
Hsp70 in Redox Homeostasis



Abstract
:1. Introduction
2. Hsp70 System
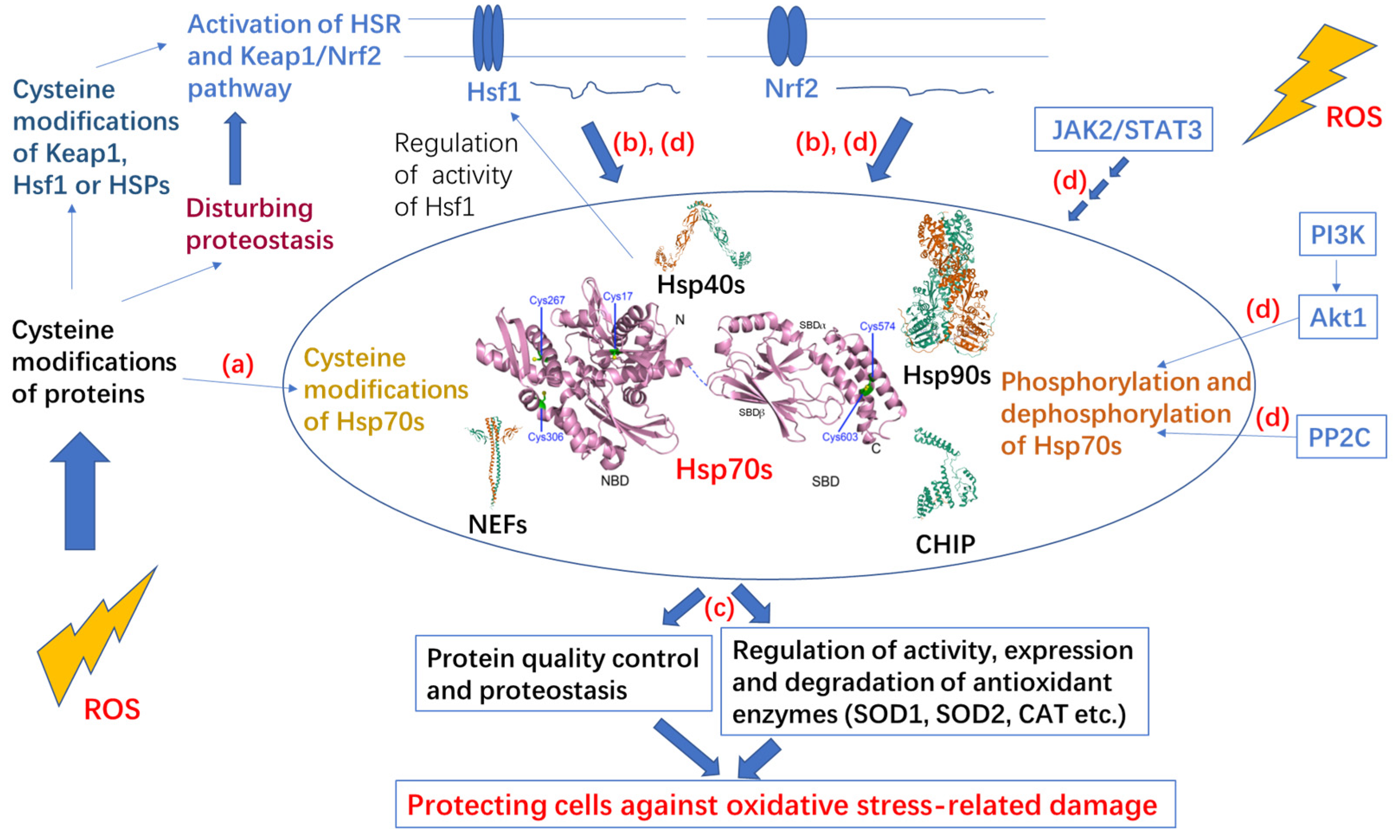
3. Post-Translational Modifications of Hsp70 under Oxidative Stress
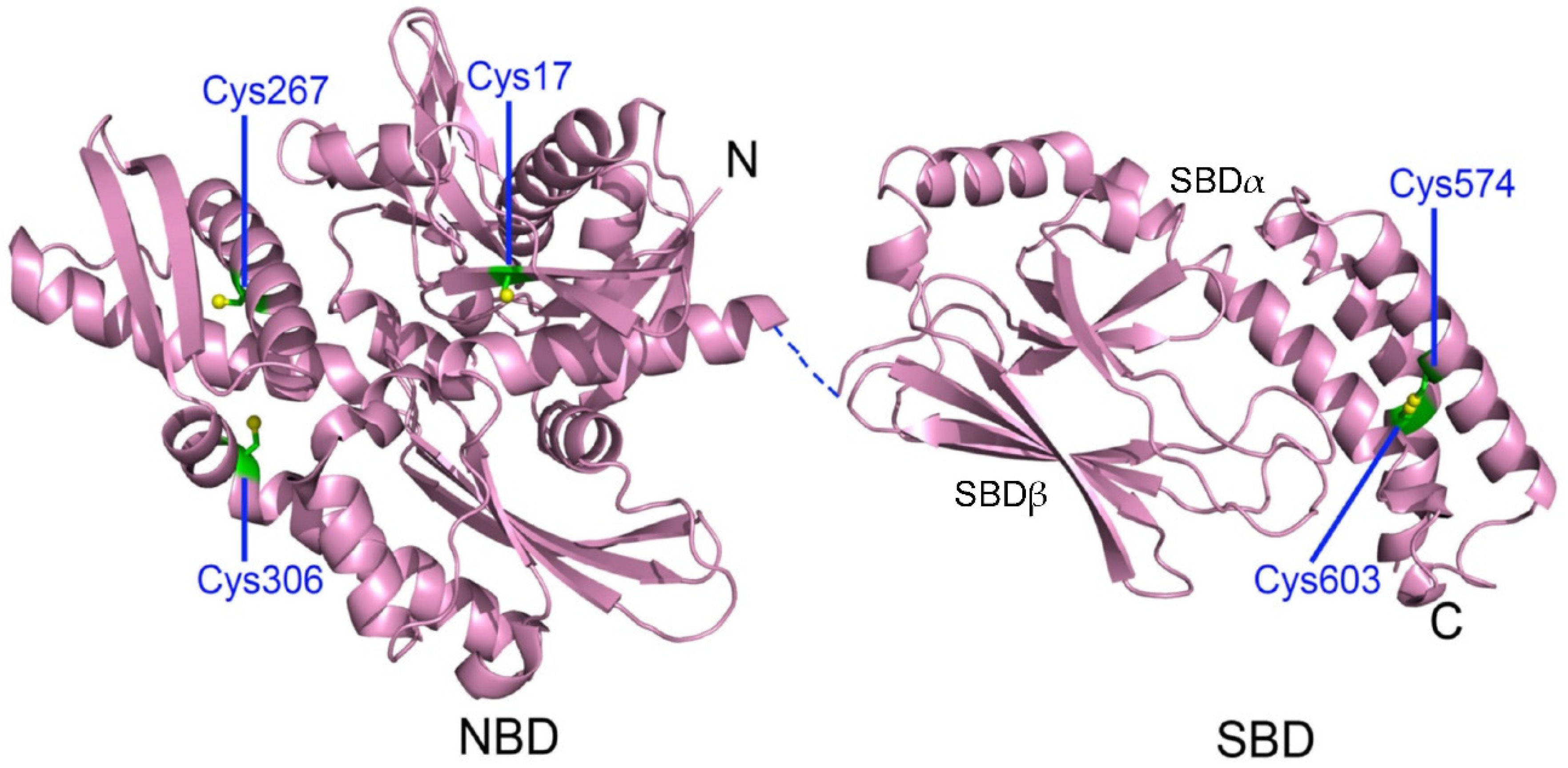
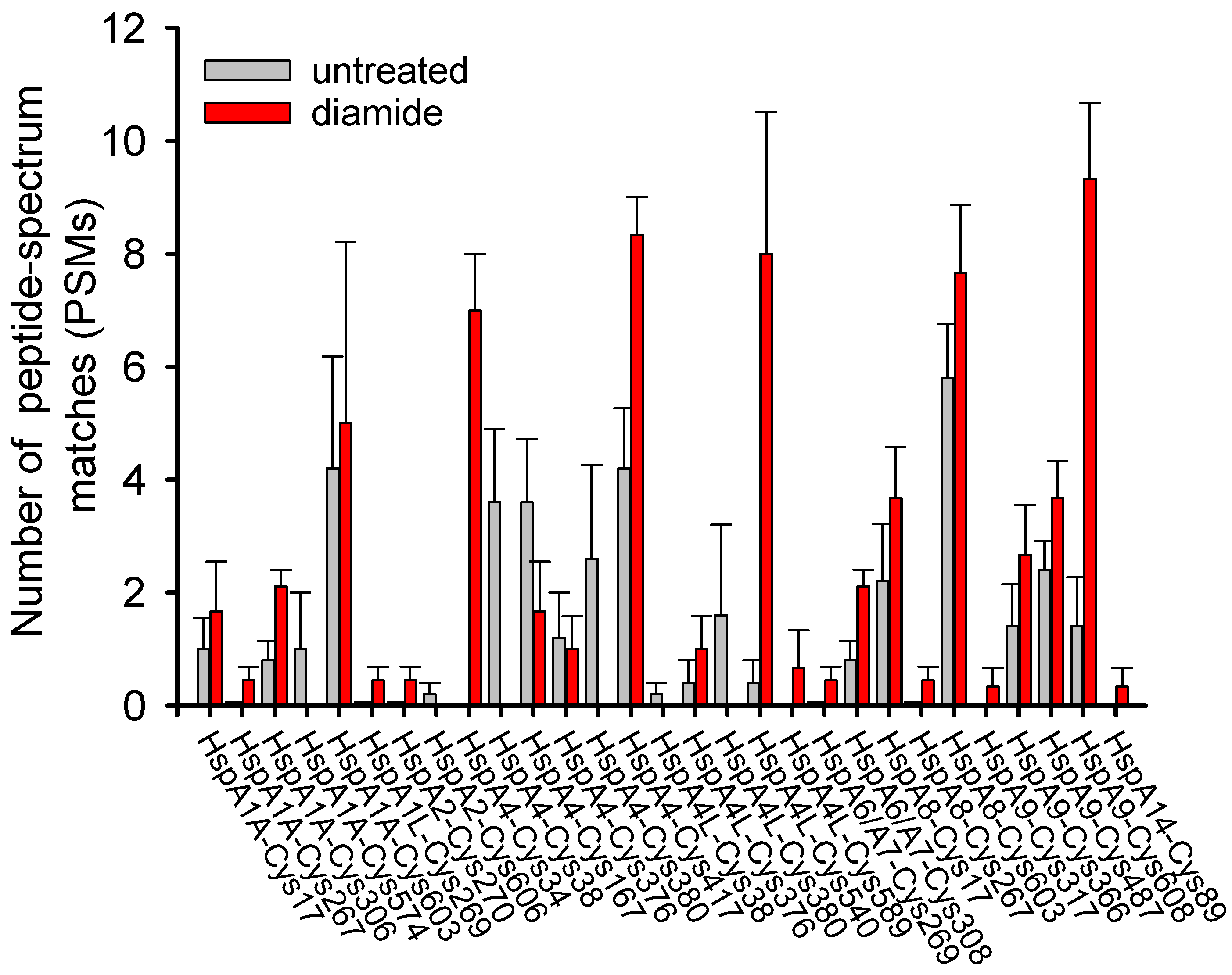
3.1. Cysteine Oxidation of Hsp70 and Redox Homeostasis
3.2. Covalent Modifications of Hsp70 and Redox Homeostasis
4. Protection Effect and Upregulated Expression of Hsp70 under Oxidative Stress
5. Hsp70 Participates in Multiple Redox-Related Signaling Pathways
6. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Holmström, K.M.; Finkel, T. Cellular mechanisms and physiological consequences of redox-dependent signalling. Nat. Rev. Mol. Cell Biol. 2014, 15, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.; Chandel, N.S. Regulation of redox balance in cancer and T cells. J. Biol. Chem. 2018, 293, 7499–7507. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, X.; Cueto, R.; Effi, C.; Zhang, Y.; Tan, H.; Qin, X.; Ji, Y.; Yang, X.; Wang, H. Biochemical basis and metabolic interplay of redox regulation. Redox Biol. 2019, 26, 101284. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, P.D.; Huang, B.W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef] [Green Version]
- Tabak, O.; Gelisgen, R.; Erman, H.; Erdenen, F.; Muderrisoglu, C.; Aral, H.; Uzun, H. Oxidative lipid, protein, and DNA damage as oxidative stress markers in vascular complications of diabetes mellitus. Clin. Investig. Med. 2011, 34, E163–E171. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Wang, X.; Shigenaga, M.K.; Yeo, H.C.; Mori, A.; Ames, B.N. Immobilization stress causes oxidative damage to lipid, protein, and DNA in the brain of rats. Faseb J. 1996, 10, 1532–1538. [Google Scholar] [CrossRef]
- Yang, W.S.; Stockwell, B.R. Ferroptosis: Death by Lipid Peroxidation. Trends. Cell Biol. 2016, 26, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Forman, H.J.; Ursini, F.; Maiorino, M. An overview of mechanisms of redox signaling. J. Mol. Cell Cardiol. 2014, 73, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Forman, H.J. Redox signaling: An evolution from free radicals to aging. Free Radic. Biol. Med. 2016, 97, 398–407. [Google Scholar] [CrossRef] [Green Version]
- Turell, L.; Zeida, A.; Trujillo, M. Mechanisms and consequences of protein cysteine oxidation: The role of the initial short-lived intermediates. Essays Biochem. 2020, 64, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Carroll, K.S.; Liebler, D.C. The Expanding Landscape of the Thiol Redox Proteome. Mol. Cell Proteom. 2016, 15, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkel, T. Signal transduction by reactive oxygen species. J. Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szyller, J.; Bil-Lula, I. Heat Shock Proteins in Oxidative Stress and Ischemia/Reperfusion Injury and Benefits from Physical Exercises: A Review to the Current Knowledge. Oxid. Med. Cell Longev. 2021, 2021, 6678457. [Google Scholar] [CrossRef]
- Kalmar, B.; Greensmith, L. Induction of heat shock proteins for protection against oxidative stress. Adv. Drug Deliv. Rev. 2009, 61, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef] [Green Version]
- Korovila, I.; Hugo, M.; Castro, J.P.; Weber, D.; Höhn, A.; Grune, T.; Jung, T. Proteostasis, oxidative stress and aging. Redox Biol. 2017, 13, 550–567. [Google Scholar] [CrossRef]
- Fernández-Fernández, M.R.; Valpuesta, J.M. Hsp70 chaperone: A master player in protein homeostasis. F1000Research 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Large, A.T.; Goldberg, M.D.; Lund, P.A. Chaperones and protein folding in the archaea. Biochem. Soc. Trans. 2009, 37, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Rebeaud, M.E.; Mallik, S.; Goloubinoff, P.; Tawfik, D.S. On the evolution of chaperones and cochaperones and the expansion of proteomes across the Tree of Life. Proc. Natl. Acad. Sci. USA 2021, 118, e2020885118. [Google Scholar] [CrossRef]
- Rosenzweig, R.; Nillegoda, N.B.; Mayer, M.P.; Bukau, B. The Hsp70 chaperone network. Nat. Rev. Mol. Cell Biol. 2019, 20, 665–680. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.E. The HSP70 family and cancer. Carcinogenesis 2013, 34, 1181–1188. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, L.; Young, J.C. Function and Chemotypes of Human Hsp70 Chaperones. Curr. Top. Med. Chem. 2016, 16, 2812–2828. [Google Scholar] [CrossRef] [PubMed]
- Kabani, M.; Martineau, C.N. Multiple hsp70 isoforms in the eukaryotic cytosol: Mere redundancy or functional specificity? Curr. Genom. 2008, 9, 338–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radons, J. The human HSP70 family of chaperones: Where do we stand? Cell Stress Chaperones 2016, 21, 379–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertelsen, E.B.; Chang, L.; Gestwicki, J.E.; Zuiderweg, E.R. Solution conformation of wild-type E. coli Hsp70 (DnaK) chaperone complexed with ADP and substrate. Proc. Natl. Acad. Sci. USA 2009, 106, 8471–8476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuravleva, A.; Gierasch, L.M. Allosteric signal transmission in the nucleotide-binding domain of 70-kDa heat shock protein (Hsp70) molecular chaperones. Proc. Natl. Acad. Sci. USA 2011, 108, 6987–6992. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zuiderweg, E.R. The 70-kDa heat shock protein chaperone nucleotide-binding domain in solution unveiled as a molecular machine that can reorient its functional subdomains. Proc. Natl. Acad. Sci. USA 2004, 101, 10272–10277. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Leu, J.I.; Murphy, M.E.; George, D.L.; Marmorstein, R. Crystal structure of the stress-inducible human heat shock protein 70 substrate-binding domain in complex with peptide substrate. PLoS ONE 2014, 9, e103518. [Google Scholar] [CrossRef] [Green Version]
