
Rank-Rankl-Opg Axis in Multiple Sclerosis: The Contribution of Placenta

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Placental Tissues from Healthy Pregnant Women
2.2. Serum Collection and Processing
2.3. Human Placenta Explant Culture and Treatment
2.4. Astrocytes Cell Culture and Treatment
2.5. Immunoprecipitation
2.6. RNA Isolation and qRT-PCR
2.7. Western Blot Analysis
2.8. Immunohistochemistry
2.9. Immunofluorescence
2.10. ELISA
2.11. Statistical Analyses
3. Results
3.1. OPG and sRANKL in Serum of MS-Affected and Healthy Pregnant and Non-Pregnant Women
3.1.1. Pregnancy Increases OPG Serum Levels and Differentially Regulates Srankl in MS-Affected and Healthy Women
3.1.2. Pregnancy Influences the Balance between Srankl and Opg in Ms-Affected Women, Returning It to the Conditions of Healthy Subjects
3.1.3. OPG in Maternal and Cord Blood Serum
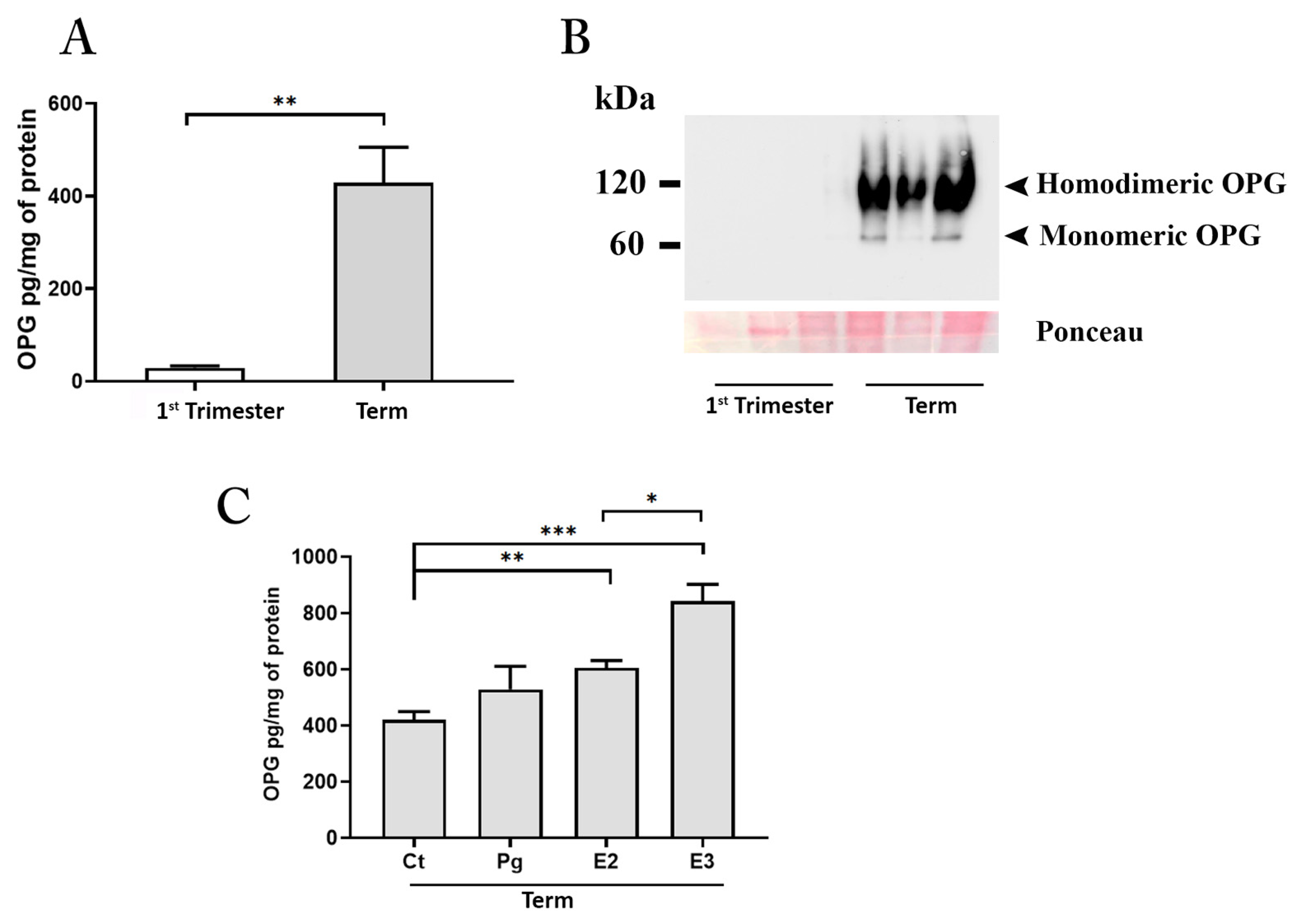
3.2. Human Placenta Is an Invaluable Source of OPG
3.2.1. OPG Production and Distribution in Placental Tissues
3.2.2. OPG Is Highly Released by Placenta Explant Cultures Established from Tissues at Term of Pregnancy
3.3. Role of RANK-RANKL-OPG System in Astrocytes
3.3.1. Astrocytes Activation by RANKL
3.3.2. The Role of Placental OPG in Astrocyte Activation
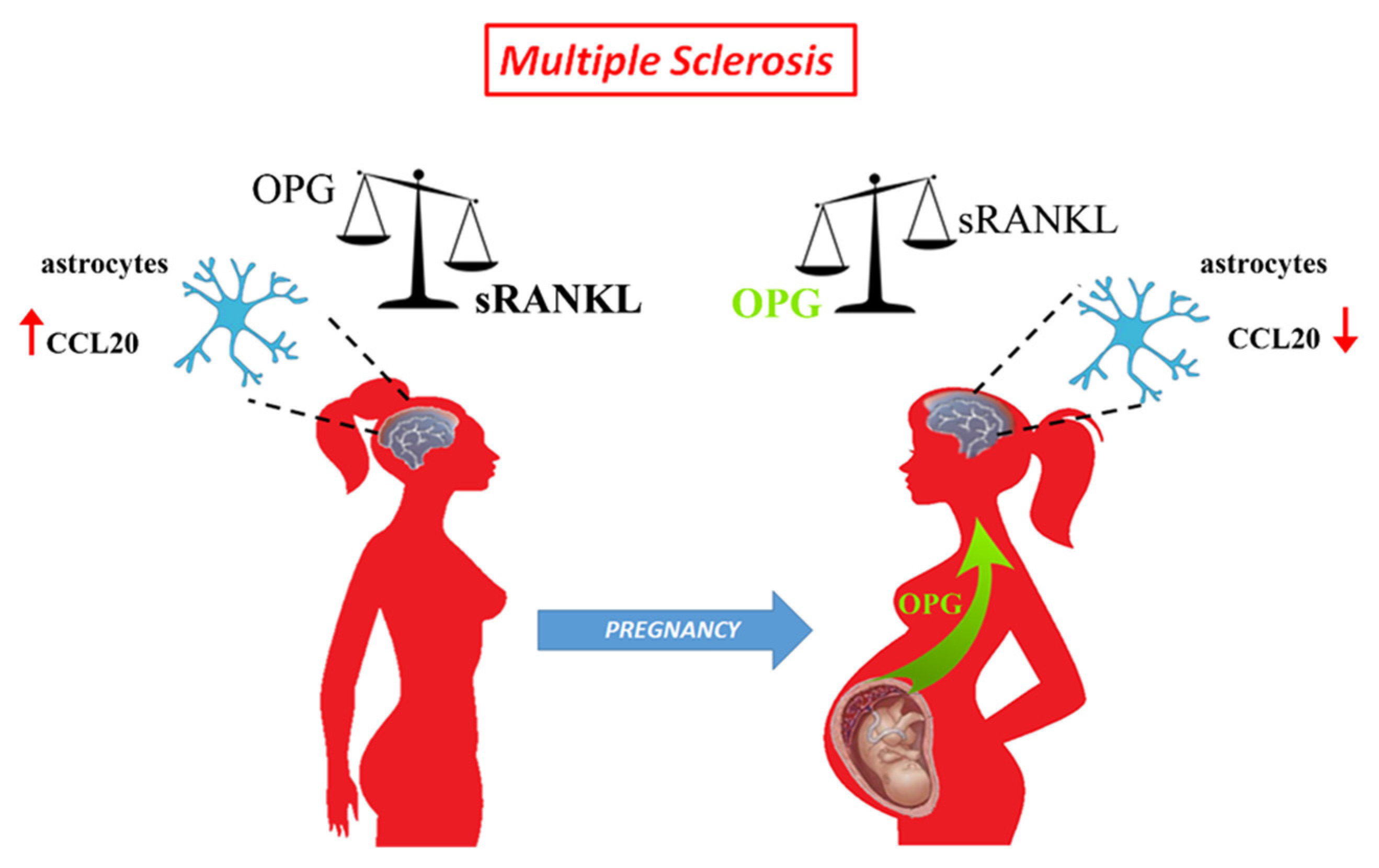
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Confavreux, C.; Hutchinson, M.; Hours, M.M.; Cortinovis-Tourniaire, P.; Moreau, T. Rate of pregnancy-related relapse in multiple sclerosis. Pregnancy in Multiple Sclerosis Group. N. Engl. J. Med. 1998, 339, 285–291. [Google Scholar] [CrossRef]
- Dendrou, C.A.; Fugger, L.; Friese, M.A. Immunopathology of multiple sclerosis. Nat. Rev. Immunol. 2015, 15, 545–558. [Google Scholar] [CrossRef]
- Voskuhl, R.R.; Gold, S.M. Sex-related factors in multiple sclerosis susceptibility and progression. Nat. Rev. Neurol. 2012, 8, 255–263. [Google Scholar] [CrossRef]
- Roux, T.; Courtillot, C.; Debs, R.; Touraine, P.; Lubetzki, C.; Papeix, C. Fecundity in women with multiple sclerosis: An observational mono-centric study. J. Neurol. 2015, 262, 957–960. [Google Scholar] [CrossRef]
- Arafa, A.; Wang, X.; Dong, J.Y.; Eshak, E.S. Does multiple sclerosis increase the risk of preeclampsia? A systematic review and meta-analysis. Hypertens Pregnancy 2021, 40, 180–185. [Google Scholar] [CrossRef]
