
Interactions between the Nociceptin and Toll-like Receptor Systems


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures
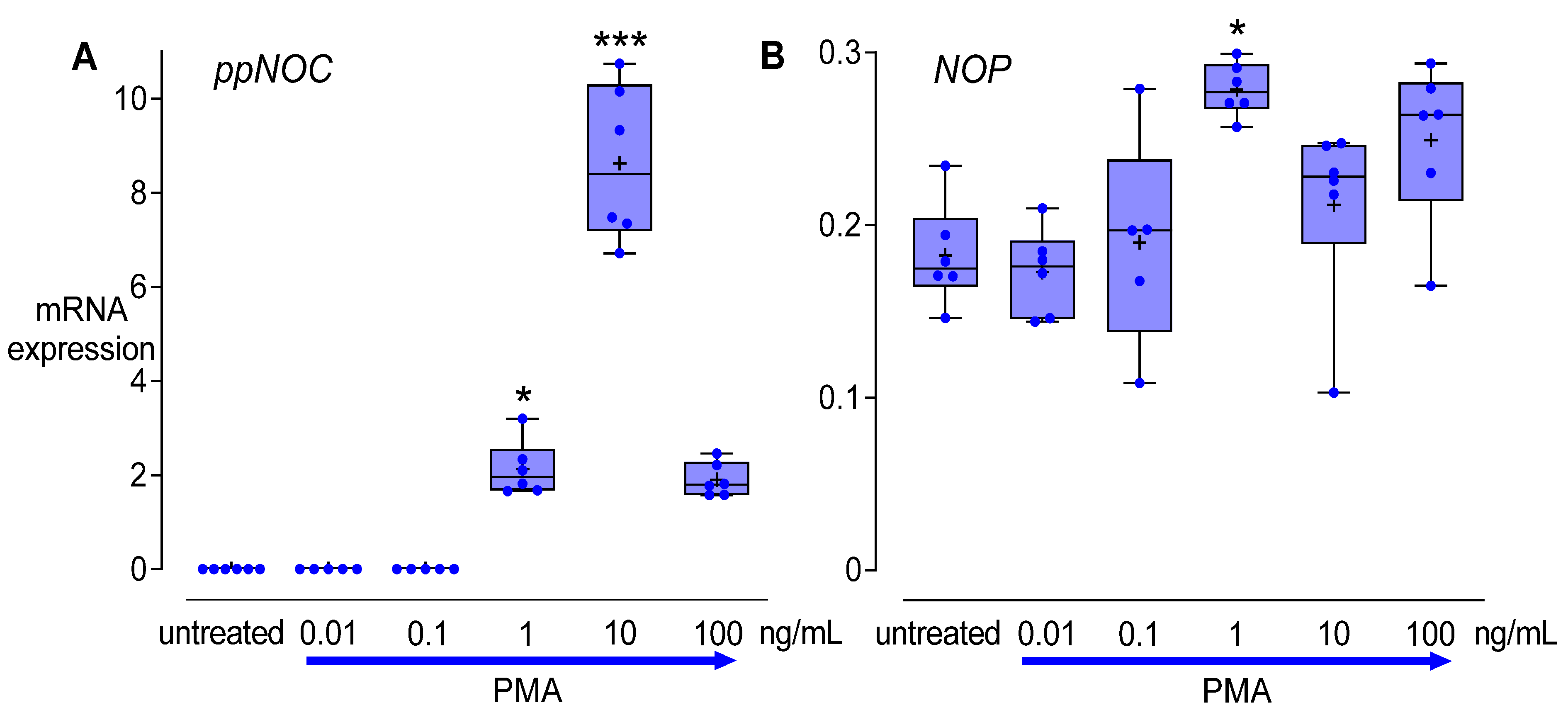
2.1.1. Cell Line Screening and Dose-Response Experiments
2.1.2. Cell Viability
2.1.3. Co-Stimulation of THP-1 with PMA and TLR Agonists
2.1.4. Stimulation of THP-1 with Nociceptin
2.2. RNA Isolation, cDNA Synthesis and Relative Quantification
2.3. Flow Cytometry
2.3.1. Measurement of Cell Membrane Proteins
2.3.2. Measurement of Intracellular Proteins
2.4. Statistical Analysis
3. Results
3.1. Nociceptin and NOP Expression in Different Cell Lines
3.2. Dose-Dependent Effects of PMA
3.3. Cell Viability
3.4. Nociceptin, NOP, and TLR Expression in THP-1
3.5. Effects of TLR Agonists on the Nociceptin System
3.6. Effects of Activation of the Nociceptin System on TLR Expression
4. Discussion
4.1. PMA-Induced THP-1 Model
4.2. Effects of TLR Signaling on the Nociceptin System
4.3. Nociceptin Effects on TLRs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lambert, D.G. The nociceptin/orphanin FQ receptor: A target with broad therapeutic potential. Nat. Rev. Drug Discov. 2008, 7, 694–710. [Google Scholar] [CrossRef] [PubMed]
- Calo, G.; Lambert, D.G. Nociceptin/orphanin FQ receptor ligands and translational challenges: Focus on cebranopadol as an innovative analgesic. Br. J. Anaesth. 2018, 121, 1105–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroder, W.; Lambert, D.G.; Ko, M.C.; Koch, T. Functional plasticity of the N/OFQ-NOP receptor system determines analgesic properties of NOP receptor agonists. Br. J. Pharmacol. 2014, 171, 3777–3800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bird, M.F.; McDonald, J.; Horley, B.; O’Doherty, J.P.; Fraser, B.; Gibson, C.L.; Guerrini, R.; Caló, G.; Lambert, D.G. MOP and NOP receptor interaction: Studies with a dual expression system and bivalent peptide ligands. PLoS ONE 2022, 17, 0260880. [Google Scholar] [CrossRef]
- Gavioli, E.C.; de Medeiros, I.U.; Monteiro, M.C.; Calo, G.; Romao, P.R. Nociceptin/orphanin FQ-NOP receptor system in inflammatory and immune-mediated diseases. Vitam. Horm. 2015, 97, 241–266. [Google Scholar]
- Serrano-Gomez, A.; Thompson, J.P.; Lambert, D.G. Nociceptin/orphanin FQ in inflammation and sepsis. Br. J. Anaesth. 2011, 106, 6–12. [Google Scholar] [CrossRef] [Green Version]
- Stamer, U.M.; Book, M.; Comos, C.; Zhang, L.; Nauck, F.; Stuber, F. Expression of the nociceptin precursor and nociceptin receptor is modulated in cancer and septic patients. Br. J. Anaesth. 2011, 106, 566–572. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Stuber, F.; Stamer, U.M. Inflammatory mediators influence the expression of nociceptin and its receptor in human whole blood cultures. PLoS ONE 2013, 8, e74138. [Google Scholar] [CrossRef] [Green Version]
