
Enhanced Autophagic Flux, Suppressed Apoptosis and Reduced Macrophage Infiltration by Dasatinib in Kidneys of Obese Mice

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Size Estimation
2.2. Experimental Design of the Study, Animals and Diet
2.3. Drug Preparation
2.4. Blood and Tissue Sampling
2.5. Assessment of Glucose, Kidney Function Tests and Lipid Profile
2.6. mRNA Quantification of the Studied Genes
2.7. Detection of the Studied Proteins Expression by Western Blotting
2.8. Histopathology
2.9. Histopathological Assessment of Renal Cortical Damage
2.10. Immunohistochemical Staining
2.11. Morphometric Analysis of Immunohistochemical Results
2.12. Statistical Analysis
3. Results
3.1. Results of Final Body Weights and Kidney Weights
3.2. Results of Fasting Glucose Level and Renal Function Tests
3.3. Effect of Dasatinib on Lipid Profile in Obese Mice
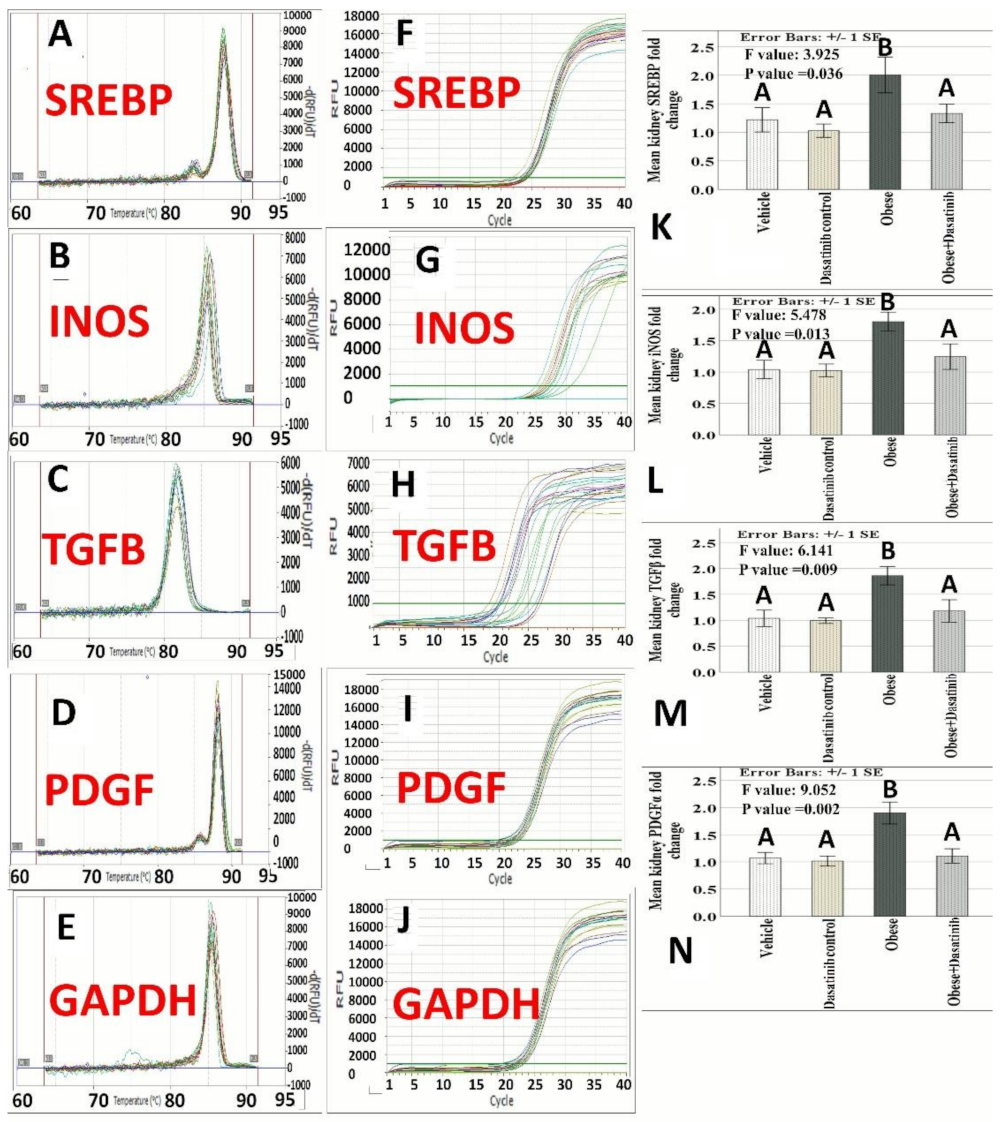
3.4. The Expression Profile of iNOS, TGF-β, SREBP and PDGFA
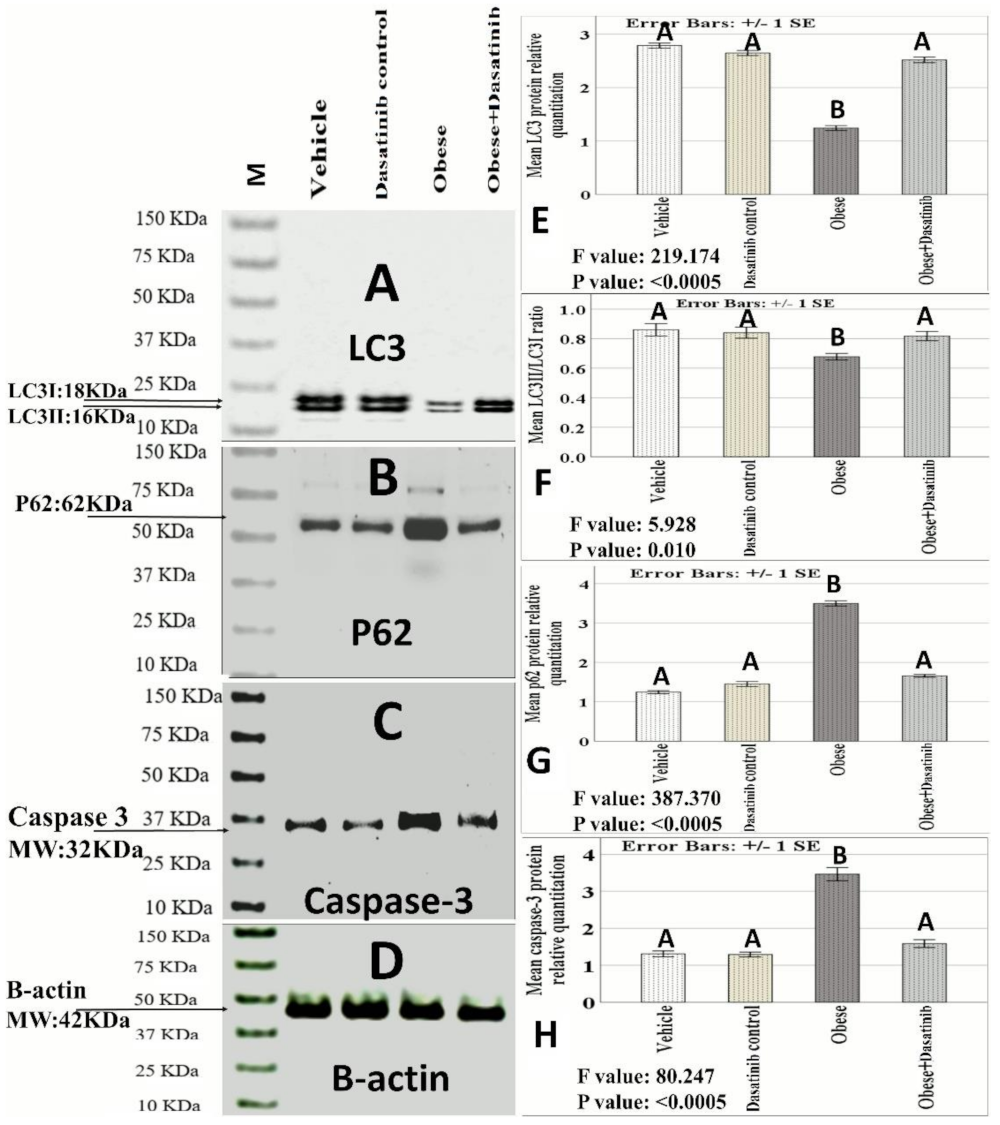
3.5. Effect of Dasatinib on Protein Expression as Assessed by Western Blotting
3.6. Effect of Dasatinib on Renal Histopathological Changes Induced by OD
3.7. The Histopathological Score
3.8. Immunohistochemical Results
3.9. Morphometric Assessment of Immunohistochemical Findings
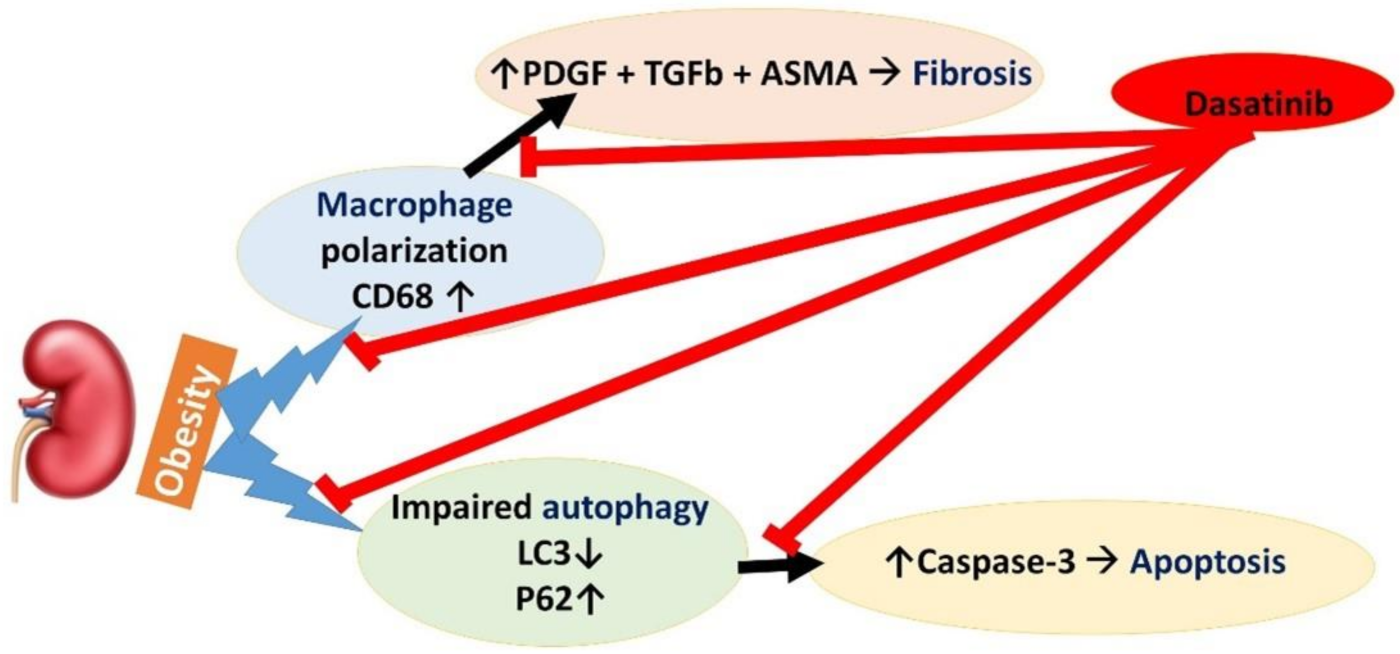
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, S.; Cao, C.; Deng, T.; Zhou, Z. Obesity-Related Glomerulopathy: A Latent Change in Obesity Requiring More Attention. Kidney Blood Press. Res. 2020, 45, 510–522. [Google Scholar] [CrossRef] [PubMed]