- Zhuravleva, A.; Gierasch, L.M. Substrate-binding domain conformational dynamics mediate Hsp70 allostery. Proc. Natl. Acad. Sci. USA 2015, 112, E2865–E2873. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Gong, W.; Cusack, S.A.; Wu, H.; Loovers, H.M.; Zhang, H.; Perrett, S.; Jones, G.W. The β6/β7 region of the Hsp70 substrate-binding domain mediates heat-shock response and prion propagation. Cell Mol. Life Sci. 2018, 75, 1445–1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Zhang, H.; Cuskelly, D.D.; Doyle, S.; Perrett, S.; Jones, G.W. Mutational analysis of the Hsp70 substrate-binding domain: Correlating molecular-level changes with in vivo function. Mol. Microbiol. 2021, 115, 1262–1276. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Rossi, P.; Kalodimos, C.G. Structural basis for client recognition and activity of Hsp40 chaperones. Science 2019, 365, 1313–1319. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y.; Sakai, J.; Skach, W.R. Endoplasmic reticulum protein quality control is determined by cooperative interactions between Hsp/c70 protein and the CHIP E3 ligase. J. Biol. Chem. 2013, 288, 31069–31079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smock, R.G.; Blackburn, M.E.; Gierasch, L.M. Conserved, disordered C terminus of DnaK enhances cellular survival upon stress and DnaK in vitro chaperone activity. J. Biol. Chem. 2011, 286, 31821–31829. [Google Scholar] [CrossRef] [Green Version]
- Gong, W.; Hu, W.; Xu, L.; Wu, H.; Wu, S.; Zhang, H.; Wang, J.; Jones, G.W.; Perrett, S. The C-terminal GGAP motif of Hsp70 mediates substrate recognition and stress response in yeast. J. Biol. Chem. 2018, 293, 17663–17675. [Google Scholar] [CrossRef] [Green Version]
- Zuiderweg, E.R.; Bertelsen, E.B.; Rousaki, A.; Mayer, M.P.; Gestwicki, J.E.; Ahmad, A. Allostery in the Hsp70 chaperone proteins. Top Curr. Chem. 2013, 328, 99–153. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Hong, L.; Wang, Y.; Yu, J.; Yang, J.; Yang, J.; Zhang, H.; Perrett, S. Kinetics of the conformational cycle of Hsp70 reveals the importance of the dynamic and heterogeneous nature of Hsp70 for its function. Proc. Natl. Acad. Sci. USA 2020, 117, 7814–7823. [Google Scholar] [CrossRef] [Green Version]
- Kityk, R.; Kopp, J.; Sinning, I.; Mayer, M.P. Structure and dynamics of the ATP-bound open conformation of Hsp70 chaperones. Mol. Cell 2012, 48, 863–874. [Google Scholar] [CrossRef] [Green Version]
- Qi, R.; Sarbeng, E.B.; Liu, Q.; Le, K.Q.; Xu, X.; Xu, H.; Yang, J.; Wong, J.L.; Vorvis, C.; Hendrickson, W.A.; et al. Allosteric opening of the polypeptide-binding site when an Hsp70 binds ATP. Nat. Struct. Mol. Biol. 2013, 20, 900–907. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Liu, Q.; Liu, Q.; Hendrickson, W.A. Conformational equilibria in allosteric control of Hsp70 chaperones. Mol. Cell 2021, 81, 3919–3933. [Google Scholar] [CrossRef] [PubMed]
- Meng, W.; Clerico, E.M.; McArthur, N.; Gierasch, L.M. Allosteric landscapes of eukaryotic cytoplasmic Hsp70s are shaped by evolutionary tuning of key interfaces. Proc. Natl. Acad. Sci. USA 2018, 115, 11970–11975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuravleva, A.; Clerico, E.M.; Gierasch, L.M. An interdomain energetic tug-of-war creates the allosterically active state in Hsp70 molecular chaperones. Cell 2012, 151, 1296–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faust, O.; Abayev-Avraham, M.; Wentink, A.S.; Maurer, M.; Nillegoda, N.B.; London, N.; Bukau, B.; Rosenzweig, R. HSP40 proteins use class-specific regulation to drive HSP70 functional diversity. Nature 2020, 587, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Polier, S.; Dragovic, Z.; Hartl, F.U.; Bracher, A. Structural basis for the cooperation of Hsp70 and Hsp110 chaperones in protein folding. Cell 2008, 133, 1068–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bracher, A.; Verghese, J. The nucleotide exchange factors of Hsp70 molecular chaperones. Front. Mol. Biosci. 2015, 2, 10. [Google Scholar] [CrossRef]
- Qiu, X.B.; Shao, Y.M.; Miao, S.; Wang, L. The diversity of the DnaJ/Hsp40 family, the crucial partners for Hsp70 chaperones. Cell Mol. Life Sci. 2006, 63, 2560–2570. [Google Scholar] [CrossRef]
- Kravats, A.N.; Hoskins, J.R.; Reidy, M.; Johnson, J.L.; Doyle, S.M.; Genest, O.; Masison, D.C.; Wickner, S. Functional and physical interaction between yeast Hsp90 and Hsp70. Proc. Natl. Acad. Sci. USA 2018, 115, E2210–E2219. [Google Scholar] [CrossRef] [Green Version]
- Winkler, J.; Tyedmers, J.; Bukau, B.; Mogk, A. Hsp70 targets Hsp100 chaperones to substrates for protein disaggregation and prion fragmentation. J. Cell Biol. 2012, 198, 387–404. [Google Scholar] [CrossRef] [Green Version]
- Stankiewicz, M.; Nikolay, R.; Rybin, V.; Mayer, M.P. CHIP participates in protein triage decisions by preferentially ubiquitinating Hsp70-bound substrates. Febs J. 2010, 277, 3353–3367. [Google Scholar] [CrossRef]
- Dice, J.F. Chaperone-mediated autophagy. Autophagy 2007, 3, 295–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Zhang, H.; Gong, W.; Liu, Z.; Wu, H.; Hu, W.; Chen, X.; Wang, L.; Wu, S.; Chen, C.; et al. S-Glutathionylation of human inducible Hsp70 reveals a regulatory mechanism involving the C-terminal α-helical lid. J. Biol. Chem. 2020, 295, 8302–8324. [Google Scholar] [CrossRef] [Green Version]
