
Emerging Evidence of Noncoding RNAs in Bleb Scarring after Glaucoma Filtration Surgery



Abstract
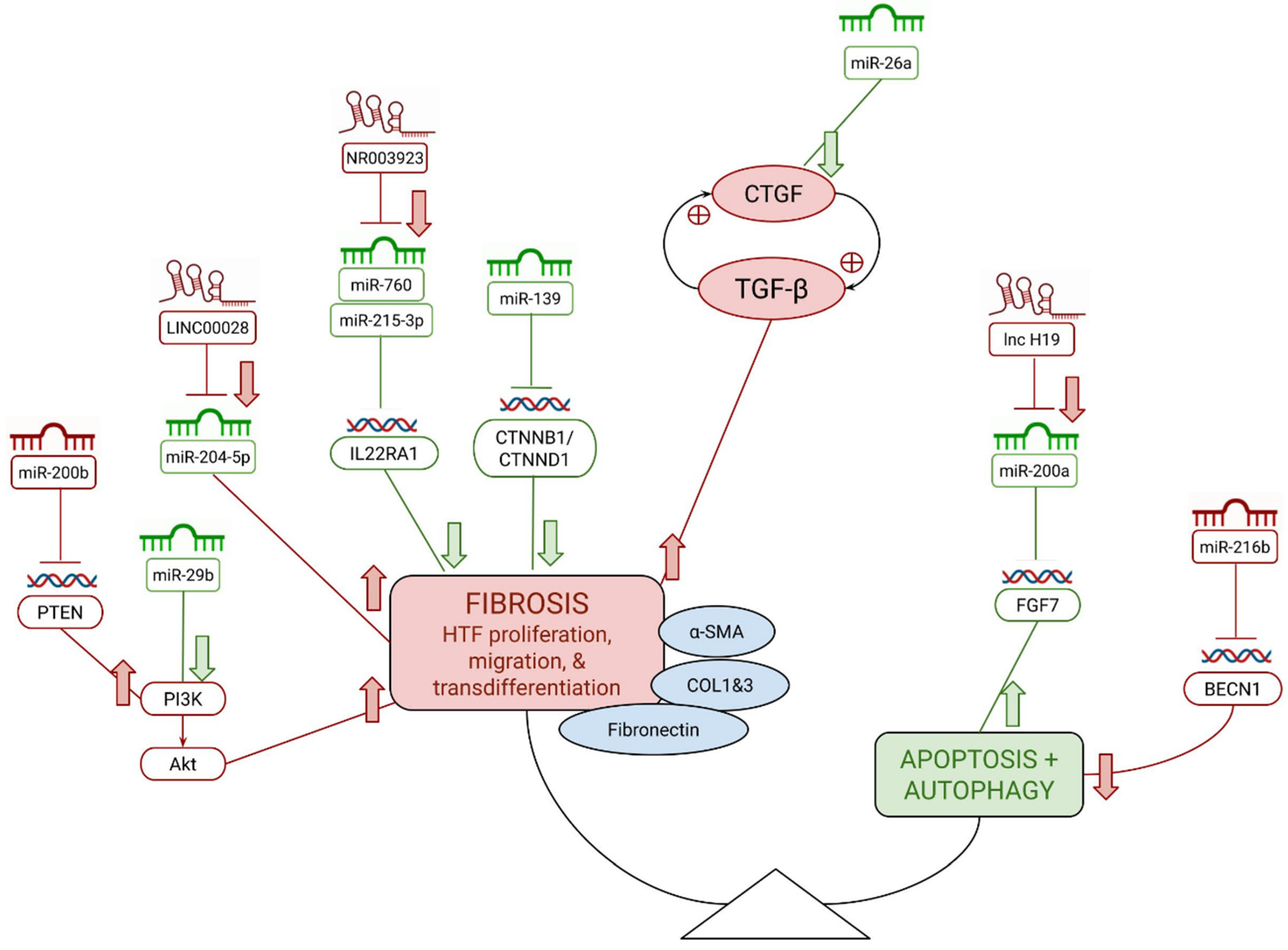
:1. Introduction
2. Methods
3. Results and Discussion
3.1. microRNAs
3.2. Antifibrotic miRNAs
3.2.1. miR-26a
3.2.2. miR-29b
3.2.3. miR-139
3.2.4. miR-200a
3.3. Pro-Fibrotic miRNAs
3.3.1. miRNA-200b
3.3.2. miR-200c
3.3.3. miR-216b
3.4. Long Noncoding RNAs
3.5. Pro-Fibrotic lncRNAs
3.5.1. H19
3.5.2. NR003923
3.5.3. LINC00028
4. Conclusions

{kind=link}
| Noncoding RNA | Studies | Pro/Anti-Fibrotic Role |
|---|---|---|
| miR-26a | Wang, Deng, and He, 2018 [30] Bao et al., 2018 [26] | Anti-fibrotic |
| miR-29b | Li et al., 2012 [38] Ran, Zhu, and Feng, 2015 [29] | Anti-fibrotic |
| miR-139 | Deng et al., 2019 [27] | Anti-fibrotic |
| miR-200a | Peng et al., 2019 [28] | Anti-fibrotic |
| miR-200b | Tong et al., 2014 [37] Tong et al., 2019 [32] | Pro-fibrotic |
| miR-216b | Xu et al., 2014 [33] | Pro-fibrotic |
| lnc H19 | Zhu et al., 2020 [21] | Pro-fibrotic |
| lnc NR003923 | Zhao et al., 2019 [47] | Pro-fibrotic |
| LINC00028 | Sui et al., 2020 [45] | Pro-fibrotic |
Author Contributions
Funding
Conflicts of Interest
References
- Allison, K.; Patel, D.; Alabi, O. Epidemiology of Glaucoma: The Past, Present, and Predictions for the Future. Cureus 2020, 12, e11686. [Google Scholar] [CrossRef]
- Chen, T.C.; Wilensky, J.T.; Viana, M.A. Long-Term Follow-up of Initially Successful Trabeculectomy. Ophthalmology 1997, 104, 1120–1125. [Google Scholar] [CrossRef]
- Gedde, S.J.; Schiffman, J.C.; Feuer, W.J.; Herndon, L.W.; Brandt, J.D.; Budenz, D.L. Tube versus Trabeculectomy Study Group. Treatment Outcomes in the Tube Versus Trabeculectomy (TVT) Study after Five Years of Follow-Up. Am. J. Ophthalmol. 2012, 153, 789–803. [Google Scholar] [CrossRef] [Green Version]
- Cabourne, E.; Clarke, J.C.K.; Schlottmann, P.G.; Evans, J.R. Mitomycin C versus 5-Fluorouracil for Wound Healing in Glaucoma Surgery. Cochrane Database Syst. Rev. 2015, 2015, CD006259. [Google Scholar] [CrossRef] [PubMed]
- Fan Gaskin, J.C.; Nguyen, D.Q.; Soon Ang, G.; O’Connor, J.; Crowston, J.G. Wound Healing Modulation in Glaucoma Filtration Surgery–Conventional Practices and New Perspectives: The Role of Antifibrotic Agents (Part I). J. Curr. Glaucoma Pract. 2014, 8, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Kirwan, J.F.; Lockwood, A.J.; Shah, P.; Macleod, A.; Broadway, D.C.; King, A.J.; McNaught, A.I.; Agrawal, P.; Trabeculectomy Outcomes Group Audit Study Group. Trabeculectomy in the 21st Century: A Multicenter Analysis. Ophthalmology 2013, 120, 2532–2539. [Google Scholar] [CrossRef] [PubMed]
