Google Scholar] [CrossRef]
- Marois, L.; Pare, G.; Vaillancourt, M.; Rollet-Labelle, E.; Naccache, P.H. Fc gammaRIIIb triggers raft-dependent calcium influx in IgG-mediated responses in human neutrophils. J. Biol. Chem. 2011, 286, 3509–3519. [Google Scholar] [CrossRef] [Green Version]
- Marois, L.; Vaillancourt, M.; Marois, S.; Proulx, S.; Pare, G.; Rollet-Labelle, E.; Naccache, P.H. The ubiquitin ligase c-Cbl down-regulates FcgammaRIIa activation in human neutrophils. J. Immunol. 2009, 182, 2374–2384. [Google Scholar] [CrossRef] [Green Version]
- Popa-Nita, O.; Rollet-Labelle, E.; Thibault, N.; Gilbert, C.; Bourgoin, S.G.; Naccache, P.H. Crystal-induced neutrophil activation. IX. Syk-dependent activation of class Ia phosphatidylinositol 3-kinase. J. Leukoc. Biol. 2007, 82, 763–773. [Google Scholar] [CrossRef]
- Nishi, H.; Furuhashi, K.; Cullere, X.; Saggu, G.; Miller, M.J.; Chen, Y.; Rosetti, F.; Hamilton, S.L.; Yang, L.; Pittman, S.P.; et al. Neutrophil FcgammaRIIA promotes IgG-mediated glomerular neutrophil capture via Abl/Src kinases. J. Clin. Investig. 2017, 127, 3810–3826. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, S.; Sitaru, C.; Jakus, Z.; Anderson, K.E.; Damoulakis, G.; Davidson, K.; Hirose, M.; Juss, J.; Oxley, D.; Chessa, T.A.; et al. PI3Kbeta plays a critical role in neutrophil activation by immune complexes. Sci. Signal. 2011, 4, ra23. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chouinard, F.C.; Davis, L.; Gilbert, C.; Bourgoin, S.G. Functional Role of AGAP2/PIKE-A in Fcγ Receptor-Mediated Phagocytosis. Cells 2023, 12, 72. https://doi.org/10.3390/cells12010072
Chouinard FC, Davis L, Gilbert C, Bourgoin SG. Functional Role of AGAP2/PIKE-A in Fcγ Receptor-Mediated Phagocytosis. Cells. 2023; 12(1):72. https://doi.org/10.3390/cells12010072
Chicago/Turabian StyleChouinard, François C., Lynn Davis, Caroline Gilbert, and Sylvain G. Bourgoin. 2023. "Functional Role of AGAP2/PIKE-A in Fcγ Receptor-Mediated Phagocytosis" Cells 12, no. 1: 72. https://doi.org/10.3390/cells12010072