


Exploring the Nerve Regenerative Capacity of Compounds with Differing Affinity for PPARγ In Vitro and In Vivo
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures
2.1.1. SCL4.1/F7 Schwann Cell Line
2.1.2. PC12 Neuronal Cell Line
2.2. Fabrication of 3D EngNT Co-Cultures
2.3. Immunocytochemistry
2.4. Surgical Nerve Injury Models In Vivo
2.5. Nerve Tissue Harvest
2.6. Nerve Tissue Analysis
2.6.1. Cryo-Sectioning
2.6.2. Immunohistochemistry
2.7. Image Analysis and Quantification
2.8. Functional Outcomes In Vivo
2.8.1. Electrophysiology
2.8.2. Static Sciatic Index (SSI)
2.8.3. Von Frey
2.9. Statistical Analysis
3. Results
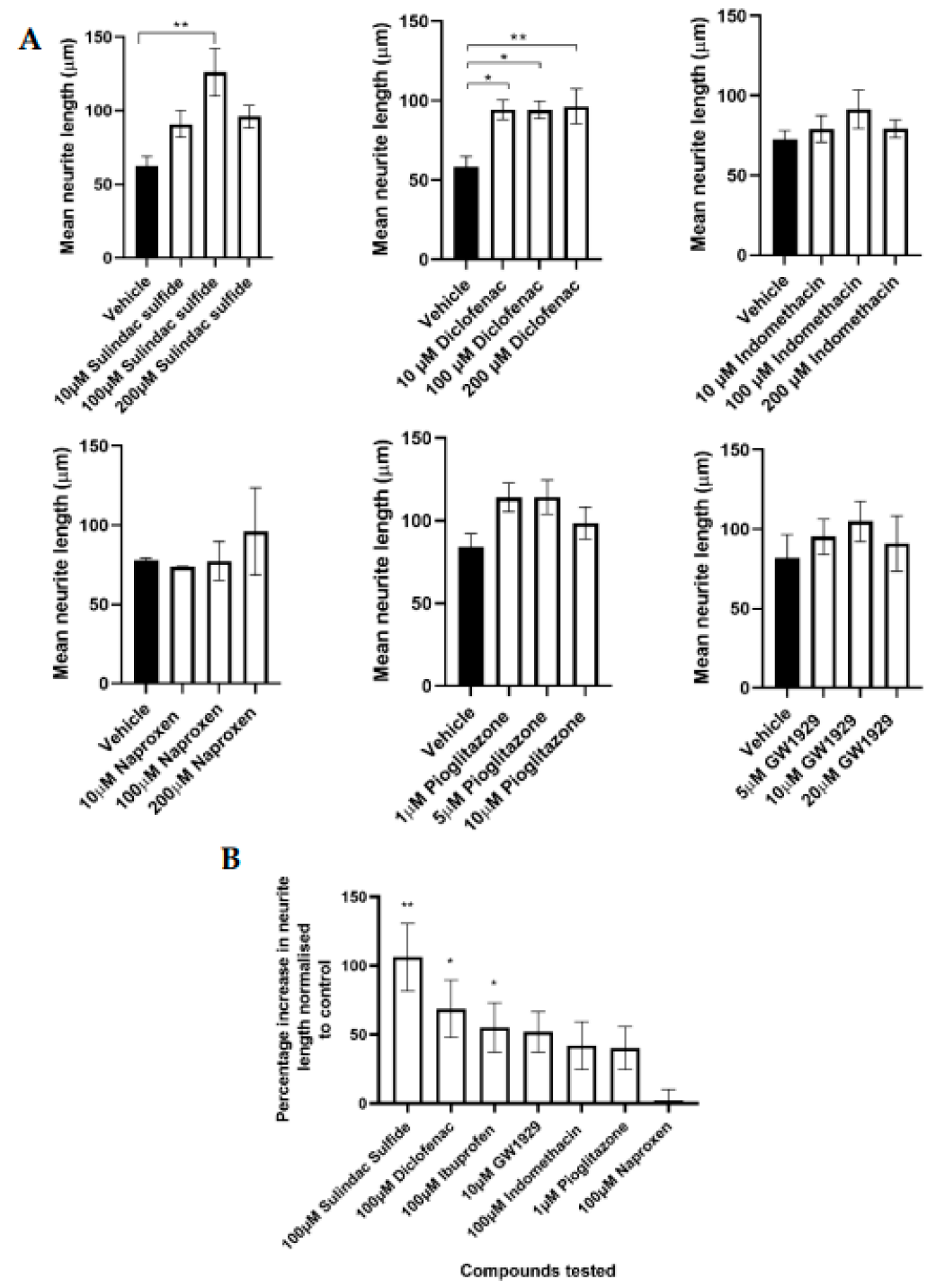
3.1. Regenerative Capacity of PPARγ Agonists In Vitro
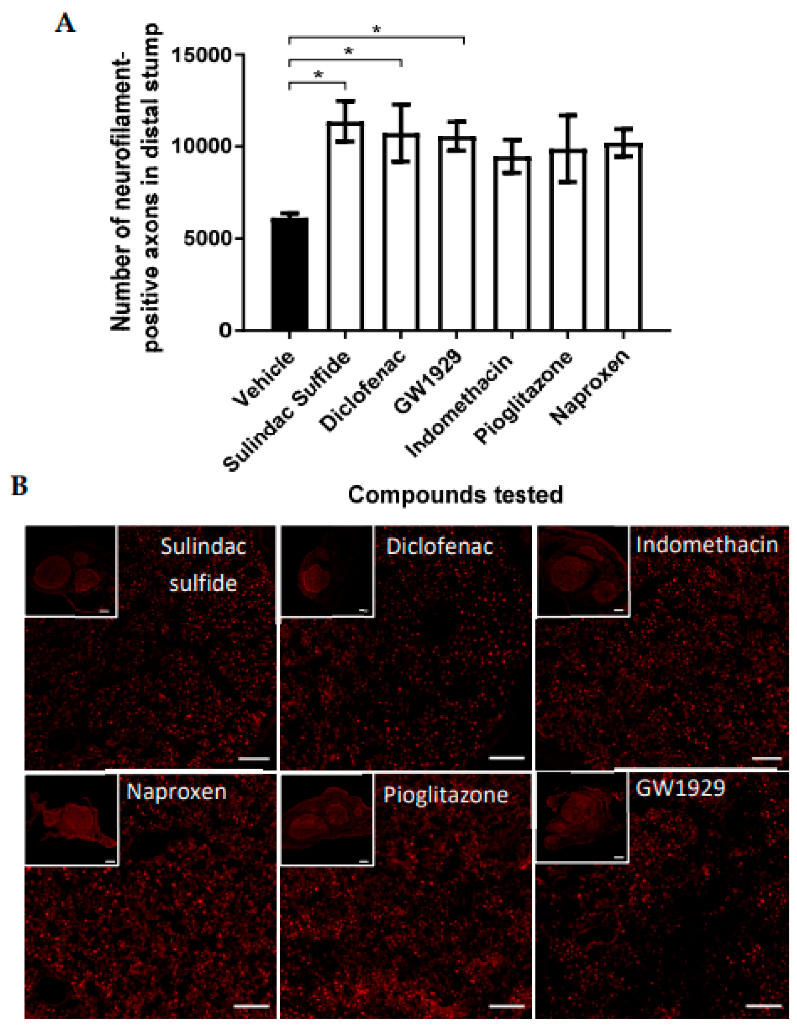
3.2. Effect on Neuronal Growth of PPARγ Agonists In Vivo
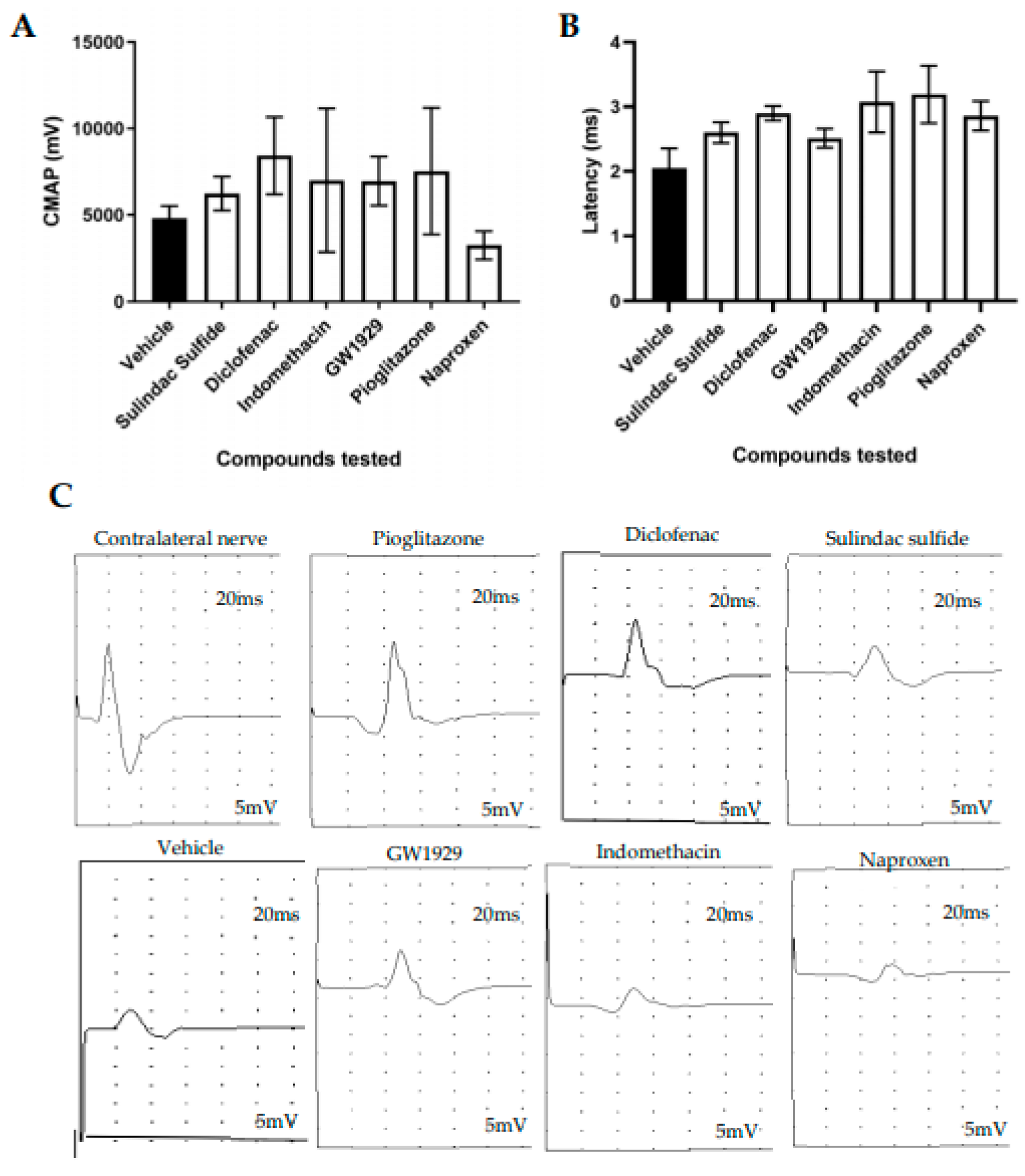
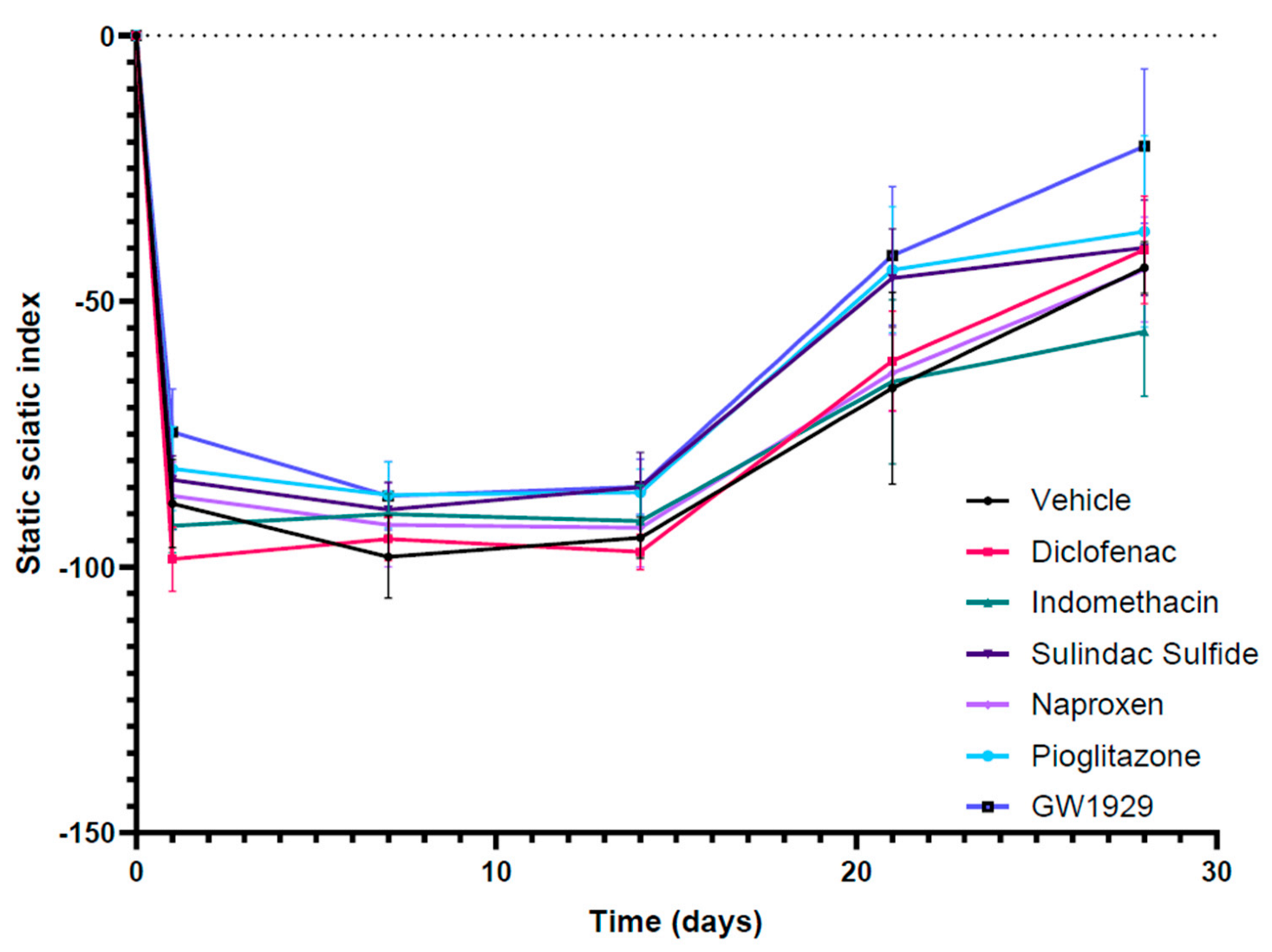
3.3. Functional Recovery Following In Vivo Treatment with PPARγ Agonists
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hoke, A.; Brushart, T. Introduction to special issue: Challenges and opportunities for regeneration in the peripheral nervous system. Exp. Neurol. 2010, 223, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wada, K.; Nakajima, A.; Katayama, K.; Kudo, C.; Shibuya, A.; Kubota, N.; Terauchi, Y.; Tachibana, M.; Miyoshi, H.; Kamisaki, Y.; et al. Peroxisome proliferator-activated receptor gamma-mediated regulation of neural stem cell proliferation and differentiation. J. Biol. Chem. 2006, 281, 12673–12681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamagishi, S.; Ogasawara, S.; Mizukami, H.; Yajima, N.; Wada, R.; Sugawara, A.; Yagihashi, S. Correction of protein kinase C activity and macrophage migration in peripheral nerve by pioglitazone, peroxisome proliferator activated-gamma-ligand, in insulin-deficient diabetic rats. J. Neurochem. 