- Al-Hashimi, M.; McDonald, J.; Thompson, J.P.; Lambert, D.G. Evidence for nociceptin/orphanin FQ (NOP) but not micro (MOP), delta (DOP) or kappa (KOP) opioid receptor mRNA in whole human blood. Br. J. Anaesth. 2016, 116, 423–429. [Google Scholar] [CrossRef] [Green Version]
- Vijay, K. Toll-like receptors in immunity and inflammatory diseases: Past, present, and future. Int. Immunopharmacol. 2018, 59, 391–412. [Google Scholar] [CrossRef]
- Lacagnina, M.J.; Watkins, L.R.; Grace, P.M. Toll-like receptors and their role in persistent pain. Pharmacol. Ther. 2018, 184, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Tabata, H.; Sasaki, M.; Kataoka-Sasaki, Y.; Shinkai, N.; Ichihara, K.; Masumori, N.; Kocsis, J.D.; Honmou, O. Possible role of intravenous administration of mesenchymal stem cells to alleviate interstitial cystitis/bladder pain syndrome in a Toll-like receptor-7 agonist-induced experimental animal model in rat. BMC Urol. 2021, 21, 156. [Google Scholar] [CrossRef] [PubMed]
- Kwok, Y.H.; Tuke, J.; Nicotra, L.L.; Grace, P.M.; Rolan, P.E.; Hutchinson, M.R. TLR 2 and 4 responsiveness from isolated peripheral blood mononuclear cells from rats and humans as potential chronic pain biomarkers. PLoS ONE 2013, 8, e77799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, M.; Choi, S. Toll-like Receptor-Dependent Negative Effects of Opioids: A Battle between Analgesia and Hyperalgesia. Front. Immunol. 2017, 8, 642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, C.W.; Aravind, S.; Das, S.; Davis, R.L. Pharmacological characterization of LPS and opioid interactions at the toll-like receptor 4. Br. J. Pharmacol. 2013, 168, 1421–1429. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Yang, M.; Chen, C.; Liu, L.; Wei, X.; Zeng, S. Toll-Like Receptor 4 (TLR4)/Opioid Receptor Pathway Crosstalk and Impact on Opioid Analgesia, Immune Function, and Gastrointestinal Motility. Front. Immunol. 2020, 11, 1455. [Google Scholar] [CrossRef]
- Sauer, R.S.; Hackel, D.; Morschel, L.; Sahlbach, H.; Wang, Y.; Mousa, S.A.; Roewer, N.; Brack, A.; Rittner, H.L. Toll like receptor (TLR)-4 as a regulator of peripheral endogenous opioid-mediated analgesia in inflammation. Mol. Pain 2014, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Stuber, F.; Lippuner, C.; Schiff, M.; Stamer, U.M. Phorbol-12-myristate-13-acetate induces nociceptin in human Mono Mac 6 cells via multiple transduction signaling pathways. Br. J. Anaesth. 2016, 117, 250–257. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Stuber, F.; Lippuner, C.; Schiff, M.; Stamer, U.M. ERK and p38 contribute to the regulation of nociceptin and the nociceptin receptor in human peripheral blood leukocytes. Mol. Pain 2019, 15, 1744806919828921. [Google Scholar] [CrossRef] [Green Version]