- Lukowski, Z.L.; Min, J.; Beattie, A.R.; Meyers, C.A.; Levine, M.A.; Stoller, G.; Schultz, G.S.; Samuelson, D.A.; Sherwood, M.B. Prevention of Ocular Scarring After Glaucoma Filtering Surgery Using the Monoclonal Antibody LT1009 (Sonepcizumab) in a Rabbit Model. J. Glaucoma 2013, 22, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Perez, C.I.; Mellado, F.; Jones, A.; Colvin, R. Trabeculectomy Combined with Collagen Matrix Implant (Ologen). J. Glaucoma 2017, 26, 54–58. [Google Scholar] [CrossRef]
- Song, D.-S.; Qian, J.; Chen, Z.-J. Ologen Implant versus Mitomycin-C for Trabeculectomy: A Meta-Analysis. Medicine 2019, 98, e16094. [Google Scholar] [CrossRef]
- Liu, X.; Du, L.; Li, N. The Effects of Bevacizumab in Augmenting Trabeculectomy for Glaucoma: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Medicine 2016, 95, e3223. [Google Scholar] [CrossRef]
- Molasy, M.; Walczak, A.; Szaflik, J.; Szaflik, J.P.; Majsterek, I. MicroRNAs in Glaucoma and Neurodegenerative Diseases. J. Hum. Genet. 2017, 62, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef] [Green Version]
- Kapranov, P.; Cheng, J.; Dike, S.; Nix, D.A.; Duttagupta, R.; Willingham, A.T.; Stadler, P.F.; Hertel, J.; Hackermüller, J.; Hofacker, I.L.; et al. RNA Maps Reveal New RNA Classes and a Possible Function for Pervasive Transcription. Science 2007, 316, 1484–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayed, D.; Abdellatif, M. MicroRNAs in Development and Disease. Physiol. Rev. 2011, 91, 827–887. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The MicroRNA Spectrum in 12 Body Fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef]
- Jayaram, H.; Phillips, J.I.; Lozano, D.C.; Choe, T.E.; Cepurna, W.O.; Johnson, E.C.; Morrison, J.C.; Gattey, D.M.; Saugstad, J.A.; Keller, K.E. Comparison of MicroRNA Expression in Aqueous Humor of Normal and Primary Open-Angle Glaucoma Patients Using PCR Arrays: A Pilot Study. Investig. Ophthalmol. Vis. Sci. 2017, 58, 2884–2890. [Google Scholar] [CrossRef] [Green Version]
- Xu, S. MicroRNA Expression in the Eyes and Their Significance in Relation to Functions. Prog. Retin. Eye Res. 2009, 28, 87–116. [Google Scholar] [CrossRef]
- Zhang, L.; Dong, Y.; Wang, Y.; Gao, J.; Lv, J.; Sun, J.; Li, M.; Wang, M.; Zhao, Z.; Wang, J.; et al. Long Non-Coding RNAs in Ocular Diseases: New and Potential Therapeutic Targets. FEBS J. 2019, 286, 2261–2272. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wang, W.; Zhu, W.; Dong, J.; Cheng, Y.; Yin, Z.; Shen, F. Mechanisms and Functions of Long Non-Coding RNAs at Multiple Regulatory Levels. Int. J. Mol. Sci. 2019, 20, 5573. [Google Scholar] [CrossRef] [Green Version]