- Altunkaynak, M.E.; Özbek, E.; Altunkaynak, B.Z.; Can, İ.; Unal, D.; Unal, B. The effects of high-fat diet on the renal structure and morphometric parametric of kidneys in rats. J. Anat. 2008, 212, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Deng, L.; Wang, L.; Zhang, Y.; Weng, Q.; Yin, H.; Pan, Y.; Tong, C.; Wang, J.; Liang, G. Inhibition of Mitogen-Activated Protein Kinases/Nuclear Factor κB–Dependent Inflammation by a Novel Chalcone Protects the Kidney from High Fat Diet–Induced Injuries in Mice. J. Pharmacol. Exp. Ther. 2015, 355, 235–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salim, H.M.; Kurnia, L.F.; Bintarti, T.W.; Handayani, H. The Effects of High-fat Diet on Histological Changes of Kidneys in Rats. Biomol. Health Sci. J. 2018, 1, 109–112. [Google Scholar] [CrossRef] [Green Version]
- Kok, H.M.; Falke, L.L.; Goldschmeding, R.; Nguyen, T.Q. Targeting CTGF, EGF and PDGF pathways to prevent progression of kidney disease. Nat. Rev. Nephrol. 2014, 10, 700–711. [Google Scholar] [CrossRef]
- Martinez-Martinez, E.; Ibarrola, J.; Calvier, L.; Fernandez-Celis, A.; Leroy, C.; Cachofeiro, V.; Rossignol, P.; Lopez-Andres, N. Galectin-3 Blockade Reduces Renal Fibrosis in Two Normotensive Experimental Models of Renal Damage. PLoS ONE 2016, 11, e0166272. [Google Scholar] [CrossRef]
- Aroor, A.R.; Habibi, J.; Nistala, R.; Ramirez-Perez, F.I.; Martinez-Lemus, L.A.; Jaffe, I.Z.; Sowers, J.R.; Jia, G.; Whaley-Connell, A. Diet-Induced Obesity Promotes Kidney Endothelial Stiffening and Fibrosis Dependent on the Endothelial Mineralocorticoid Receptor. Hypertension 2019, 73, 849–858. [Google Scholar] [CrossRef]