- Grunwald, M.S.; Pires, A.S.; Zanotto-Filho, A.; Gasparotto, J.; Gelain, D.P.; Demartini, D.R.; Schöler, C.M.; de Bittencourt, P.I., Jr.; Moreira, J.C. The oxidation of HSP70 is associated with functional impairment and lack of stimulatory capacity. Cell Stress Chaperones 2014, 19, 913–925. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gibney, P.A.; West, J.D.; Morano, K.A. The yeast Hsp70 Ssa1 is a sensor for activation of the heat shock response by thiol-reactive compounds. Mol. Biol. Cell 2012, 23, 3290–3298. [Google Scholar] [CrossRef] [PubMed]
- Weerapana, E.; Wang, C.; Simon, G.M.; Richter, F.; Khare, S.; Dillon, M.B.; Bachovchin, D.A.; Mowen, K.; Baker, D.; Cravatt, B.F. Quantitative reactivity profiling predicts functional cysteines in proteomes. Nature 2010, 468, 790–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.A.; Wang, Y.; Cheng, H.; Zhang, Q.; Ge, W.; Guo, D. RedoxDB--a curated database for experimentally verified protein oxidative modification. Bioinformatics 2012, 28, 2551–2552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Huang, B.; Zhang, L.; Zhang, Y.; Zhao, Y.; Guo, X.; Qiao, X.; Chen, C. SNObase, a database for S-nitrosation modification. Protein Cell 2012, 3, 929–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.J.; Lu, C.T.; Lee, T.Y.; Chen, Y.J. dbGSH: A database of S-glutathionylation. Bioinformatics 2014, 30, 2386–2388. [Google Scholar] [CrossRef]
- Chen, Y.J.; Lu, C.T.; Su, M.G.; Huang, K.Y.; Ching, W.C.; Yang, H.H.; Liao, Y.C.; Chen, Y.J.; Lee, T.Y. dbSNO 2.0: A resource for exploring structural environment, functional and disease association and regulatory network of protein S-nitrosylation. Nucleic Acids Res. 2015, 43, D503–D511. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Zhang, Q.; Li, S.; Cheng, B.; Xue, H.; Wei, Z.; Shao, T.; Liu, Z.X.; Cheng, H.; Wang, Z. iCysMod: An integrative database for protein cysteine modifications in eukaryotes. Brief Bioinform. 2021, 22, bbaa400. [Google Scholar] [CrossRef]
- Fratelli, M.; Gianazza, E.; Ghezzi, P. Redox proteomics: Identification and functional role of glutathionylated proteins. Expert Rev. Proteomics 2004, 1, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yang, J.; Wu, S.; Gong, W.; Chen, C.; Perrett, S. Glutathionylation of the Bacterial Hsp70 Chaperone DnaK Provides a Link between Oxidative Stress and the Heat Shock Response. J. Biol. Chem. 2016, 291, 6967–6981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamer, J.; Multhaup, G.; Tomoyasu, T.; McCarty, J.S.; Rüdiger, S.; Schönfeld, H.J.; Schirra, C.; Bujard, H.; Bukau, B. A cycle of binding and release of the DnaK, DnaJ and GrpE chaperones regulates activity of the Escherichia coli heat shock transcription factor sigma32. Embo J. 1996, 15, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Müller, A.; Hoffmann, J.H.; Meyer, H.E.; Narberhaus, F.; Jakob, U.; Leichert, L.I. Nonnative disulfide bond formation activates the σ32-dependent heat shock response in Escherichia coli. J. Bacteriol. 2013, 195, 2807–2816. [Google Scholar] [CrossRef] [Green Version]
- Winter, J.; Linke, K.; Jatzek, A.; Jakob, U. Severe oxidative stress causes inactivation of DnaK and activation of the redox-regulated chaperone Hsp33. Mol. Cell 2005, 17, 381–392. [Google Scholar] [CrossRef]
- Chung, H.S.; Murray, C.I.; Venkatraman, V.; Crowgey, E.L.; Rainer, P.P.; Cole, R.N.; Bomgarden, R.D.; Rogers, J.C.; Balkan, W.; Hare, J.M.; et al. Dual Labeling Biotin Switch Assay to Reduce Bias Derived From Different Cysteine Subpopulations: A Method to Maximize S-Nitrosylation Detection. Circ. Res. 2015, 117, 846–857. [Google Scholar] [CrossRef] [Green Version]
- Mnatsakanyan, R.; Markoutsa, S.; Walbrunn, K.; Roos, A.; Verhelst, S.H.L.; Zahedi, R.P. Proteome-wide detection of S-nitrosylation targets and motifs using bioorthogonal cleavable-linker-based enrichment and switch technique. Nat. Commun. 2019, 10, 2195. [Google Scholar] [CrossRef]
- Lefievre, L.; Chen, Y.; Conner, S.J.; Scott, J.L.; Publicover, S.J.; Ford, W.C.; Barratt, C.L. Human spermatozoa contain multiple targets for protein S-nitrosylation: An alternative mechanism of the modulation of sperm function by nitric oxide? Proteomics 2007, 7, 3066–3084. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Chen, S.C.; Wang, D.L. Shear flow increases S-nitrosylation of proteins in endothelial cells. Cardiovasc. Res. 2009, 83, 536–546. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Liu, K.; He, J.; Tian, C.; Yu, X.; Yang, J. Direct Proteomic Mapping of Cysteine Persulfidation. Antioxid. Redox Signal 2020, 33, 1061–1076. [Google Scholar] [CrossRef]
- Wu, Q.; Zhao, B.; Weng, Y.; Shan, Y.; Li, X.; Hu, Y.; Liang, Z.; Yuan, H.; Zhang, L.; Zhang, Y. Site-Specific Quantification of Persulfidome by Combining an Isotope-Coded Affinity Tag with Strong Cation-Exchange-Based Fractionation. Anal. Chem. 2019, 91, 14860–14864. [Google Scholar] [CrossRef] [PubMed]
- Miyata, Y.; Rauch, J.N.; Jinwal, U.K.; Thompson, A.D.; Srinivasan, S.; Dickey, C.A.; Gestwicki, J.E. Cysteine reactivity distinguishes redox sensing by the heat-inducible and constitutive forms of heat shock protein 70. Chem. Biol. 2012, 19, 1391–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, L.; Liu, K.; Ferreira, R.B.; Carroll, K.S.; Yang, J. Proteome-Wide Analysis of Cysteine S-Sulfenylation Using a Benzothiazine-Based Probe. Curr. Protoc. Protein Sci. 2019, 95, e76. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Gupta, V.; Carroll, K.S.; Liebler, D.C. Site-specific mapping and quantification of protein S-sulphenylation in cells. Nat. Commun. 2014, 5, 4776. [Google Scholar] [CrossRef] [Green Version]
- Akter, S.; Fu, L.; Jung, Y.; Conte, M.L.; Lawson, J.R.; Lowther, W.T.; Sun, R.; Liu, K.; Yang, J.; Carroll, K.S. Chemical proteomics reveals new targets of cysteine sulfinic acid reductase. Nat. Chem. Biol. 2018, 14, 995–1004. [Google Scholar] [CrossRef]