- Gajda-Deryło, B.; Stahnke, T.; Struckmann, S.; Warsow, G.; Birke, K.; Birke, M.T.; Hohberger, B.; Rejdak, R.; Fuellen, G.; Jünemann, A.G. Comparison of Cytokine/Chemokine Levels in Aqueous Humor of Primary Open-Angle Glaucoma Patients with Positive or Negative Outcome Following Trabeculectomy. Biosci. Rep. 2019, 39, BSR20181894. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Dai, L.; Li, X.; Zhang, Z.; Liu, Y.; Quan, F.; Zhang, P.; Yu, L. Role of the Long Noncoding RNA H19 in TGF-Β1-Induced Tenon’s Capsule Fibroblast Proliferation and Extracellular Matrix Deposition. Exp. Cell Res. 2020, 387, 111802. [Google Scholar] [CrossRef] [PubMed]
- Kok, H.M.; Falke, L.L.; Goldschmeding, R.; Nguyen, T.Q. Targeting CTGF, EGF and PDGF Pathways to Prevent Progression of Kidney Disease. Nat. Rev. Nephrol. 2014, 10, 700–711. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, D.G.; Ko, J.-A.; Iwata, W.; Okumichi, H.; Kiuchi, Y. An in Vitro Study of Scarring Formation Mediated by Human Tenon Fibroblasts: Effect of Y-27632, a Rho Kinase Inhibitor. Cell Biochem. Funct. 2019, 37, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Mietzner, R.; Breunig, M. Causative Glaucoma Treatment: Promising Targets and Delivery Systems. Drug Discov. Today 2019, 24, 1606–1613. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, O.; Kitano-Izutani, A.; Tomoyose, K.; Reinach, P.S. Pathobiology of Wound Healing after Glaucoma Filtration Surgery. BMC Ophthalmol. 2015, 15 (Suppl. 1), 157. [Google Scholar] [CrossRef] [Green Version]
- Bao, H.; Jiang, K.; Meng, K.; Liu, W.; Liu, P.; Du, Y.; Wang, D. TGF-Β2 Induces Proliferation and Inhibits Apoptosis of Human Tenon Capsule Fibroblast by MiR-26 and Its Targeting of CTGF. Biomed. Pharmacother. Biomed. Pharmacother. 2018, 104, 558–565. [Google Scholar] [CrossRef]
- Deng, M.; Hou, S.-Y.; Tong, B.-D.; Yin, J.-Y.; Xiong, W. The Smad2/3/4 Complex Binds MiR-139 Promoter to Modulate TGFβ-Induced Proliferation and Activation of Human Tenon’s Capsule Fibroblasts through the Wnt Pathway. J. Cell. Physiol. 2019, 234, 13342–13352. [Google Scholar] [CrossRef]
- Peng, H.; Sun, Y.-B.; Hao, J.-L.; Lu, C.-W.; Bi, M.-C.; Song, E. Neuroprotective Effects of Overexpressed MicroRNA-200a on Activation of Glaucoma-Related Retinal Glial Cells and Apoptosis of Ganglion Cells via Downregulating FGF7-Mediated MAPK Signaling Pathway. Cell. Signal. 2019, 54, 179–190. [Google Scholar] [CrossRef]
- Ran, W.; Zhu, D.; Feng, Q. TGF-Β2 Stimulates Tenon’s Capsule Fibroblast Proliferation in Patients with Glaucoma via Suppression of MiR-29b Expression Regulated by Nrf2. Int. J. Clin. Exp. Pathol. 2015, 8, 4799–4806. [Google Scholar]