- Nguyen, D.; Ping, F.; Mu, W.; Hill, P.; Atkins, R.C.; Chadban, S.J. Macrophage accumulation in human progressive diabetic nephropathy. Nephrology 2006, 11, 226–231. [Google Scholar] [CrossRef]
- Hunley, T.E.; Ma, L.J.; Kon, V. Scope and mechanisms of obesity-related renal disease. Curr. Opin. Nephrol. Hypertens. 2010, 19, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Lu, Y.H.; Tang, C.; Xue, M.; Li, X.Y.; Chang, Y.P.; Cheng, Y.; Li, T.; Yu, X.C.; Sun, B.; et al. Calcium Dobesilate Restores Autophagy by Inhibiting the VEGF/PI3K/AKT/mTOR Signaling Pathway. Front. Pharmacol. 2019, 10, 886. [Google Scholar] [CrossRef] [Green Version]
- Aplenc, R.; Blaney, S.M.; Strauss, L.C.; Balis, F.M.; Shusterman, S.; Ingle, A.M.; Agrawal, S.; Sun, J.; Wright, J.J.; Adamson, P.C. Pediatric phase I trial and pharmacokinetic study of dasatinib: A report from the children’s oncology group phase I consortium. J. Clin. Oncol. 2011, 29, 839. [Google Scholar] [CrossRef]
- Sasaki, K.; Lahoti, A.; Jabbour, E.; Jain, P.; Pierce, S.; Borthakur, G.; Daver, N.; Kadia, T.; Pemmaraju, N.; Ferrajoli, A.; et al. Clinical Safety and Efficacy of Nilotinib or Dasatinib in Patients with Newly Diagnosed Chronic-Phase Chronic Myelogenous Leukemia and Pre-Existing Liver and/or Renal Dysfunction. Clin. Lymphoma Myeloma Leuk. 2016, 16, 152–162. [Google Scholar] [CrossRef] [Green Version]
- Elsayed, H.R.H.; El-Nablaway, M.; Othman, B.H.; Abdalla, A.M.; El Nashar, E.M.; Abd-Elmonem, M.M.; El-Gamal, R. Can Dasatinib Ameliorate the Hepatic changes, Induced by Long Term Western Diet, in Mice? Ann. Anat. 2021, 234, 151626. [Google Scholar] [CrossRef]