- Valek, L.; Heidler, J.; Scheving, R.; Wittig, I.; Tegeder, I. Nitric oxide contributes to protein homeostasis by S-nitrosylations of the chaperone HSPA8 and the ubiquitin ligase UBE2D. Redox Biol. 2019, 20, 217–235. [Google Scholar] [CrossRef]
- Wang, J.; Sevier, C.S. Formation and Reversibility of BiP Protein Cysteine Oxidation Facilitate Cell Survival during and post Oxidative Stress. J. Biol. Chem. 2016, 291, 7541–7557. [Google Scholar] [CrossRef] [Green Version]
- Flohé, L. The fairytale of the GSSG/GSH redox potential. Biochim. Biophys. Acta 2013, 1830, 3139–3142. [Google Scholar] [CrossRef]
- Wang, J.; Pareja, K.A.; Kaiser, C.A.; Sevier, C.S. Redox signaling via the molecular chaperone BiP protects cells against endoplasmic reticulum-derived oxidative stress. Elife 2014, 3, e03496. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Marsh, H.M.; Sevier, C.S. A Conserved Cysteine within the ATPase Domain of the Endoplasmic Reticulum Chaperone BiP is Necessary for a Complete Complement of BiP Activities. J. Mol. Biol. 2016, 428, 4168–4184. [Google Scholar] [CrossRef] [Green Version]
- Breitzig, M.; Bhimineni, C.; Lockey, R.; Kolliputi, N. 4-Hydroxy-2-nonenal: A critical target in oxidative stress? Am. J. Physiol. Cell Physiol. 2016, 311, C537–C543. [Google Scholar] [CrossRef]
- Chaudhary, P.; Sharma, R.; Sharma, A.; Vatsyayan, R.; Yadav, S.; Singhal, S.S.; Rauniyar, N.; Prokai, L.; Awasthi, S.; Awasthi, Y.C. Mechanisms of 4-hydroxy-2-nonenal induced pro- and anti-apoptotic signaling. Biochemistry 2010, 49, 6263–6275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaganjac, M.; Cindrić, M.; Jakovčević, A.; Žarković, K.; Žarković, N. Lipid peroxidation in brain tumors. Neurochem. Int. 2021, 149, 105118. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Jiménez, E.; Ramírez-Hernández, A.A.; Santos-Álvarez, J.C.; Velázquez-Enríquez, J.M.; Pina-Canseco, S.; Baltiérrez-Hoyos, R.; Vásquez-Garzón, V.R. Involvement of 4-hydroxy-2-nonenal in the pathogenesis of pulmonary fibrosis. Mol. Cell Biochem. 2021, 476, 4405–4419. [Google Scholar] [CrossRef] [PubMed]
- Skorokhod, O.A.; Davalos-Schafler, D.; Gallo, V.; Valente, E.; Ulliers, D.; Notarpietro, A.; Mandili, G.; Novelli, F.; Persico, M.; Taglialatela-Scafati, O.; et al. Oxidative stress-mediated antimalarial activity of plakortin, a natural endoperoxide from the tropical sponge Plakortis simplex. Free Radic. Biol. Med. 2015, 89, 624–637. [Google Scholar] [CrossRef] [PubMed]
- Carbone, D.L.; Doorn, J.A.; Kiebler, Z.; Sampey, B.P.; Petersen, D.R. Inhibition of Hsp72-mediated protein refolding by 4-hydroxy-2-nonenal. Chem. Res. Toxicol. 2004, 17, 1459–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galligan, J.J.; Fritz, K.S.; Backos, D.S.; Shearn, C.T.; Smathers, R.L.; Jiang, H.; MacLean, K.N.; Reigan, P.R.; Petersen, D.R. Oxidative stress-mediated aldehyde adduction of GRP78 in a mouse model of alcoholic liver disease: Functional independence of ATPase activity and chaperone function. Free Radic. Biol. Med. 2014, 73, 411–420. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.L.; Chen, H.; Wang, J.; Xia, T.; Sun, H.; Yuan, C.H.; Liu, S.L.; Chen, J.B. 4-HNE Induces Apoptosis of Human Retinal Pigment Epithelial Cells by Modifying HSP70. Curr. Med. Sci. 2019, 39, 442–448. [Google Scholar] [CrossRef]
- Polla, B.S.; Kantengwa, S.; François, D.; Salvioli, S.; Franceschi, C.; Marsac, C.; Cossarizza, A. Mitochondria are selective targets for the protective effects of heat shock against oxidative injury. Proc. Natl. Acad. Sci. USA 1996, 93, 6458–6463. [Google Scholar] [CrossRef] [Green Version]
- Khomenko, I.P.; Bakhtina, L.Y.; Zelenina, O.M.; Kruglov, S.V.; Manukhina, E.B.; Bayda, L.A.; Malyshev, I.Y. Role of heat shock proteins HSP70 and HSP32 in the protective effect of adaptation of cultured HT22 hippocampal cells to oxidative stress. Bull. Exp. Biol. Med. 2007, 144, 174–177. [Google Scholar] [CrossRef]
- Watanabe, S.; Ageta-Ishihara, N.; Nagatsu, S.; Takao, K.; Komine, O.; Endo, F.; Miyakawa, T.; Misawa, H.; Takahashi, R.; Kinoshita, M.; et al. SIRT1 overexpression ameliorates a mouse model of SOD1-linked amyotrophic lateral sclerosis via HSF1/HSP70i chaperone system. Mol. Brain 2014, 7, 62. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Wharton, W.; Moseley, P.; Shi, H. Heat shock protein 70 regulates cellular redox status by modulating glutathione-related enzyme activities. Cell Stress Chaperones 2007, 12, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Broome, C.S.; Kayani, A.C.; Palomero, J.; Dillmann, W.H.; Mestril, R.; Jackson, M.J.; McArdle, A. Effect of lifelong overexpression of HSP70 in skeletal muscle on age-related oxidative stress and adaptation after nondamaging contractile activity. Faseb J. 2006, 20, 1549–1551. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Voloboueva, L.A.; Ouyang, Y.; Emery, J.F.; Giffard, R.G. Overexpression of mitochondrial Hsp70/Hsp75 in rat brain protects mitochondria, reduces oxidative stress, and protects from focal ischemia. J. Cereb. Blood Flow Metab. 2009, 29, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.H.; Chang, Y.J.; Lin, S.; Yang, W.Y. Hsc70/Stub1 promotes the removal of individual oxidatively stressed peroxisomes. Nat. Commun. 2020, 11, 5267. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yu, Y.; Gorshkov, B.; Haigh, S.; Bordan, Z.; Weintraub, D.; Rudic, R.D.; Chakraborty, T.; Barman, S.A.; Verin, A.D.; et al. Hsp70 Suppresses Mitochondrial Reactive Oxygen Species and Preserves Pulmonary Microvascular Barrier Integrity Following Exposure to Bacterial Toxins. Front. Immunol. 2018, 9, 1309. [Google Scholar] [CrossRef]