- Finney, S.J.; Leaver, S.K.; Evans, T.W.; Burke-Gaffney, A. Differences in lipopolysaccharide- and lipoteichoic acid-induced cytokine/chemokine expression. Intensive Care Med. 2012, 38, 324–332. [Google Scholar] [CrossRef] [Green Version]
- Boyette, L.B.; Macedo, C.; Hadi, K.; Elinoff, B.D.; Walters, J.T.; Ramaswami, B.; Chalasani, G.; Taboas, J.M.; Lakkis, F.G.; Metes, D.M. Phenotype, function, and differentiation potential of human monocyte subsets. PLoS ONE 2017, 12, e0176460. [Google Scholar] [CrossRef] [PubMed]
- Medepalli, K.; Alphenaar, B.W.; Keynton, R.S.; Sethu, P. A new technique for reversible permeabilization of live cells for intracellular delivery of quantum dots. Nanotechnology 2013, 24, 205101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.P.; Serrano-Gomez, A.; McDonald, J.; Ladak, N.; Bowrey, S.; Lambert, D.G. The Nociceptin/Orphanin FQ system is modulated in patients admitted to ICU with sepsis and after cardiopulmonary bypass. PLoS ONE 2013, 8, e76682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, J.P.; Thompson, J.P.; Young, S.P.; Gold, S.J.; McDonald, J.; Rowbotham, D.J.; Lambert, D.G. Nociceptin and urotensin-II concentrations in critically ill patients with sepsis. Br. J. Anaesth. 2008, 100, 810–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbeek, T.A.; Jarbadan, N.R.; Davis, C.; Caldwell, J. Nociceptin is present in synovial fluid of patients undergoing total knee arthroplasty. J. Orthop. Surg. Res. 2020, 15, 266. [Google Scholar] [CrossRef]
- Serhan, C.N.; Fierro, I.M.; Chiang, N.; Pouliot, M. Cutting edge: Nociceptin stimulates neutrophil chemotaxis and recruitment: Inhibition by aspirin-triggered-15-epi-lipoxin A4. J. Immunol. 2001, 166, 3650–3654. [Google Scholar] [CrossRef] [Green Version]
- Lambert, D.G.; Bird, M.F.; Rowbotham, D.J. Cebranopadol: A first in-class example of a nociceptin/orphanin FQ receptor and opioid receptor agonist. Br. J. Anaesth. 2015, 114, 364–366. [Google Scholar] [CrossRef] [Green Version]
- Christoph, A.; Eerdekens, M.H.; Kok, M.; Volkers, G.; Freynhagen, R. Cebranopadol, a novel first-in-class analgesic drug candidate: First experience in patients with chronic low back pain in a randomized clinical trial. Pain 2017, 158, 1813–1824. [Google Scholar] [CrossRef] [Green Version]
- Ubaldi, M.; Cannella, N.; Borruto, A.M.; Petrella, M.; Micioni Di Bonaventura, M.V.; Soverchia, L.; Stopponi, S.; Weiss, F.; Cifani, C.; Ciccocioppo, R. Role of Nociceptin/Orphanin FQ-NOP Receptor System in the Regulation of Stress-Related Disorders. Int J. Mol. Sci. 2021, 22, 12956. [Google Scholar] [CrossRef]