- Cuello, J.P.; Martinez Gines, M.L.; Martin Barriga, M.L.; de Andres, C. Multiple sclerosis and pregnancy: A single-centre prospective comparative study. Neurologia 2017, 32, 92–98. [Google Scholar] [CrossRef]
- Arneth, B.M. Pregnancy in patients with multiple sclerosis. J. Investig. Med. 2022, 70, 14–19. [Google Scholar] [CrossRef]
- Graham, J.J.; Longhi, M.S.; Heneghan, M.A. T helper cell immunity in pregnancy and influence on autoimmune disease progression. J. Autoimmun. 2021, 121, 102651. [Google Scholar] [CrossRef]
- Clark, K.; Zhang, S.; Barthe, S.; Kumar, P.; Pivetti, C.; Kreutzberg, N.; Reed, C.; Wang, Y.; Paxton, Z.; Farmer, D.; et al. Placental Mesenchymal Stem Cell-Derived Extracellular Vesicles Promote Myelin Regeneration in an Animal Model of Multiple Sclerosis. Cells 2019, 8, 1497. [Google Scholar] [CrossRef] [Green Version]
- Martire, S.; Montarolo, F.; Spadaro, M.; Perga, S.; Sforza, M.L.; Marozio, L.; Frezet, F.; Bruno, S.; Chiabotto, G.; Deregibus, M.C.; et al. A First Phenotypic and Functional Characterization of Placental Extracellular Vesicles from Women with Multiple Sclerosis. Int. J. Mol. Sci. 2021, 22, 2875. [Google Scholar] [CrossRef]
- Landek-Salgado, M.A.; Rose, N.R.; Caturegli, P. Placenta suppresses experimental autoimmune hypophysitis through soluble TNF receptor 1. J. Autoimmun. 2012, 38, J88–J96. [Google Scholar] [CrossRef] [Green Version]
- Petraglia, F.; Florio, P.; Nappi, C.; Genazzani, A.R. Peptide signaling in human placenta and membranes: Autocrine, paracrine, and endocrine mechanisms. Endocr. Rev. 1996, 17, 156–186. [Google Scholar] [CrossRef] [Green Version]
- Napso, T.; Yong, H.E.J.; Lopez-Tello, J.; Sferruzzi-Perri, A.N. The Role of Placental Hormones in Mediating Maternal Adaptations to Support Pregnancy and Lactation. Front. Physiol. 2018, 9, 1091. [Google Scholar] [CrossRef]
- Wataganara, T.; Bianchi, D.W. Fetal cell-free nucleic acids in the maternal circulation: New clinical applications. Ann. N. Y. Acad. Sci. 2004, 1022, 90–99. [Google Scholar] [CrossRef]
- Nair, S.; Salomon, C. Extracellular vesicles as critical mediators of maternal-fetal communication during pregnancy and their potential role in maternal metabolism. Placenta 2020, 98, 60–68. [Google Scholar] [CrossRef]