- Pistono, C.; Monti, M.C.; Boiocchi, C.; Berzolari, F.G.; Osera, C.; Mallucci, G.; Cuccia, M.; Pascale, A.; Montomoli, C.; Bergamaschi, R. Response to oxidative stress of peripheral blood mononuclear cells from multiple sclerosis patients and healthy controls. Cell Stress Chaperones 2020, 25, 81–91. [Google Scholar] [CrossRef]
- Yan, L.J.; Christians, E.S.; Liu, L.; Xiao, X.; Sohal, R.S.; Benjamin, I.J. Mouse heat shock transcription factor 1 deficiency alters cardiac redox homeostasis and increases mitochondrial oxidative damage. Embo J. 2002, 21, 5164–5172. [Google Scholar] [CrossRef] [Green Version]
- Dayalan Naidu, S.; Kostov, R.V.; Dinkova-Kostova, A.T. Transcription factors Hsf1 and Nrf2 engage in crosstalk for cytoprotection. Trends. Pharmacol. Sci. 2015, 36, 6–14. [Google Scholar] [CrossRef]
- Girard, P.M.; Peynot, N.; Lelièvre, J.M. Differential correlations between changes to glutathione redox state, protein ubiquitination, and stress-inducible HSPA chaperone expression after different types of oxidative stress. Cell Stress Chaperones 2018, 23, 985–1002. [Google Scholar] [CrossRef]
- Doulias, P.T.; Kotoglou, P.; Tenopoulou, M.; Keramisanou, D.; Tzavaras, T.; Brunk, U.; Galaris, D.; Angelidis, C. Involvement of heat shock protein-70 in the mechanism of hydrogen peroxide-induced DNA damage: The role of lysosomes and iron. Free Radic. Biol. Med. 2007, 42, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Adachi, M.; Liu, Y.; Fujii, K.; Calderwood, S.K.; Nakai, A.; Imai, K.; Shinomura, Y. Oxidative stress impairs the heat stress response and delays unfolded protein recovery. PLoS ONE 2009, 4, e7719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, A.; Moulin, D.; Hupont, S.; Koufany, M.; Netter, P.; Reboul, P.; Jouzeau, J.Y. Oxidative stress-induced expression of HSP70 contributes to the inhibitory effect of 15d-PGJ2 on inducible prostaglandin pathway in chondrocytes. Free Radic. Biol. Med. 2014, 76, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Reeg, S.; Jung, T.; Castro, J.P.; Davies, K.J.A.; Henze, A.; Grune, T. The molecular chaperone Hsp70 promotes the proteolytic removal of oxidatively damaged proteins by the proteasome. Free Radic. Biol. Med. 2016, 99, 153–166. [Google Scholar] [CrossRef] [Green Version]
- Kiffin, R.; Christian, C.; Knecht, E.; Cuervo, A.M. Activation of chaperone-mediated autophagy during oxidative stress. Mol. Biol. Cell 2004, 15, 4829–4840. [Google Scholar] [CrossRef] [Green Version]
- Dodson, M.; Darley-Usmar, V.; Zhang, J. Cellular metabolic and autophagic pathways: Traffic control by redox signaling. Free Radic. Biol. Med. 2013, 63, 207–221. [Google Scholar] [CrossRef] [Green Version]
- Zemanovic, S.; Ivanov, M.V.; Ivanova, L.V.; Bhatnagar, A.; Michalkiewicz, T.; Teng, R.J.; Kumar, S.; Rathore, R.; Pritchard, K.A., Jr.; Konduri, G.G.; et al. Dynamic Phosphorylation of the C Terminus of Hsp70 Regulates the Mitochondrial Import of SOD2 and Redox Balance. Cell Rep. 2018, 25, 2605–2616. [Google Scholar] [CrossRef] [Green Version]
- Xu, N.W.; Chen, Y.; Liu, W.; Chen, Y.J.; Fan, Z.M.; Liu, M.; Li, L.J. Inhibition of JAK2/STAT3 Signaling Pathway Suppresses Proliferation of Burkitt’s Lymphoma Raji Cells via Cell Cycle Progression, Apoptosis, and Oxidative Stress by Modulating HSP70. Med. Sci. Monit. 2018, 24, 6255–6263. [Google Scholar] [CrossRef]
- Hensen, S.M.; Heldens, L.; van Enckevort, C.M.; van Genesen, S.T.; Pruijn, G.J.; Lubsen, N.H. Activation of the antioxidant response in methionine deprived human cells results in an HSF1-independent increase in HSPA1A mRNA levels. Biochimie 2013, 95, 1245–1251. [Google Scholar] [CrossRef]
- Smith, M.R.; Fernandes, J.; Go, Y.M.; Jones, D.P. Redox dynamics of manganese as a mitochondrial life-death switch. Biochem. Biophys. Res. Commun. 2017, 482, 388–398. [Google Scholar] [CrossRef] [Green Version]
- Afolayan, A.J.; Teng, R.J.; Eis, A.; Rana, U.; Broniowska, K.A.; Corbett, J.A.; Pritchard, K.; Konduri, G.G. Inducible HSP70 regulates superoxide dismutase-2 and mitochondrial oxidative stress in the endothelial cells from developing lungs. Am. J. Physiol. Lung Cell Mol. Physiol. 2014, 306, L351–L360. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Amick, J.; Chakravarti, R.; Santarriaga, S.; Schlanger, S.; McGlone, C.; Dare, M.; Nix, J.C.; Scaglione, K.M.; Stuehr, D.J.; et al. A bipartite interaction between Hsp70 and CHIP regulates ubiquitination of chaperoned client proteins. Structure 2015, 23, 472–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Shi, X.; Sheng, K.; Han, G.; Li, W.; Zhao, Q.; Jiang, B.; Feng, J.; Li, J.; Gu, Y. PI3K/Akt signaling transduction pathway, erythropoiesis and glycolysis in hypoxia (Review). Mol. Med. Rep. 2019, 19, 783–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, R.; Zhang, Y.G.; Sun, J. STAT3 activation in infection and infection-associated cancer. Mol. Cell Endocrinol. 2017, 451, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Alas, S.; Bonavida, B. Inhibition of constitutive STAT3 activity sensitizes resistant non-Hodgkin’s lymphoma and multiple myeloma to chemotherapeutic drug-mediated apoptosis. Clin. Cancer Res. 2003, 9, 316–326. [Google Scholar] [PubMed]
- Madamanchi, N.R.; Li, S.; Patterson, C.; Runge, M.S. Reactive oxygen species regulate heat-shock protein 70 via the JAK/STAT pathway. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Marinho, H.S.; Real, C.; Cyrne, L.; Soares, H.; Antunes, F. Hydrogen peroxide sensing, signaling and regulation of transcription factors. Redox Biol. 2014, 2, 535–562. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Bharadwaj, S.; O’Carroll, R.; Ovsenek, N. HSP90 interacts with and regulates the activity of heat shock factor 1 in Xenopus oocytes. Mol. Cell Biol. 1998, 18, 4949–4960. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Mosser, D.D.; Morimoto, R.I. Molecular chaperones as HSF1-specific transcriptional repressors. Genes Dev. 1998, 12, 654–666. [Google Scholar] [CrossRef] [Green Version]