- Kiguchi, N.; Kishioka, S.; Ko, M.C. [Therapeutic potentials of safe opioid analgesics targeting nociceptin/orphanin FQ peptide receptor]. Nihon Yakurigaku Zasshi 2021, 156, 139–144. [Google Scholar] [CrossRef]
- Kiguchi, N.; Ding, H.; Kishioka, S.; Ko, M.C. Nociceptin/Orphanin FQ Peptide Receptor-Related Ligands as Novel Analgesics. Curr. Top. Med. Chem. 2020, 20, 2878–2888. [Google Scholar] [CrossRef] [PubMed]
- Theus, S.A.; Cave, M.D.; Eisenach, K.D. Activated THP-1 cells: An attractive model for the assessment of intracellular growth rates of Mycobacterium tuberculosis isolates. Infect. Immun. 2004, 72, 1169–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurga, A.M.; Rojewska, E.; Piotrowska, A.; Makuch, W.; Pilat, D.; Przewlocka, B.; Mika, J. Blockade of Toll-Like Receptors (TLR2, TLR4) Attenuates Pain and Potentiates Buprenorphine Analgesia in a Rat Neuropathic Pain Model. Neural Plast. 2016, 2016, 5238730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araldi, D.; Bogen, O.; Green, P.G.; Levine, J.D. Role of Nociceptor Toll-like Receptor 4 (TLR4) in Opioid-Induced Hyperalgesia and Hyperalgesic Priming. J. Neurosci. 2019, 39, 6414–6424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franchi, S.; Moretti, S.; Castelli, M.; Lattuada, D.; Scavullo, C.; Panerai, A.E.; Sacerdote, P. Mu opioid receptor activation modulates Toll like receptor 4 in murine macrophages. Brain Behav. Immun. 2012, 26, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Li, Q. Antagonists of toll like receptor 4 maybe a new strategy to counteract opioid-induced hyperalgesia and opioid tolerance. Med. Hypotheses 2012, 79, 754–756. [Google Scholar] [CrossRef] [PubMed]
- Fiset, M.E.; Gilbert, C.; Poubelle, P.E.; Pouliot, M. Human neutrophils as a source of nociceptin: A novel link between pain and inflammation. Biochemistry 2003, 42, 10498–10505. [Google Scholar] [CrossRef] [Green Version]
- Lutfy, K.; Cowan, A. Buprenorphine: A unique drug with complex pharmacology. Curr. Neuropharmacol. 2004, 2, 395–402. [Google Scholar] [CrossRef]
- Peckham, A.M.; De La Cruz, A.; Dufresne, R.L. Kappa opioid receptor antagonism: Are opioids the answer for treatment resistant depression? Ment. Health Clin. 2018, 8, 175–183. [Google Scholar] [CrossRef]
- Azzam, A.A.H.; McDonald, J.; Lambert, D.G. Hot topics in opioid pharmacology: Mixed and biased opioids. Br. J. Anaesth. 2019, 122, e136–e145. [Google Scholar] [CrossRef]