- Phillips, T.A.; Ni, J.; Hunt, J.S. Death-inducing tumour necrosis factor (TNF) superfamily ligands and receptors are transcribed in human placentae, cytotrophoblasts, placental macrophages and placental cell lines. Placenta 2001, 22, 663–672. [Google Scholar] [CrossRef]
- Simonet, W.S.; Lacey, D.L.; Dunstan, C.R.; Kelley, M.; Chang, M.S.; Luthy, R.; Nguyen, H.Q.; Wooden, S.; Bennett, L.; Boone, T.; et al. Osteoprotegerin: A novel secreted protein involved in the regulation of bone density. Cell 1997, 89, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Walsh, M.C.; Choi, Y. Biology of the RANKL-RANK-OPG System in Immunity, Bone, and Beyond. Front. Immunol. 2014, 5, 511. [Google Scholar] [CrossRef] [Green Version]
- Ono, T.; Hayashi, M.; Sasaki, F.; Nakashima, T. RANKL biology: Bone metabolism, the immune system, and beyond. Inflamm. Regen. 2020, 40, 2. [Google Scholar] [CrossRef] [Green Version]
- Ahern, E.; Smyth, M.J.; Dougall, W.C.; Teng, M.W.L. Roles of the RANKL-RANK axis in antitumour immunity—Implications for therapy. Nat. Rev. Clin. Oncol. 2018, 15, 676–693. [Google Scholar] [CrossRef]
- Lleo, A.; Gershwin, M.E. Targeting the RANK/RANKL pathway in autoimmune disease and malignancy: Future perspectives. Expert Rev. Clin. Immunol. 2021, 17, 933–936. [Google Scholar] [CrossRef]
- Glasnovic, A.; O’Mara, N.; Kovacic, N.; Grcevic, D.; Gajovic, S. RANK/RANKL/OPG Signaling in the Brain: A Systematic Review of the Literature. Front. Neurol. 2020, 11, 590480. [Google Scholar] [CrossRef]
- Caniggia, I.; Mostachfi, H.; Winter, J.; Gassmann, M.; Lye, S.J.; Kuliszewski, M.; Post, M. Hypoxia-inducible factor-1 mediates the biological effects of oxygen on human trophoblast differentiation through TGFbeta(3). J. Clin. Investig. 2000, 105, 577–587. [Google Scholar] [CrossRef] [Green Version]
- Schock, H.; Zeleniuch-Jacquotte, A.; Lundin, E.; Grankvist, K.; Lakso, H.A.; Idahl, A.; Lehtinen, M.; Surcel, H.M.; Fortner, R.T. Hormone concentrations throughout uncomplicated pregnancies: A longitudinal study. BMC Pregnancy Childbirth 2016, 16, 146. [Google Scholar] [CrossRef]