- Wang, W.-H.; Deng, A.-J.; He, S.-G. A Key Role of MicroRNA-26a in the Scar Formation after Glaucoma Filtration Surgery. Artif. Cells Nanomed. Biotechnol. 2018, 46, 831–837. [Google Scholar] [CrossRef] [Green Version]
- Eissa, M.G.; Artlett, C.M. The MicroRNA MiR-155 Is Essential in Fibrosis. Non-Coding RNA 2019, 5, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, J.; Chen, F.; Du, W.; Zhu, J.; Xie, Z. TGF-Β1 Induces Human Tenon’s Fibroblasts Fibrosis via MiR-200b and Its Suppression of PTEN Signaling. Curr. Eye Res. 2019, 44, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Fu, Y.; Tong, J.; Fan, S.; Xu, K.; Sun, H.; Liang, Y.; Yan, C.; Yuan, Z.; Ge, Y. MicroRNA-216b/Beclin 1 Axis Regulates Autophagy and Apoptosis in Human Tenon’s Capsule Fibroblasts upon Hydroxycamptothecin Exposure. Exp. Eye Res. 2014, 123, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Drewry, M.D.; Challa, P.; Kuchtey, J.G.; Navarro, I.; Helwa, I.; Hu, Y.; Mu, H.; Stamer, W.D.; Kuchtey, R.W.; Liu, Y. Differentially Expressed MicroRNAs in the Aqueous Humor of Patients with Exfoliation Glaucoma or Primary Open-Angle Glaucoma. Hum. Mol. Genet. 2018, 27, 1263–1275. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Xiao, W.; Chen, W.; Liu, X.; Wu, M.; Bo, Q.; Luo, Y.; Ye, S.; Cao, Y.; Liu, Y. MicroRNA-26a and -26b Inhibit Lens Fibrosis and Cataract by Negatively Regulating Jagged-1/Notch Signaling Pathway. Cell Death Differ. 2017, 24, 1431–1442. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Lin, H.; Wang, Q.; Chen, W.; Luo, H.; Chen, W.; Zhang, H. Discrepant Expression of MicroRNAs in Transparent and Cataractous Human Lenses. Investig. Ophthalmol. Vis. Sci. 2012, 53, 3906–3912. [Google Scholar] [CrossRef] [Green Version]
- Tong, J.; Fu, Y.; Xu, X.; Fan, S.; Sun, H.; Liang, Y.; Xu, K.; Yuan, Z.; Ge, Y. TGF-Β1 Stimulates Human Tenon’s Capsule Fibroblast Proliferation by MiR-200b and Its Targeting of P27/Kip1 and RND3. Investig. Ophthalmol. Vis. Sci. 2014, 55, 2747–2756. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Cui, J.; Duan, X.; Chen, H.; Fan, F. Suppression of Type I Collagen Expression by MiR-29b via PI3K, Akt, and Sp1 Pathway in Human Tenon’s Fibroblasts. Investig. Ophthalmol. Vis. Sci. 2012, 53, 1670–1678. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Luo, H.; Li, N.; Duan, X. Suppression of Type I Collagen Expression by MiR-29b Via PI3K, Akt, and Sp1 Pathway, Part II: An In Vivo Investigation. Investig. Ophthalmol. Vis. Sci. 2015, 56, 6019–6028. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Chen, M.; Zeng, L.; Liu, L. Integrated Aqueous Humor CeRNA and MiRNA-TF-MRNA Network Analysis Reveals Potential Molecular Mechanisms Governing Primary Open-Angle Glaucoma Pathogenesis. bioRxiv 2020. [Google Scholar] [CrossRef]