- Gudin, J.; Fudin, J. A Narrative Pharmacological Review of Buprenorphine: A Unique Opioid for the Treatment of Chronic Pain. Pain Ther. 2020, 9, 41–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Infantino, R.; Mattia, C.; Locarini, P.; Pastore, A.L.; Maione, S.; Luongo, L. Buprenorphine: Far Beyond the “Ceiling”. Biomolecules 2021, 11, 816. [Google Scholar] [CrossRef] [PubMed]
- Bird, M.F.; McDonald, J.; Gallacher-Horley, B.; Guerrini, R.; Calo, G.; Ye, S.; Thompson, J.P.; Lambert, D.G. Functional nociceptin receptors are upregulated on human umbilical vein endothelial cells during experimental in vitro sepsis. Br. J. Anaesth. 2020, 124, E205–E206. [Google Scholar] [CrossRef]
- Williams, J.P.; Thompson, J.P.; McDonald, J.; Barnes, T.A.; Cote, T.; Rowbotham, D.J.; Lambert, D.G. Human peripheral blood mononuclear cells express nociceptin/orphanin FQ, but not mu, delta, or kappa opioid receptors. Anesth. Analg. 2007, 105, 998–1005. [Google Scholar] [CrossRef]
- Williams, J.P.; Thompson, J.P.; Rowbotham, D.J.; Lambert, D.G. Human peripheral blood mononuclear cells produce pre-pro-nociceptin/orphanin FQ mRNA. Anesth. Analg. 2008, 106, 865–866. [Google Scholar] [CrossRef]
- Petrella, C.; Giuli, C.; Broccardo, M.; Eutamene, H.; Cartier, C.; Leveque, M.; Bedini, A.; Spampinato, S.; Bueno, L.; Theodorou, V.; et al. Protective and worsening peripheral nociceptin/orphanin FQ receptor-mediated effect in a rat model of experimental colitis. Pharmacol. Res. 2013, 70, 72–79. [Google Scholar] [CrossRef]
- Sobczak, M.; Mokrowiecka, A.; Cygankiewicz, A.I.; Zakrzewski, P.K.; Salaga, M.; Storr, M.; Kordek, R.; Malecka-Panas, E.; Krajewska, W.M.; Fichna, J. Anti-inflammatory and antinociceptive action of an orally available nociceptin receptor agonist SCH 221510 in a mouse model of inflammatory bowel diseases. J. Pharmacol. Exp. Ther. 2014, 348, 401–409. [Google Scholar] [CrossRef] [Green Version]
- Alt, C.; Lam, J.S.; Harrison, M.T.; Kershaw, K.M.; Samuelsson, S.; Toll, L.; D’Andrea, A. Nociceptin/orphanin FQ inhibition with SB612111 ameliorates dextran sodium sulfate-induced colitis. Eur. J. Pharmacol. 2012, 683, 285–293. [Google Scholar] [CrossRef]
- Carvalho, D.; Petronilho, F.; Vuolo, F.; Machado, R.A.; Constantino, L.; Guerrini, R.; Calo, G.; Gavioli, E.C.; Streck, E.L.; Dal-Pizzol, F. The nociceptin/orphanin FQ-NOP receptor antagonist effects on an animal model of sepsis. Intensive Care Med. 2008, 34, 2284–2290. [Google Scholar] [CrossRef]
- Harter, L.; Mica, L.; Stocker, R.; Trentz, O.; Keel, M. Increased expression of toll-like receptor-2 and -4 on leukocytes from patients with sepsis. Shock 2004, 22, 403–409. [Google Scholar] [CrossRef]
- Salomao, R.; Martins, P.S.; Brunialti, M.K.; Fernandes Mda, L.; Martos, L.S.; Mendes, M.E.; Gomes, N.E.; Rigato, O. TLR signaling pathway in patients with sepsis. Shock 2008, 30 (Suppl. 1), 73–77. [Google Scholar] [CrossRef] [PubMed]