- Shang, W.Q.; Li, H.; Liu, L.B.; Chang, K.K.; Yu, J.J.; Xie, F.; Li, M.Q.; Yu, J.J. RANKL/RANK interaction promotes the growth of cervical cancer cells by strengthening the dialogue between cervical cancer cells and regulation of IL-8 secretion. Oncol. Rep. 2015, 34, 3007–3016. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.S.; Santolaya-Forgas, J.; Romero, R.; Espinoza, J.; Goncalves, L.F.; Kim, Y.M.; Edwin, S.; Yoon, B.H.; Nien, J.K.; Hassan, S.; et al. Maternal plasma osteoprotegerin concentration in normal pregnancy. Am. J. Obstet. Gynecol. 2005, 193, 1011–1015. [Google Scholar] [CrossRef] [Green Version]
- Carmona-Fernandes, D.; Santos, M.J.; Perpetuo, I.P.; Fonseca, J.E.; Canhao, H. Soluble receptor activator of nuclear factor kappaB ligand/osteoprotegerin ratio is increased in systemic lupus erythematosus patients. Arthritis Res. Ther. 2011, 13, R175. [Google Scholar] [CrossRef] [Green Version]
- Jauniaux, E.; Watson, A.L.; Hempstock, J.; Bao, Y.P.; Skepper, J.N.; Burton, G.J. Onset of maternal arterial blood flow and placental oxidative stress. A possible factor in human early pregnancy failure. Am. J. Pathol. 2000, 157, 2111–2122. [Google Scholar] [CrossRef]
- Tomoyasu, A.; Goto, M.; Fujise, N.; Mochizuki, S.; Yasuda, H.; Morinaga, T.; Tsuda, E.; Higashio, K. Characterization of monomeric and homodimeric forms of osteoclastogenesis inhibitory factor. Biochem. Biophys. Res. Commun. 1998, 245, 382–387. [Google Scholar] [CrossRef]
- Nelson, C.A.; Warren, J.T.; Wang, M.W.; Teitelbaum, S.L.; Fremont, D.H. RANKL employs distinct binding modes to engage RANK and the osteoprotegerin decoy receptor. Structure 2012, 20, 1971–1982. [Google Scholar] [CrossRef] [Green Version]
- Hughes, C.E.; Nibbs, R.J.B. A guide to chemokines and their receptors. FEBS J. 2018, 285, 2944–2971. [Google Scholar] [CrossRef]
- Nakashima, T.; Takayanagi, H. Osteoimmunology: Crosstalk between the immune and bone systems. J. Clin. Immunol. 2009, 29, 555–567. [Google Scholar] [CrossRef]
- Walsh, M.C.; Choi, Y. Regulation of T cell-associated tissues and T cell activation by RANKL-RANK-OPG. J. Bone Miner. Metab. 2021, 39, 54–63. [Google Scholar] [CrossRef]