2008, 104, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Wang, Q.; Zhou, Z.; Wang, Y.; Liu, Y.; Ji, Y.; Liu, F. Changes of peroxisome proliferator-activated receptor-gamma on crushed rat sciatic nerves and differentiated primary Schwann cells. J. Mol. Neurosci. 2012, 47, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Liu, F.; Yan, M.; Ji, H.; Hu, L.; Li, X.; Qian, J.; He, X.; Zhang, L.; Shen, A.; et al. Peroxisome proliferator-activated receptor-gamma agonists suppress iNOS expression induced by LPS in rat primary Schwann cells. J. Neuroimmunol. 2010, 218, 36–47. [Google Scholar] [CrossRef]
- Lezana, J.P.; Dagan, S.Y.; Robinson, A.; Goldstein, R.S.; Fainzilber, M.; Bronfman, F.C.; Bronfman, M. Axonal PPARgamma promotes neuronal regeneration after injury. Dev. Neurobiol. 2016, 76, 688–701. [Google Scholar] [CrossRef]
- Wakino, S.; Hayashi, K.; Kanda, T.; Tatematsu, S.; Homma, K.; Yoshioka, K.; Takamatsu, I.; Saruta, T. Peroxisome proliferator-activated receptor gamma ligands inhibit Rho/Rho kinase pathway by inducing protein tyrosine phosphatase SHP-2. Circ. Res. 2004, 95, e45–e55. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.M.; Gordon, T.; Zochodne, D.W.; Power, H.A. Improving peripheral nerve regeneration: From molecular mechanisms to potential therapeutic targets. Exp. Neurol. 2014, 261, 826–835. [Google Scholar] [CrossRef]
- Madura, T.; Tomita, K.; Terenghi, G. Ibuprofen improves functional outcome after axotomy and immediate repair in the peripheral nervous system. J. Plast. Reconstr. Aesthet. Surg. 2011, 64, 1641–1646. [Google Scholar] [CrossRef]
- Hiraga, A.; Kuwabara, S.; Doya, H.; Kanai, K.; Fujitani, M.; Taniguchi, J.; Arai, K.; Mori, M.; Hattori, T.; Yamashita, T. Rho-kinase inhibition enhances axonal regeneration after peripheral nerve injury. J. Peripher. Nerv. Syst. 2006, 11, 217–224. [Google Scholar] [CrossRef]