- Vallée, A.; Lecarpentier, Y.; Guillevin, R.; Vallée, J.-N. Interactions between TGF-Β1, Canonical WNT/β-Catenin Pathway and PPAR γ in Radiation-Induced Fibrosis. Oncotarget 2017, 8, 90579–90604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, L.; Browne, G.; Tulchinsky, E. ZEB/MiR-200 Feedback Loop: At the Crossroads of Signal Transduction in Cancer. Int. J. Cancer 2013, 132, 745–754. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, W.; Jiang, Y.; A-Tuya; Bai, D.; Li, L.; Lu, Z.-J.; Chang, H.; Zhang, T.-Z. GRGM-13 Comprising 13 Plant and Animal Products, Inhibited Oxidative Stress Induced Apoptosis in Retinal Ganglion Cells by Inhibiting P2RX7/P38 MAPK Signaling Pathway. Biomed. Pharmacother. 2018, 101, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Luna, C.; Li, G.; Huang, J.; Qiu, J.; Wu, J.; Yuan, F.; Epstein, D.L.; Gonzalez, P. Regulation of Trabecular Meshwork Cell Contraction and Intraocular Pressure by MiR-200c. PLoS ONE 2012, 7, e51688. [Google Scholar] [CrossRef] [PubMed]
- Sui, H.; Fan, S.; Liu, W.; Li, Y.; Zhang, X.; Du, Y.; Bao, H. LINC00028 Regulates the Development of TGFβ1-Treated Human Tenon Capsule Fibroblasts by Targeting MiR-204-5p. Biochem. Biophys. Res. Commun. 2020, 525, 197–203. [Google Scholar] [CrossRef]
- Cai, X.; Cullen, B.R. The Imprinted H19 Noncoding RNA Is a Primary MicroRNA Precursor. RNA 2007, 13, 313–316. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zhang, F.; Pan, Z.; Luo, H.; Liu, K.; Duan, X. LncRNA NR_003923 Promotes Cell Proliferation, Migration, Fibrosis, and Autophagy via the MiR-760/MiR-215-3p/IL22RA1 Axis in Human Tenon’s Capsule Fibroblasts. Cell Death Dis. 2019, 10, 594. [Google Scholar] [CrossRef]
- Kong, X.; Feng, D.; Mathews, S.; Gao, B. Hepatoprotective and Anti-Fibrotic Functions of Interleukin-22: Therapeutic Potential for the Treatment of Alcoholic Liver Disease. J. Gastroenterol. Hepatol. 2013, 28, 56–60. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Li, Y.; Fan, J.; Zhang, X.; Luan, J.; Bian, Q.; Ding, T.; Wang, Y.; Wang, Z.; Song, P.; et al. Interleukin-22 Ameliorated Renal Injury and Fibrosis in Diabetic Nephropathy through Inhibition of NLRP3 Inflammasome Activation. Cell Death Dis. 2017, 8, e2937. [Google Scholar] [CrossRef] [Green Version]