- Fata, J.E.; Kong, Y.Y.; Li, J.; Sasaki, T.; Irie-Sasaki, J.; Moorehead, R.A.; Elliott, R.; Scully, S.; Voura, E.B.; Lacey, D.L.; et al. The osteoclast differentiation factor osteoprotegerin-ligand is essential for mammary gland development. Cell 2000, 103, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Hanada, R.; Leibbrandt, A.; Hanada, T.; Kitaoka, S.; Furuyashiki, T.; Fujihara, H.; Trichereau, J.; Paolino, M.; Qadri, F.; Plehm, R.; et al. Central control of fever and female body temperature by RANKL/RANK. Nature 2009, 462, 505–509. [Google Scholar] [CrossRef]
- Darnay, B.G.; Haridas, V.; Ni, J.; Moore, P.A.; Aggarwal, B.B. Characterization of the intracellular domain of receptor activator of NF-kappaB (RANK). Interaction with tumor necrosis factor receptor-associated factors and activation of NF-kappab and c-Jun N-terminal kinase. J. Biol. Chem. 1998, 273, 20551–20555. [Google Scholar] [CrossRef] [Green Version]
- Seshasayee, D.; Wang, H.; Lee, W.P.; Gribling, P.; Ross, J.; Van Bruggen, N.; Carano, R.; Grewal, I.S. A novel in vivo role for osteoprotegerin ligand in activation of monocyte effector function and inflammatory response. J. Biol. Chem. 2004, 279, 30202–30209. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Li, M.; Larocque, R.; Zhang, F.; Malhotra, A.; Chen, J.; Linhardt, R.J.; Konermann, L.; Xu, D. Dimerization interface of osteoprotegerin revealed by hydrogen-deuterium exchange mass spectrometry. J. Biol. Chem. 2018, 293, 17523–17535. [Google Scholar] [CrossRef] [Green Version]
- Papadaki, M.; Rinotas, V.; Violitzi, F.; Thireou, T.; Panayotou, G.; Samiotaki, M.; Douni, E. New Insights for RANKL as a Proinflammatory Modulator in Modeled Inflammatory Arthritis. Front. Immunol. 2019, 10, 97. [Google Scholar] [CrossRef] [Green Version]
- Gregorczyk, I.; Jasiecka-Mikolajczyk, A.; Maslanka, T. Blockade of NF-kappaB Translocation and of RANKL/RANK Interaction Decreases the Frequency of Th2 and Th17 Cells Capable of IL-4 and IL-17 Production, Respectively, in a Mouse Model of Allergic Asthma. Molecules 2021, 26, 3117. [Google Scholar] [CrossRef]
- Glasnovic, A.; Stojic, M.; Dezmalj, L.; Tudoric-Deno, I.; Romic, D.; Jelec, V.; Vrca, A.; Vuletic, V.; Grcevic, D. RANKL/RANK/OPG Axis Is Deregulated in the Cerebrospinal Fluid of Multiple Sclerosis Patients at Clinical Onset. Neuroimmunomodulation 2018, 25, 23–33. [Google Scholar] [CrossRef]