- Joshi, A.R.; Bobylev, I.; Zhang, G.; Sheikh, K.A.; Lehmann, H.C. Inhibition of Rho-kinase differentially affects axon regeneration of peripheral motor and sensory nerves. Exp. Neurol. 2015, 263, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.M.; Moore, L.B.; Smith-Oliver, T.A.; Wilkison, W.O.; Willson, T.M.; Kliewer, S.A. An antidiabetic thiazolidinedione is a high affinity ligand for peroxisome proliferator-activated receptor gamma (PPAR gamma). J. Biol. Chem. 1995, 270, 12953–12956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintanilla, R.A.; Godoy, J.A.; Alfaro, I.; Cabezas, D.; von Bernhardi, R.; Bronfman, M.; Inestrosa, N.C. Thiazolidinediones promote axonal growth through the activation of the JNK pathway. PLoS ONE 2013, 8, e65140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammadi, R.; Hirsaee, M.A.; Amini, K. Improvement of functional recovery of transected peripheral nerve by means of artery grafts filled with diclofenac. Int. J. Surg. 2013, 11, 259–264. [Google Scholar] [CrossRef] [Green Version]
- Rayner, M.L.D.; Grillo, A.; Williams, G.R.; Tawfik, E.; Zhang, T.; Volitaki, C.; Craig, D.Q.M.; Healy, J.; Phillips, J.B. Controlled local release of PPARgamma agonists from biomaterials to treat peripheral nerve injury. J. Neural Eng. 2020, 17, 046030. [Google Scholar] [CrossRef] [PubMed]
- Tamaddonfard, E.; Farshid, A.A.; Samadi, F.; Eghdami, K. Effect of vitamin B12 on functional recovery and histopathologic changes of tibial nerve-crushed rats. Drug Res. 2014, 64, 470–475. [Google Scholar] [CrossRef]
- Kopp, M.A.; Liebscher, T.; Niedeggen, A.; Laufer, S.; Brommer, B.; Jungehulsing, G.J.; Strittmatter, S.M.; Dirnagl, U.; Schwab, J.M. Small-molecule-induced Rho-inhibition: NSAIDs after spinal cord injury. Cell Tissue Res. 2012, 349, 119–132. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Budel, S.; Baughman, K.; Gould, G.; Song, K.H.; Strittmatter, S.M. Ibuprofen enhances recovery from spinal cord injury by limiting tissue loss and stimulating axonal growth. J. Neurotrauma 2009, 26, 81–95. [Google Scholar] [CrossRef]