- Kmiecik, S.W.; Mayer, M.P. Molecular mechanisms of heat shock factor 1 regulation. Trends. Biochem. Sci. 2021. [Google Scholar] [CrossRef]
- Paul, S.; Ghosh, S.; Mandal, S.; Sau, S.; Pal, M. NRF2 transcriptionally activates the heat shock factor 1 promoter under oxidative stress and affects survival and migration potential of MCF7 cells. J. Biol. Chem. 2018, 293, 19303–19316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Ahn, Y.H.; Benjamin, I.J.; Honda, T.; Hicks, R.J.; Calabrese, V.; Cole, P.A.; Dinkova-Kostova, A.T. HSF1-dependent upregulation of Hsp70 by sulfhydryl-reactive inducers of the KEAP1/NRF2/ARE pathway. Chem. Biol. 2011, 18, 1355–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hensen, S.M.; Heldens, L.; van Genesen, S.T.; Pruijn, G.J.; Lubsen, N.H. A delayed antioxidant response in heat-stressed cells expressing a non-DNA binding HSF1 mutant. Cell Stress Chaperones 2013, 18, 455–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiktorowicz, J.E.; Stafford, S.; Rea, H.; Urvil, P.; Soman, K.; Kurosky, A.; Perez-Polo, J.R.; Savidge, T.C. Quantification of Cysteinyl S-Nitrosylation by Fluorescence in Unbiased Proteomic Studies. Biochemistry 2011, 50, 5601–5614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansong, C.; Wu, S.; Meng, D.; Liu, X.; Brewer, H.M.; Kaiser, B.L.D.; Nakayasu, E.; Cort, J.R.; Pevzner, P.; Smith, R.; et al. Top-down proteomics reveals a unique protein S-thiolation switch in Salmonella Typhimurium in response to infection-like conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 10153–10158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michelet, L.; Zaffagnini, M.; Vanacker, H.; Le Maréchal, P.; Marchand, C.; Schroda, M.; Lemaire, S.; Decottignies, P. In Vivo Targets of S-Thiolation in Chlamydomonas reinhardtii. J. Biol. Chem. 2008, 283, 21571–21578. [Google Scholar] [CrossRef]
- Huang, J.; Willems, P.; Wei, B.; Tian, C.; Ferreira, R.B.; Bodra, N.; Gache, S.A.M.; Wahni, K.; Liu, K.; Vertommen, D.; et al. Mining for protein S-sulfenylation in Arabidopsis uncovers redox-sensitive sites. Proc. Natl. Acad. Sci. USA 2019, 116, 21256–21261. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Huang, X.; Chen, L.; Sun, X.; Lu, C.; Zhang, L.; Wang, Y.; Zuo, J. Site-Specific Nitrosoproteomic Identification of Endogenously S-Nitrosylated Proteins in Arabidopsis. Plant Physiol. 2015, 167, 1731–1746. [Google Scholar] [CrossRef] [Green Version]
- Chaki, M.; Shekariesfahlan, A.; Ageeva, A.; Mengel, A.; von Toerne, C.; Durner, J.; Lindermayr, C. Identification of nuclear target proteins for S-nitrosylation in pathogen-treated Arabidopsis thaliana cell cultures. Plant Sci. 2015, 238, 115–126. [Google Scholar] [CrossRef]
- Fares, A.; Rossignol, M.; Peltier, J.-B. Proteomics investigation of endogenous S-nitrosylation in Arabidopsis. Biochem. Biophys. Res. Commun. 2011, 416, 331–336. [Google Scholar] [CrossRef]
- Aroca, A.; Benito, J.M.; Gotor, C.; Romero, L.C. Persulfidation proteome reveals the regulation of protein function by hydrogen sulfide in diverse biological processes in Arabidopsis. J. Exp. Bot. 2017, 68, 4915–4927. [Google Scholar] [CrossRef] [Green Version]
- Gong, B.; Shi, Q. Identifying S-nitrosylated proteins and unraveling S-nitrosoglutathione reductase-modulated sodic alkaline stress tolerance in Solanum lycopersicum L. Plant Physiol. Biochem. 2019, 142, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Gong, B.; Yan, Y.; Zhang, L.; Cheng, F.; Liu, Z.; Shi, Q. Unravelling GSNOR-Mediated S-Nitrosylation and Multiple Developmental Programs in Tomato Plants. Plant Cell Physiol. 2019, 60, 2523–2537. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Chen, Y.; Yang, F.; Zhang, L.; Gong, Y.; Adilijiang, G.; Gao, Y.; Deng, H. Development of a Clickable Probe for Profiling of Protein Glutathionylation in the Central Cellular Metabolism of E. coli and Drosophila. Chem. Biol. 2015, 22, 1461–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, D.; Gaffrey, M.J.; Guo, J.; Hatchell, K.E.; Chu, R.K.; Clauss, T.R.; Aldrich, J.T.; Wu, S.; Purvine, S.; Camp, D.G.; et al. Proteomic identification and quantification of S-glutathionylation in mouse macrophages using resin-assisted enrichment and isobaric labeling. Free Radic. Biol. Med. 2013, 67, 460–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, J.; Kodali, V.K.; Gaffrey, M.J.; Guo, J.; Chu, R.K.; Camp, D.G.; Smith, R.D.; Thrall, B.D.; Qian, W.-J. Quantitative Profiling of Protein S-Glutathionylation Reveals Redox-Dependent Regulation of Macrophage Function during Nanoparticle-Induced Oxidative Stress. ACS Nano 2015, 10, 524–538. [Google Scholar] [CrossRef] [Green Version]
- Fowler, N.J.; Blanford, C.F.; de Visser, S.; Warwicker, J. Features of reactive cysteines discovered through computation: From kinase inhibition to enrichment around protein degrons. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Vanhecke, G.C.; Abeywardana, M.Y.; Huang, B.; Ahn, Y.; Young-Hoon, A. Isotopically Labeled Clickable Glutathione to Quantify Protein S-Glutathionylation. ChemBioChem 2019, 21, 853–859. [Google Scholar] [CrossRef]
- Gould, N.S.; Evans, P.; Martínez-Acedo, P.; Marino, S.M.; Gladyshev, V.N.; Carroll, K.S.; Ischiropoulos, H. Site-Specific Proteomic Mapping Identifies Selectively Modified Regulatory Cysteine Residues in Functionally Distinct Protein Networks. Chem. Biol. 2015, 22, 965–975. [Google Scholar] [CrossRef] [Green Version]