- Kwok, Y.H.; Hutchinson, M.R.; Gentgall, M.G.; Rolan, P.E. Increased responsiveness of peripheral blood mononuclear cells to in vitro TLR 2, 4 and 7 ligand stimulation in chronic pain patients. PLoS ONE 2012, 7, e44232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaaf, B.; Luitjens, K.; Goldmann, T.; van Bremen, T.; Sayk, F.; Dodt, C.; Dalhoff, K.; Droemann, D. Mortality in human sepsis is associated with downregulation of Toll-like receptor 2 and CD14 expression on blood monocytes. Diagn. Pathol. 2009, 4, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chanput, W.; Mes, J.J.; Wichers, H.J. THP-1 cell line: An in vitro cell model for immune modulation approach. Int. Iimmunopharmacol. 2014, 23, 37–45. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence 5′ to 3′ | RT-qPCR Reaction Conditions | |
|---|---|---|---|
| HPRT1 | Fwd | TGACCTTGATTTATTTTGCATACC | |
| Rev | CGAGCAAGACGTTCAGTCCT | ||
| GAPDH | Fwd | AGCCACATCGCTCAGACAC | |
| Rev | GCCCAATACGACCAAATCC | ||
| ppNOC | Fwd | GGACAGCTTCGACCTGGAG | pre-incubation |
| Rev | TGACCTTGGTGCATGGAGT | 95 °C, 10 min | |
| NOP | Fwd | CCCAAGGAGGTTGCAGAA | amplification (45 cycles) |
| Rev | GCCGTAGATAACCTCCCAGA | 95 °C, 10 s | |
| TLR2 | Fwd | CTCTCGGTGTCGGAATGTC | 60 °C, 30 s |
| Rev | AGGATCAGCAGGAACAGAGC | 72 °C, 1 s | |
| TLR4 | Fwd | CAAGATGCCCCTTCCATTT | |
| Rev | TCCTTAGGAATTAGCCACTAGACTTT | ||
| TLR7 | Fwd | GCCCCCAAGATGGTTTAAG | |
| Rev | GCATCCCCAATTTCTTTGG | ||
| TLR9 | Fwd | CGCTACTGGTGCTATCCAGA | |
| Rev | AGCCCAGGGAGGAGCTAAG |
| Stimulation | ppNOC | NOP |
|---|---|---|
| untreated | 0 | 0.08 (0.07/0.10) |
| LTA 10 µg/mL | 0 | 0.05 (0.04/0.08) |
| LPS 100 ng/mL | 0 | 0.04 (0.04/0.06) |
| IMQ 10 µg/mL | 0 | 0.09 (0.07/0.11) |
| ODN 2216 1 µM | 0 | 0.05 (0.03/0.07) |
| PMA 5 ng/mL | 8.90 (8.20/9.73) | 0.04 (0.03/0.06) |
| PMA+LTA | 0.03 (0.01/0.03) ** | 0.04 (0.04/0.05) |
| PMA+LPS | 0.04 (0.01/0.07) ** | 0.04 (0.03/0.06) |
| PMA+IMQ | 2.21 (1.77/2.41) ** | 0.05 (0.04/0.08) |
| PMA+ODN 2216 | 7.51 (6.43/7.90) * | 0.06 (0.02/0.11) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Stamer, U.M.; Huang, M.Y.-Y.; Stüber, F. Interactions between the Nociceptin and Toll-like Receptor Systems. Cells 2022, 11, 1085. https://doi.org/10.3390/cells11071085
Zhang L, Stamer UM, Huang MY-Y, Stüber F. Interactions between the Nociceptin and Toll-like Receptor Systems. Cells. 2022; 11(7):1085. https://doi.org/10.3390/cells11071085
Chicago/Turabian StyleZhang, Lan, Ulrike M. Stamer, Melody Ying-Yu Huang, and Frank Stüber. 2022. "Interactions between the Nociceptin and Toll-like Receptor Systems" Cells 11, no. 7: 1085. https://doi.org/10.3390/cells11071085