- Lacey, D.L.; Boyle, W.J.; Simonet, W.S.; Kostenuik, P.J.; Dougall, W.C.; Sullivan, J.K.; San Martin, J.; Dansey, R. Bench to bedside: Elucidation of the OPG-RANK-RANKL pathway and the development of denosumab. Nat. Rev. Drug Discov. 2012, 11, 401–419. [Google Scholar] [CrossRef]
- Hofbauer, L.C.; Khosla, S.; Dunstan, C.R.; Lacey, D.L.; Boyle, W.J.; Riggs, B.L. The roles of osteoprotegerin and osteoprotegerin ligand in the paracrine regulation of bone resorption. J. Bone Miner. Res. 2000, 15, 2–12. [Google Scholar] [CrossRef]
- Sintzel, M.B.; Rametta, M.; Reder, A.T. Vitamin D and Multiple Sclerosis: A Comprehensive Review. Neurol Ther. 2018, 7, 59–85. [Google Scholar] [CrossRef] [Green Version]
- Burton, G.J.; Fowden, A.L. The placenta: A multifaceted, transient organ. Philos. Trans. R Soc. Lond. B Biol. Sci. 2015, 370, 20140066. [Google Scholar] [CrossRef] [Green Version]
- Spadaro, M.; Martire, S.; Marozio, L.; Mastromauro, D.; Montanari, E.; Perga, S.; Montarolo, F.; Brescia, F.; Balbo, A.; Botta, G.; et al. Immunomodulatory Effect of Pregnancy on Leukocyte Populations in Patients With Multiple Sclerosis: A Comparison of Peripheral Blood and Decidual Placental Tissue. Front. Immunol. 2019, 10, 1935. [Google Scholar] [CrossRef] [Green Version]
- Schneeweis, L.A.; Willard, D.; Milla, M.E. Functional dissection of osteoprotegerin and its interaction with receptor activator of NF-kappaB ligand. J. Biol. Chem. 2005, 280, 41155–41164. [Google Scholar] [CrossRef] [Green Version]
- Yano, K.; Shibata, O.; Mizuno, A.; Kobayashi, F.; Higashio, K.; Morinaga, T.; Tsuda, E. Immunological study on circulating murine osteoprotegerin/osteoclastogenesis inhibitory factor (OPG/OCIF): Possible role of OPG/OCIF in the prevention of osteoporosis in pregnancy. Biochem. Biophys. Res. Commun. 2001, 288, 217–224. [Google Scholar] [CrossRef]
- Yano, K.; Tsuda, E.; Washida, N.; Kobayashi, F.; Goto, M.; Harada, A.; Ikeda, K.; Higashio, K.; Yamada, Y. Immunological characterization of circulating osteoprotegerin/osteoclastogenesis inhibitory factor: Increased serum concentrations in postmenopausal women with osteoporosis. J. Bone Miner. Res. 1999, 14, 518–527. [Google Scholar] [CrossRef]