- Su, D.; Shukla, A.K.; Chen, B.; Kim, J.-S.; Nakayasu, E.; Qu, Y.; Aryal, U.; Weitz, K.; Clauss, T.R.; Monroe, M.E.; et al. Quantitative site-specific reactivity profiling of S-nitrosylation in mouse skeletal muscle using cysteinyl peptide enrichment coupled with mass spectrometry. Free Radic. Biol. Med. 2013, 57, 68–78. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-J.; Ku, W.-C.; Lin, P.-Y.; Chou, H.-C.; Khoo, K.-H. S-Alkylating Labeling Strategy for Site-Specific Identification of theS-Nitrosoproteome. J. Proteome Res. 2010, 9, 6417–6439. [Google Scholar] [CrossRef]
- Lee, T.-Y.; Chen, Y.-J.; Lu, C.-T.; Ching, W.-C.; Teng, Y.-C.; Huang, H.-D. dbSNO: A database of cysteine S-nitrosylation. Bioinformatics 2012, 28, 2293–2295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaręba-Kozioł, M.; Szwajda, A.; Dadlez, M.; Wysłouch-Cieszyńska, A.; Lalowski, M. Global Analysis of S-nitrosylation Sites in the Wild Type (APP) Transgenic Mouse Brain-Clues for Synaptic Pathology. Mol. Cell. Proteom. 2014, 13, 2288–2305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibáñez-Vea, M.; Huang, H.; De Morentin, X.M.; Pérez, E.; Gato, M.; Zuazo, M.; Arasanz, H.; Fernandez-Irigoyen, J.; Santamaría, E.; Fernández-Hinojal, G.; et al. Characterization of Macrophage Endogenous S-Nitrosoproteome Using a Cysteine-Specific Phosphonate Adaptable Tag in Combination with TiO2 Chromatography. J. Proteome Res. 2018, 17, 1172–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohr, M.J.; Aponte, A.M.; Sun, J.; Wang, G.; Murphy, E.; Gucek, M.; Steenbergen, C. Characterization of potentialS-nitrosylation sites in the myocardium. Am. J. Physiol. Circ. Physiol. 2011, 300, H1327–H13352011. [Google Scholar] [CrossRef] [Green Version]
- Doulias, P.-T.; Tenopoulou, M.; Greene, J.L.; Raju, K.; Ischiropoulos, H. Nitric Oxide Regulates Mitochondrial Fatty Acid Metabolism Through Reversible Protein S -Nitrosylation. Sci. Signal. 2013, 6, rs1. [Google Scholar] [CrossRef] [Green Version]
- Kohr, M.; Aponte, A.M.; Sun, J.; Gucek, M.; Steenbergen, C.; Murphy, E. Measurement of S -Nitrosylation Occupancy in the Myocardium With Cysteine-Reactive Tandem Mass Tags. Circ. Res. 2012, 111, 1308–1312. [Google Scholar] [CrossRef] [Green Version]
- Qu, Z.; Meng, F.; Bomgarden, R.D.; Viner, R.I.; Li, J.; Rogers, J.C.; Cheng, J.; Greenlief, C.M.; Cui, J.; Lubahn, D.B.; et al. Proteomic Quantification and Site-Mapping of S-Nitrosylated Proteins Using Isobaric iodoTMT Reagents. J. Proteome Res. 2014, 13, 3200–3211. [Google Scholar] [CrossRef]
- Gu, L.; Robinson, R.A.S. High-throughput endogenous measurement of S-nitrosylation in Alzheimer's disease using oxidized cysteine-selective cPILOT. Anal. 2016, 141, 3904–3915. [Google Scholar] [CrossRef] [Green Version]
- Raju, K.; Doulias, P.-T.; Evans, P.; Krizman, E.N.; Jackson, J.G.; Horyn, O.; Daikhin, Y.; Nissim, I.; Yudkoff, M.; Nissim, I.; et al. Regulation of brain glutamate metabolism by nitric oxide and S-nitrosylation. Sci. Signal. 2015, 8, ra68. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.G.; Aldous, S.G.; Andreassi, C.; Cuda, G.; Gaspari, M.; Riccio, A. Proteomic analysis of S-nitrosylated nuclear proteins in rat cortical neurons. Sci. Signal. 2018, 11. [Google Scholar] [CrossRef] [Green Version]
- Pan, K.-T.; Chen, Y.-Y.; Pu, T.-H.; Chao, Y.-S.; Yang, C.-Y.; Bomgarden, R.D.; Rogers, J.C.; Meng, T.-C.; Khoo, K.-H. Mass Spectrometry-Based Quantitative Proteomics for Dissecting Multiplexed Redox Cysteine Modifications in Nitric Oxide-Protected Cardiomyocyte Under Hypoxia. Antioxidants Redox Signal. 2014, 20, 1365–1381. [Google Scholar] [CrossRef]
- Zhang, H.-H.; Lechuga, T.J.; Chen, Y.; Yang, Y.; Huang, L.; Chen, D.-B. Quantitative Proteomics Analysis of VEGF-Responsive Endothelial Protein S-Nitrosylation Using Stable Isotope Labeling by Amino Acids in Cell Culture (SILAC) and LC-MS/MS. Biol. Reprod. 2016, 94, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Luo, X.; Li, Y.; Chen, L.; Ma, W.; Huang, J.; Cui, J.; Zhao, Y.; Xue, Y.; Zuo, Z.; et al. DeepNitro: Prediction of Protein Nitration and Nitrosylation Sites by Deep Learning. Genom. Proteom. Bioinform. 2018, 16, 294–306. [Google Scholar] [CrossRef]
- Chou, C.-C.; Chiang, B.-Y.; Lin, J.C.-Y.; Pan, K.-T.; Lin, C.-H.; Khoo, K.-H. Characteristic Tandem Mass Spectral Features Under Various Collision Chemistries for Site-Specific Identification of Protein S-Glutathionylation. J. Am. Soc. Mass Spectrom. 2014, 26, 120–132. [Google Scholar] [CrossRef] [PubMed]
- Konstantinidis, D.; Paletas, K.; Koliakos, G.; Kaloyianni, M. The ambiguous role of the Na+–H+ exchanger isoform 1 (NHE1) in leptin-induced oxidative stress in human monocytes. Cell Stress Chaperon- 2009, 14, 591–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoppe, G.; Chai, Y.-C.; Crabb, J.W.; Sears, J. Protein s-glutathionylation in retinal pigment epithelium converts heat shock protein 70 to an active chaperone. Exp. Eye Res. 2004, 78, 1085–1092. [Google Scholar] [CrossRef]
- Majmudar, J.D.; Konopko, A.M.; Labby, K.J.; Tom, C.T.M.B.; Crellin, J.E.; Prakash, A.; Martin, B.R. Harnessing Redox Cross-Reactivity To Profile Distinct Cysteine Modifications. J. Am. Chem. Soc. 2016, 138, 1852–1859. [Google Scholar] [CrossRef] [Green Version]
- Forrester, M.T.; Thompson, J.W.; Foster, M.W.; Nogueira, L.; Moseley, M.A.; Stamler, J.S. Proteomic analysis of S-nitrosylation and denitrosylation by resin-assisted capture. Nat. Biotechnol. 2009, 27, 557–559. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Ding, J.; Wu, L.-Y.; Chou, K.-C. iSNO-PseAAC: Predict Cysteine S-Nitrosylation Sites in Proteins by Incorporating Position Specific Amino Acid Propensity into Pseudo Amino Acid Composition. PLoS ONE 2013, 8, e55844. [Google Scholar] [CrossRef] [Green Version]