- Lehmann, J.M.; Lenhard, J.M.; Oliver, B.B.; Ringold, G.M.; Kliewer, S.A. Peroxisome proliferator-activated receptors alpha and gamma are activated by indomethacin and other non-steroidal anti-inflammatory drugs. J. Biol. Chem. 1997, 272, 3406–3410. [Google Scholar] [CrossRef] [Green Version]
- Dill, J.; Patel, A.R.; Yang, X.L.; Bachoo, R.; Powell, C.M.; Li, S. A molecular mechanism for ibuprofen-mediated RhoA inhibition in neurons. J. Neurosci. 2010, 30, 963–972. [Google Scholar] [CrossRef] [Green Version]
- Fu, Q.; Hue, J.; Li, S. Nonsteroidal anti-inflammatory drugs promote axon regeneration via RhoA inhibition. J. Neurosci. 2007, 27, 4154–4164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Su, Y.; Li, B.; Liu, F.; Ryder, J.W.; Wu, X.; Gonzalez-DeWhitt, P.A.; Gelfanova, V.; Hale, J.E.; May, P.C.; et al. Nonsteroidal anti-inflammatory drugs can lower amyloidogenic Abeta42 by inhibiting Rho. Science 2003, 302, 1215–1217. [Google Scholar] [CrossRef] [PubMed]
- Puhl, A.C.; Milton, F.A.; Cvoro, A.; Sieglaff, D.H.; Campos, J.C.; Bernardes, A.; Filgueira, C.S.; Lindemann, J.L.; Deng, T.; Neves, F.A.; et al. Mechanisms of peroxisome proliferator activated receptor gamma regulation by non-steroidal anti-inflammatory drugs. Nucl. Recept. Signal. 2015, 13, e004. [Google Scholar] [CrossRef] [Green Version]
- Rayner, M.L.D.; Healy, J.; Phillips, J.B. Repurposing Small Molecules to Target PPAR-gamma as New Therapies for Peripheral Nerve Injuries. Biomolecules 2021, 11, 1301. [Google Scholar] [CrossRef] [PubMed]
- Xing, B.; Li, H.; Wang, H.; Mukhopadhyay, D.; Fisher, D.; Gilpin, C.J.; Li, S. RhoA-inhibiting NSAIDs promote axonal myelination after spinal cord injury. Exp. Neurol. 2011, 231, 247–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, J.B.; Brown, R. Micro-structured materials and mechanical cues in 3D collagen gels. Methods Mol. Biol. 2011, 695, 183–196. [Google Scholar] [CrossRef]