- Hofbauer, L.C.; Khosla, S.; Dunstan, C.R.; Lacey, D.L.; Spelsberg, T.C.; Riggs, B.L. Estrogen stimulates gene expression and protein production of osteoprotegerin in human osteoblastic cells. Endocrinology 1999, 140, 4367–4370. [Google Scholar] [CrossRef]
- Ysrraelit, M.C.; Correale, J. Impact of sex hormones on immune function and multiple sclerosis development. Immunology 2019, 156, 9–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berkane, N.; Liere, P.; Oudinet, J.P.; Hertig, A.; Lefevre, G.; Pluchino, N.; Schumacher, M.; Chabbert-Buffet, N. From Pregnancy to Preeclampsia: A Key Role for Estrogens. Endocr. Rev. 2017, 38, 123–144. [Google Scholar] [CrossRef] [PubMed]
- Guerrini, M.M.; Okamoto, K.; Komatsu, N.; Sawa, S.; Danks, L.; Penninger, J.M.; Nakashima, T.; Takayanagi, H. Inhibition of the TNF Family Cytokine RANKL Prevents Autoimmune Inflammation in the Central Nervous System. Immunity 2015, 43, 1174–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemmer, B.; Kerschensteiner, M.; Korn, T. Role of the innate and adaptive immune responses in the course of multiple sclerosis. Lancet Neurol 2015, 14, 406–419. [Google Scholar] [CrossRef]
- Johnson, A.C.; Cipolla, M.J. The cerebral circulation during pregnancy: Adapting to preserve normalcy. Physiology 2015, 30, 139–147. [Google Scholar] [CrossRef]
- Hawkins, C.P.; Munro, P.M.; MacKenzie, F.; Kesselring, J.; Tofts, P.S.; du Boulay, E.P.; Landon, D.N.; McDonald, W.I. Duration and selectivity of blood-brain barrier breakdown in chronic relapsing experimental allergic encephalomyelitis studied by gadolinium-DTPA and protein markers. Brain 1990, 113, 365–378. [Google Scholar] [CrossRef]
- Mony, J.T.; Khorooshi, R.; Owens, T. Chemokine receptor expression by inflammatory T cells in EAE. Front. Cell Neurosci. 2014, 8, 187. [Google Scholar] [CrossRef] [Green Version]
- Reboldi, A.; Coisne, C.; Baumjohann, D.; Benvenuto, F.; Bottinelli, D.; Lira, S.; Uccelli, A.; Lanzavecchia, A.; Engelhardt, B.; Sallusto, F. C-C chemokine receptor 6-regulated entry of TH-17 cells into the CNS through the choroid plexus is required for the initiation of EAE. Nat. Immunol. 2009, 10, 514–523. [Google Scholar] [CrossRef]
- Moser, T.; Akgun, K.; Proschmann, U.; Sellner, J.; Ziemssen, T. The role of TH17 cells in multiple sclerosis: Therapeutic implications. Autoimmun. Rev. 2020, 19, 102647. [Google Scholar] [CrossRef]