- Friedman, R.C.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most Mammalian MRNAs Are Conserved Targets of MicroRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Camarillo, C.; Swerdel, M.; Hart, R.P. Comparison of Microarray and Quantitative Real-Time PCR Methods for Measuring MicroRNA Levels in MSC Cultures. Methods Mol Bol. 2011, 698, 419–429. [Google Scholar] [CrossRef] [Green Version]
- Moldovan, L.; Batte, K.E.; Trgovcich, J.; Wisler, J.; Marsh, C.B.; Piper, M. Methodological Challenges in Utilizing MiRNAs as Circulating Biomarkers. J. Cell. Mol. Med. 2014, 18, 371–390. [Google Scholar] [CrossRef] [PubMed]
- Creighton, C.J.; Reid, J.G.; Gunaratne, P.H. Expression Profiling of MicroRNAs by Deep Sequencing. Brief. Bioinform. 2009, 10, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Romano, G.L.; Platania, C.B.M.; Drago, F.; Salomone, S.; Ragusa, M.; Barbagallo, C.; Di Pietro, C.; Purrello, M.; Reibaldi, M.; Avitabile, T.; et al. Retinal and Circulating MiRNAs in Age-Related Macular Degeneration: An In Vivo Animal and Human Study. Front. Pharmacol. 2017, 8, 168. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Xiao, X.; Wang, C.; Zhang, X.; Li, F.; Zhou, Y.; Kijlstra, A.; Yang, P. Decreased MicroRNA-155 Expression in Ocular Behcet’s Disease but Not in Vogt Koyanagi Harada Syndrome. Investig. Ophthalmol. Vis. Sci. 2012, 53, 5665–5674. [Google Scholar] [CrossRef] [Green Version]
- Ørom, U.A.; Kauppinen, S.; Lund, A.H. LNA-Modified Oligonucleotides Mediate Specific Inhibition of MicroRNA Function. Gene 2006, 372, 137–141. [Google Scholar] [CrossRef]
- Junn, E.; Mouradian, M.M. MicroRNAs in Neurodegenerative Diseases and Their Therapeutic Potential. Pharmacol. Ther. 2012, 133, 142–150. [Google Scholar] [CrossRef] [Green Version]
- Bonneau, E.; Neveu, B.; Kostantin, E.; Tsongalis, G.J.; De Guire, V. How Close Are MiRNAs from Clinical Practice? A Perspective on the Diagnostic and Therapeutic Market. EJIFCC 2019, 30, 114–127. [Google Scholar]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Lee, S.-S. Therapeutic Advances of MiRNAs: A Preclinical and Clinical Update. J. Adv. Res. 2021, 28, 127–138. [Google Scholar] [CrossRef]
- Pfeiffer, N.; Voykov, B.; Renieri, G.; Bell, K.; Richter, P.; Weigel, M.; Thieme, H.; Wilhelm, B.; Lorenz, K.; Feindor, M.; et al. First-in-Human Phase I Study of ISTH0036, an Antisense Oligonucleotide Selectively Targeting Transforming Growth Factor Beta 2 (TGF-Β2), in Subjects with Open-Angle Glaucoma Undergoing Glaucoma Filtration Surgery. PLoS ONE 2017, 12, e0188899. [Google Scholar] [CrossRef] [Green Version]