- Ben-Lulu, S.; Ziv, T.; Weisman-Shomer, P.; Benhar, M. Nitrosothiol-Trapping-Based Proteomic Analysis of S-Nitrosylation in Human Lung Carcinoma Cells. PLoS ONE 2017, 12, e0169862. [Google Scholar] [CrossRef]
- Liu, M.; Hou, J.; Huang, L.; Huang, X.; Heibeck, T.H.; Zhao, R.; Pasa-Tolic, L.; Smith, R.D.; Li, Y.; Fu, K.; et al. Site-Specific Proteomics Approach for Study Protein S-Nitrosylation. Anal. Chem. 2010, 82, 7160–7168. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Li, Y.; Huang, X.; Wei, M.; Huang, Y.; Tang, Z.; Huang, H.; Zhou, W.; Wang, Y.; Hu, J. Extensive protein S-nitrosylation associated with human pancreatic ductal adenocarcinoma pathogenesis. Cell Death Dis. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cellular Processes | Critical Signaling Molecules Modified by ROS |
|---|---|
| proliferation and survival | mitogen-activated protein kinases (MAPKs), phosphatidylinositol-4,5-bisphosphate 3-kinase (PI3K), phosphatase and tensin homolog deleted on chromosome ten (PTEN) and protein tyrosine phosphatases |
| redox homeostasis | thioredoxin, peroxiredoxin, redox factor-1 (Ref-1) and Kelch-like ECH-associated protein 1 (Keap1)/nuclear factor erythroid 2-related factor 2 (Nrf2) |
| mitochondrial oxidative stress and aging | p66Shc |
| iron homeostasis | iron response element–iron regulatory protein (IRE-IRP) containing iron–sulfur cluster |
| DNA damage response | ataxia telangiectasia mutated (ATM) |
| Hsp70 Homolog | Cysteine Residues (Entry in UniProtKB) |
|---|---|
| Escherichia coli DnaK | Cys15 (P0A6Y8) |
| Escherichia coli HscA (Hsc66) | Cys315, Cys448 (P0A6Z1) |
| Escherichia coli HscC (Hsc62) | Cys236, Cys242, Cys261, Cys344, Cys360 (P77319) |
| Saccharomyces cerevisiae Ssa1 (cytosol) | Cys15, Cys264, Cys303 (P10591) |
| Saccharomyces cerevisiae Ssa2 (cytosol) | Cys15, Cys264, Cys303 (P10592) |
| Saccharomyces cerevisiae Ssa3 (cytosol) | Cys15, Cys304 (P09435) |
| Saccharomyces cerevisiae Ssa4 (cytosol) | Cys15, Cys304 (P22202) |
| Saccharomyces cerevisiae Ssb1 (cytosol) | Cys20, Cys435, Cys454 (P11484) |
| Saccharomyces cerevisiae Ssb2 (cytosol) | Cys20, Cys454 (P40150) |
| Saccharomyces cerevisiae Sse1 (cytosol Hsp110) | Cys142, Cys211, Cys228, Cys380, Cys484 (P32589) |
| Saccharomyces cerevisiae Sse2 (cytosol Hsp110) | Cys142, Cys211, Cys380, Cys484 (P32590) |
| Saccharomyces cerevisiae Ssz1 (cytosol) | Cys81, Cys86 (P38788) |
| Saccharomyces cerevisiae Ssc1 (mitochondria) | None (P0CS90) |
| Saccharomyces cerevisiae Ssc2 (Ssq1) (mitochondria) | Cys134 (Q05931) |
| Saccharomyces cerevisiae Ssc3 (Ecm10) (mitochondria) | None (P39987) |
| Saccharomyces cerevisiae Ssd1 (Kar2, BiP, Grp78) (ER) | Cys63 (P16474) |
| Saccharomyces cerevisiae Lhs1 (Grp170) (ER) | Cys520, Cys545, Cys547 (P36016) |
| Human HspA1A (Hsp72) (cytosol, nucleus, cell membrane, extracellular exosomes) | Cys17, Cys267, Cys306, Cys574, Cys603 (P0DMV8) |
| Human HspA1B (Hsp72) (cytosol, nucleus, extracellular exosomes) | Cys17, Cys267, Cys306, Cys574, Cys603 (P0DMV9) |
| Human HspA1L (cytosol, nucleus) | Cys19, Cys269, Cys308, Cys576, Cys605, Cys617, Cys622 (P34931) |
| Human HspA2 (cytosol, nucleus, cell membrane, extracellular exosomes) | Cys18, Cys191, Cys270, Cys577, Cys606 (P54652) |
| Human HspA4 (Apg2) (cytosol, extracellular exosome, mitochondrion, nucleus) | Cys13, Cys34, Cys38, Cys140, Cys146, Cys167, Cys213, Cys245, Cys270, Cys290, Cys310, Cys376, Cys380, Cys417, Cys779 (P34932) |
| Human HspA4L (Apg1, Osp94) (cytosol, nucleus) | Cys13, Cys34, Cys38, Cys140, Cys167, Cys213, Cys245, Cys270, Cys290, Cys310, Cys376, Cys380, Cys417, Cys421, Cys540, Cys589, Cys740, Cys782 (O95757) |
| Human HspA5 (BiP, Grp78) (ER, extracellular exosomes) | Cys41, Cys420 (P11021) |
| Human HspA6 (Hsp70B’) (cytosol, extracellular exosomes) | Cys19, Cys108, Cys269, Cys308, Cys387, Cys576, Cys605, Cys624 (P17066) |
| Human HspA7 (Hsp70B) (blood microparticles, extracellular exosomes) | Cys19, Cys108, Cys269, Cys308 (P48741) |
| human HspA8 (Hsc70, Hsc73) (cytosol, nucleus, cell membrane, extracellular exosomes) | Cys17, Cys267, Cys574, Cys603 (P11142) |
| Human HspA9 (Grp75, mt-Hsp70) (mitochondria, nucleus) | Cys66, Cys317, Cys366, Cys487, Cys608 (P38646) |
| Human HspA12A (extracellular exosomes, nucleus) | Cys80, Cys246, Cys502, Cys564, Cys621 (O43301) |
| Human HspA12B (endothelial cells, intracellular, blood plasma) | Cys36, Cys106, Cys250, Cys321, Cys365, Cys450, Cys570, Cys595, Cys610, Cys611, Cys626, Cys639 (Q96MM6) |
| Human HspA13 (Stch) (ER, extracellular exosomes, microsomes) | Cys43 (P48723) |
| Human HspA14 (Hsp60, Hsp70L1) (cytosol, membrane) | Cys10, Cys14, Cys89, Cys280, Cys293, Cys304, Cys311, Cys335, Cys394, Cys440, Cys492, Cys500 (Q0VDF9) |
| Human HspH1 (Hsp105, Hsp110) (microtubule, cytosol, extracellular region or secreted, nucleus) | Cys13, Cys34, Cys48, Cys140, Cys167, Cys213, Cys245, Cys270, Cys290, Cys310, Cys376, Cys380, Cys516, Cys650, Cys658, Cys796, Cys845 (Q92598) |
| Human Hyou1 (Grp170, Orp150) (ER, extracellular region or secreted) | Cys15, Cys240, Cys352, Cys805 (Q9Y4L1) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Gong, W.; Wu, S.; Perrett, S. Hsp70 in Redox Homeostasis. Cells 2022, 11, 829. https://doi.org/10.3390/cells11050829
Zhang H, Gong W, Wu S, Perrett S. Hsp70 in Redox Homeostasis. Cells. 2022; 11(5):829. https://doi.org/10.3390/cells11050829
Chicago/Turabian StyleZhang, Hong, Weibin Gong, Si Wu, and Sarah Perrett. 2022. "Hsp70 in Redox Homeostasis" Cells 11, no. 5: 829. https://doi.org/10.3390/cells11050829