- East, E.; de Oliveira, D.B.; Golding, J.P.; Phillips, J.B. Alignment of astrocytes increases neuronal growth in three-dimensional collagen gels and is maintained following plastic compression to form a spinal cord repair conduit. Tissue Eng. Part A 2010, 16, 3173–3184. [Google Scholar] [CrossRef]
- Rayner, M.L.D.; Laranjeira, S.; Evans, R.E.; Shipley, R.J.; Healy, J.; Phillips, J.B. Developing an In Vitro Model to Screen Drugs for Nerve Regeneration. Anat. Rec. 2018, 301, 1628–1637. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Phillips, J.B. Monitoring neuron and astrocyte interactions with a 3D cell culture system. Methods Mol. Biol. 2014, 1162, 113–124. [Google Scholar] [CrossRef]
- Bervar, M. Video analysis of standing--an alternative footprint analysis to assess functional loss following injury to the rat sciatic nerve. J. Neurosci. Methods 2000, 102, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Lamotte, Y.; Martres, P.; Faucher, N.; Laroze, A.; Grillot, D.; Ancellin, N.; Saintillan, Y.; Beneton, V.; Gampe, R.T., Jr. Synthesis and biological activities of novel indole derivatives as potent and selective PPARgamma modulators. Bioorg. Med. Chem. Lett. 2010, 20, 1399–1404. [Google Scholar] [CrossRef] [PubMed]
- Obermoser, V.; Mauersberger, R.; Schuster, D.; Czifersky, M.; Lipova, M.; Siegl, M.; Kintscher, U.; Gust, R. Importance of 5/6-aryl substitution on the pharmacological profile of 4′-((2-propyl-1H-benzo[d]imidazol-1-yl)methyl)-[1,1′-biphenyl]-2-carboxylic acid derived PPARgamma agonists. Eur. J. Med. Chem. 2017, 126, 590–603. [Google Scholar] [CrossRef] [PubMed]
- Quintanilla, R.A.; Jin, Y.N.; Fuenzalida, K.; Bronfman, M.; Johnson, G.V.W. Rosiglitazone treatment prevents mitochondrial dysfunction in mutant huntingtin-expressing cells: Possible role of peroxisome proliferator-activated receptor-gamma (PPARgamma) in the pathogenesis of Huntington disease. J. Biol. Chem. 2008, 283, 25628–25637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeGeer, J.; Lamarche-Vane, N. Rho GTPases in neurodegeneration diseases. Exp. Cell Res. 2013, 319, 2384–2394. [Google Scholar] [CrossRef]