- CAT-152 0102 Trabeculectomy Study Group; Khaw, P.; Grehn, F.; Holló, G.; Overton, B.; Wilson, R.; Vogel, R.; Smith, Z. A Phase III Study of Subconjunctival Human Anti-Transforming Growth Factor Beta(2) Monoclonal Antibody (CAT-152) to Prevent Scarring after First-Time Trabeculectomy. Ophthalmology 2007, 114, 1822–1830. [Google Scholar] [CrossRef]
- Lee, S.Y.; Chae, M.K.; Yoon, J.S.; Kim, C.Y. The Effect of CHIR 99021, a Glycogen Synthase Kinase-3β Inhibitor, on Transforming Growth Factor β-Induced Tenon Fibrosis. Investig. Ophthalmol. Vis. Sci. 2021, 62, 25. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Li, Y.-J.; Liu, J.-Y.; Zhang, Y.-Y.; Li, X.-M.; Wang, L.-N.; Yao, J.; Jiang, Q.; Yan, B. Identification of Corneal Neovascularization-Related Long Noncoding RNAs through Microarray Analysis. Cornea 2015, 34, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Liu, S.; Lv, Y.; Zhang, C.; Gao, H.; Tan, L.; Wang, H. Long Non-Coding RNA HOTAIR Regulates Proliferation and Invasion via Activating Notch Signalling Pathway in Retinoblastoma. J. Biosci. 2016, 41, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-Y.; Yao, J.; Li, X.-M.; Song, Y.-C.; Wang, X.-Q.; Li, Y.-J.; Yan, B.; Jiang, Q. Pathogenic Role of LncRNA-MALAT1 in Endothelial Cell Dysfunction in Diabetes Mellitus. Cell Death Dis. 2014, 5, e1506. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Yu, Y.; Ni, X.; Li, C.; Hu, Y.; Wang, J.; Chen, F.; Xi, S.; Chen, Z. LncRNA LINC00998 Inhibits the Malignant Glioma Phenotype via the CBX3-Mediated c-Met/Akt/MTOR Axis. Cell Death Dis. 2020, 11, 1032. [Google Scholar] [CrossRef]
- Xie, L.; Mao, M.; Wang, C.; Zhang, L.; Pan, Z.; Shi, J.; Duan, X.; Jia, S.; Jiang, B. Potential Biomarkers for Primary Open-Angle Glaucoma Identified by Long Noncoding RNA Profiling in the Aqueous Humor. Am. J. Pathol. 2019, 189, 739–752. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Zheng, Y.; Gao, M.; Ma, H.; Zhang, X.; Li, Y.; Wang, F.; Huang, H. Expression and Clinical Value of LncRNA MALAT1 and LncRNA ANRIL in Glaucoma Patients. Exp. Ther. Med. 2020, 19, 1329–1335. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, S.; Tam, A.L.C.; Campbell, R.; Renwick, N. Emerging Evidence of Noncoding RNAs in Bleb Scarring after Glaucoma Filtration Surgery. Cells 2022, 11, 1301. https://doi.org/10.3390/cells11081301
Yu S, Tam ALC, Campbell R, Renwick N. Emerging Evidence of Noncoding RNAs in Bleb Scarring after Glaucoma Filtration Surgery. Cells. 2022; 11(8):1301. https://doi.org/10.3390/cells11081301
Chicago/Turabian StyleYu, Sabrina, Alex L. C. Tam, Robert Campbell, and Neil Renwick. 2022. "Emerging Evidence of Noncoding RNAs in Bleb Scarring after Glaucoma Filtration Surgery" Cells 11, no. 8: 1301. https://doi.org/10.3390/cells11081301