- Jafarzadeh, A.; Bagherzadeh, S.; Ebrahimi, H.A.; Hajghani, H.; Bazrafshani, M.R.; Khosravimashizi, A.; Nemati, M.; Gadari, F.; Sabahi, A.; Iranmanesh, F.; et al. Higher circulating levels of chemokine CCL20 in patients with multiple sclerosis: Evaluation of the influences of chemokine gene polymorphism, gender, treatment and disease pattern. J. Mol. Neurosci. 2014, 53, 500–505. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Data | Not Pregnant (n = 10) | First-Trimester (n = 10) | Term (n = 20) |
|---|---|---|---|
| Maternal age | 35.5 ± 2.0 | 28.2 ± 6.8 | 37.4 ± 9.0 |
| Caucasian ethnicity (%) | - | 100 | 100 |
| Pre-pregnancy weight [mean (SD)] | - | - | 64 ± 10.7 |
| Weight gain (kg) [mean (SD)] | - | - | 10.9 ± 8.8 |
| Gestational age (weeks) [mean (SD)] | - | 9 ± 2 | 38 ± 0.6 |
| Nulliparous (%) | 21 | - | 23.2 |
| Female (%) | - | - | 77 |
| Male (%) | - | - | 23 |
| Placental weight (g) [mean (SD)] | - | 12.3 ± 3.5 | 449.8 ± 83.6 |
| Birth weight (g) [mean (SD)] | - | - | 2879.1 ± 581 |
| Clinical Data | MS Women Pregnant (n = 10) | MS Women Non-Pregnant (n = 19) |
|---|---|---|
| Mean age (years) [mean (SD)] | 34.9 ± 4.7 | 33.0 ± 8.4 |
| Median number of relapses before pregnancy [median (range)] | 2 (0–2.6) | - |
| Median number of relapses after delivery [median (range)] | 0 (2–0) | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Passaponti, S.; Ermini, L.; Acconci, G.; Severi, F.M.; Romagnoli, R.; Cutrupi, S.; Clerico, M.; Guerrera, G.; Ietta, F. Rank-Rankl-Opg Axis in Multiple Sclerosis: The Contribution of Placenta. Cells 2022, 11, 1357. https://doi.org/10.3390/cells11081357
Passaponti S, Ermini L, Acconci G, Severi FM, Romagnoli R, Cutrupi S, Clerico M, Guerrera G, Ietta F. Rank-Rankl-Opg Axis in Multiple Sclerosis: The Contribution of Placenta. Cells. 2022; 11(8):1357. https://doi.org/10.3390/cells11081357
Chicago/Turabian StylePassaponti, Sofia, Leonardo Ermini, Giulia Acconci, Filiberto Maria Severi, Roberta Romagnoli, Santina Cutrupi, Marinella Clerico, Gisella Guerrera, and Francesca Ietta. 2022. "Rank-Rankl-Opg Axis in Multiple Sclerosis: The Contribution of Placenta" Cells 11, no. 8: 1357. https://doi.org/10.3390/cells11081357