- Lee, J.K.; Paine, M.F.; Brouwer, K.L. Sulindac and its metabolites inhibit multiple transport proteins in rat and human hepatocytes. J. Pharmacol. Exp. Ther. 2010, 334, 410–418. [Google Scholar] [CrossRef] [Green Version]
- Gurpinar, E.; Grizzle, W.E.; Piazza, G.A. COX-Independent Mechanisms of Cancer Chemoprevention by Anti-Inflammatory Drugs. Front. Oncol. 2013, 3, 181. [Google Scholar] [CrossRef] [Green Version]
- Ayranci, E.; Altunkaynak, B.Z.; Aktas, A.; Ragbetli, M.C.; Kaplan, S. Prenatal exposure of diclofenac sodium affects morphology but not axon number of the median nerve of rats. Folia Neuropathol. 2013, 51, 76–86. [Google Scholar] [CrossRef]
- Canan, S.; Aktas, A.; Ulkay, M.B.; Colakoglu, S.; Ragbetli, M.C.; Ayyildiz, M.; Geuna, S.; Kaplan, S. Prenatal exposure to a non-steroidal anti-inflammatory drug or saline solution impairs sciatic nerve morphology: A stereological and histological study. Int. J. Dev. Neurosci. 2008, 26, 733–738. [Google Scholar] [CrossRef]
- Colakoglu, S.; Aktas, A.; Raimondo, S.; Turkmen, A.P.; Altunkaynak, B.Z.; Odaci, E.; Geuna, S.; Kaplan, S. Effects of prenatal exposure to diclofenac sodium and saline on the optic nerve of 4- and 20-week-old male rats: A stereological and histological study. Biotech. Histochem. 2014, 89, 136–144. [Google Scholar] [CrossRef]
- Keskin, I.; Kaplan, S.; Kalkan, S.; Sutcu, M.; Ulkay, M.B.; Esener, O.B. Evaluation of neuroprotection by melatonin against adverse effects of prenatal exposure to a nonsteroidal anti-inflammatory drug during peripheral nerve development. Int. J. Dev. Neurosci. 2015, 41, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, A.A.; Yurt, K.K.; Deniz, O.G.; Altun, G. Peripheral nerve and diclofenac sodium: Molecular and clinical approaches. J. Chem. Neuroanat. 2018, 87, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Smith, U. Pioglitazone: Mechanism of action. Int. J. Clin. Pract. Suppl. 2001, 121, 13–18. [Google Scholar]
- Zhao, Y.; Patzer, A.; Herdegen, T.; Gohlke, P.; Culman, J. Activation of cerebral peroxisome proliferator-activated receptors gamma promotes neuroprotection by attenuation of neuronal cyclooxygenase-2 overexpression after focal cerebral ischemia in rats. FASEB J. 2006, 20, 1162–1175. [Google Scholar] [CrossRef] [Green Version]
- Eto, M.; Sumi, H.; Fujimura, H.; Yoshikawa, H.; Sakoda, S. Pioglitazone promotes peripheral nerve remyelination after crush injury through CD36 upregulation. J. Peripher. Nerv. Syst. 2008, 13, 242–248. [Google Scholar] [CrossRef]
- Camara-Lemarroy, C.R.; Guzman-de la Garza, F.J.; Barrera-Oranday, E.A.; Cabello-Garcia, A.J.; Garcia-Tamez, A.; Fernandez-Garza, N.E. Celecoxib accelerates functional recovery after sciatic nerve crush in the rat. J. Brachial Plex. Peripher. Nerve Inj. 2008, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Subbanna, P.K.; Prasanna, C.G.; Gunale, B.K.; Tyagi, M.G. Acetyl salicylic acid augments functional recovery following sciatic nerve crush in mice. J. Brachial Plex. Peripher. Nerve Inj. 2007, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, R.; Mehrtash, M.; Nikonam, N.; Mehrtash, M.; Amini, K. Ketoprofen combined with artery graft entubulization improves functional recovery of transected peripheral nerves. J. Cranio-Maxillo-Facial Surg. 2014, 42, 2076–2081. [Google Scholar] [CrossRef]
- Tontonoz, P.; Spiegelman, B.M. Fat and beyond: The diverse biology of PPARgamma. Annu. Rev. Biochem. 2008, 77, 289–312. [Google Scholar] [CrossRef]
- Bouhlel, M.A.; Derudas, B.; Rigamonti, E.; Dievart, R.; Brozek, J.; Haulon, S.; Zawadzki, C.; Jude, B.; Torpier, G.; Marx, N.; et al. PPARgamma activation primes human monocytes into alternative M2 macrophages with anti-inflammatory properties. Cell Metab. 2007, 6, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Navarro, X. Functional evaluation of peripheral nerve regeneration and target reinnervation in animal models: A critical overview. Eur. J. Neurosci. 2016, 43, 271–286. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Agata, V.; Trovato, B.; Roggio, F.; Castorina, A.; Vecchio, M.; Di Rosa, M.; Musumeci, G. The role of exercise on peripheral nerve regeneration: From animal model to clinical application. Heliyon 2021, 7, e08281. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
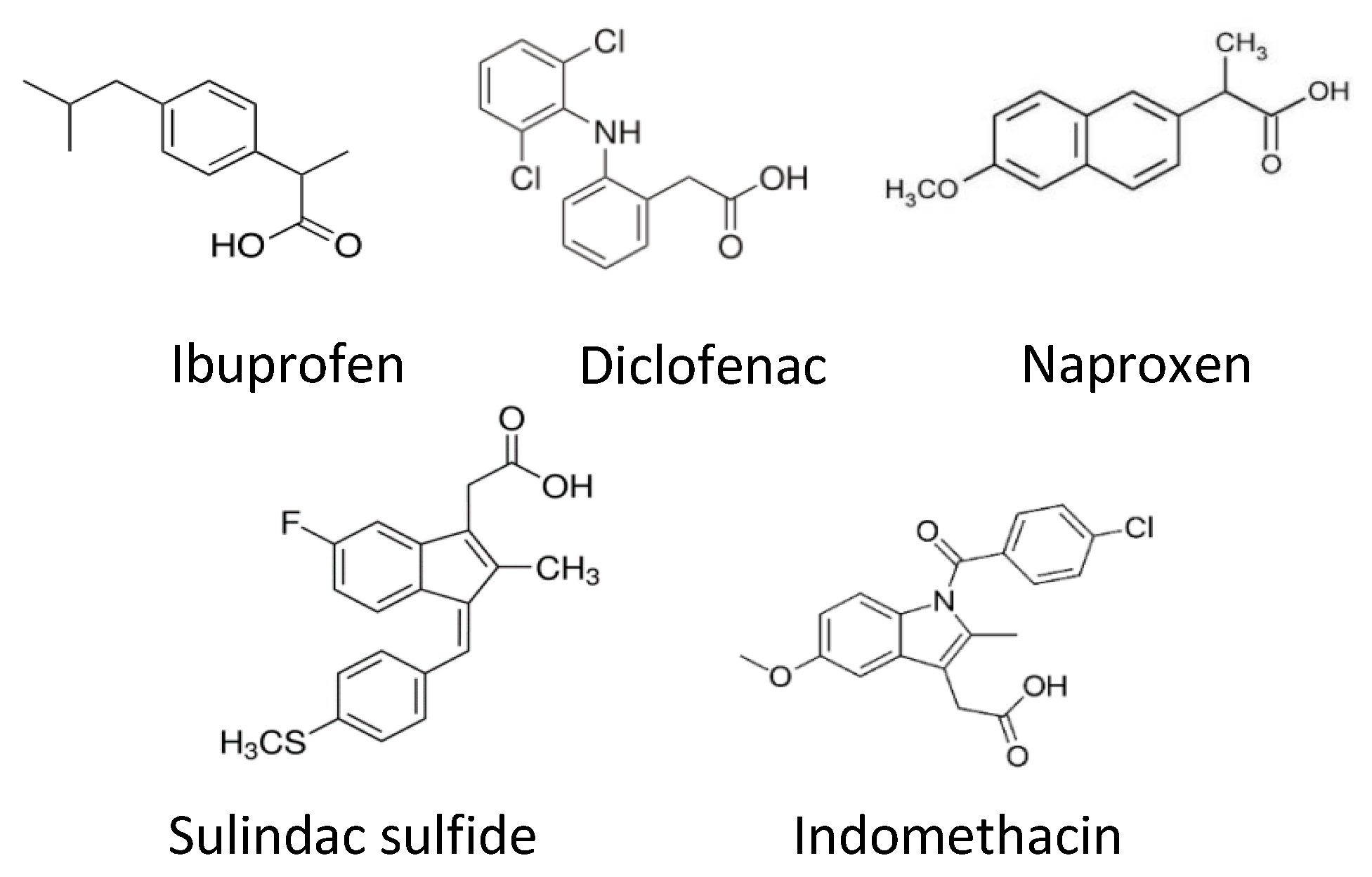{kind=link}
| Compound | PPARγ EC50 | Dose of Compound Tested In Vitro (µM) | Neurite Length Seen with Optimal Dose Percentage Increase to the Control (Mean ± SEM) |
|---|---|---|---|
| GW1929 | 13 nM [32] | 10 | 51% ± 14.74 |
| Pioglitazone | 380 nM [33] | 1 | 40% ± 15.58 |
| Sulindac sulfide | 1.87 µM [23] | 100 | 106% ± 24.61 |
| Diclofenac | 1 µM [23] | 100 | 73.93% ± 28.65 |
| Ibuprofen | 56.8 µM [23] | 100 | 55.2% ± 17.92 [28] |
| Indomethacin | 21 µM [23] | 100 | 19.3% ± 17.35 |
| Naproxen | No action [20] | 100 | 1.8% ± 8.44 |
| GW9662 | 2 nM [34] Irreversible Antagonist | 100 | −89% ± 10.41 [28] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rayner, M.L.D.; Kellaway, S.C.; Kingston, I.; Guillemot-Legris, O.; Gregory, H.; Healy, J.; Phillips, J.B. Exploring the Nerve Regenerative Capacity of Compounds with Differing Affinity for PPARγ In Vitro and In Vivo. Cells 2023, 12, 42. https://doi.org/10.3390/cells12010042
Rayner MLD, Kellaway SC, Kingston I, Guillemot-Legris O, Gregory H, Healy J, Phillips JB. Exploring the Nerve Regenerative Capacity of Compounds with Differing Affinity for PPARγ In Vitro and In Vivo. Cells. 2023; 12(1):42. https://doi.org/10.3390/cells12010042
Chicago/Turabian StyleRayner, Melissa L. D., Simon C. Kellaway, Isabel Kingston, Owein Guillemot-Legris, Holly Gregory, Jess Healy, and James B. Phillips. 2023. "Exploring the Nerve Regenerative Capacity of Compounds with Differing Affinity for PPARγ In Vitro and In Vivo" Cells 12, no. 1: 42. https://doi.org/10.3390/cells12010042