- Tanaka, H.; Hino, H.; Moriya, S.; Kazama, H.; Miyazaki, M.; Takano, N.; Hiramoto, M.; Tsukahara, K.; Miyazawa, K. Comparison of autophagy inducibility in various tyrosine kinase inhibitors and their enhanced cytotoxicity via inhibition of autophagy in cancer cells in combined treatment with azithromycin. Biochem. Biophys. Rep. 2020, 22, 100750. [Google Scholar] [CrossRef]
- Naqvi, K.; Jabbour, E.; Skinner, J.; Yilmaz, M.; Ferrajoli, A.; Bose, P.; Thompson, P.; Alvarado, Y.; Jain, N.; Takahashi, K. Early results of lower dose dasatinib (50 mg daily) as frontline therapy for newly diagnosed chronic-phase chronic myeloid leukemia. Cancer 2018, 124, 2740–2747. [Google Scholar] [CrossRef] [Green Version]
- Abbas, A.; Mirza, M.M.; Ganti, A.K.; Tendulkar, K. Renal Toxicities of Targeted Therapies. Target Oncol. 2015, 10, 487–499. [Google Scholar] [CrossRef]
- Hickson, L.J.; Langhi Prata, L.G.P.; Bobart, S.A.; Evans, T.K.; Giorgadze, N.; Hashmi, S.K.; Herrmann, S.M.; Jensen, M.D.; Jia, Q.; Jordan, K.L.; et al. Senolytics decrease senescent cells in humans: Preliminary report from a clinical trial of Dasatinib plus Quercetin in individuals with diabetic kidney disease. eBioMedicine 2019, 47, 446–456. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.R.; Puranik, A.S.; Jiang, K.; Chen, X.; Zhu, X.Y.; Taylor, I.; Khodadadi-Jamayran, A.; Lerman, A.; Hickson, L.J.; Childs, B.G.; et al. Progressive Cellular Senescence Mediates Renal Dysfunction in Ischemic Nephropathy. J. Am. Soc. Nephrol. 2021, 32, 1987–2004. [Google Scholar] [CrossRef]
- Hassan, N.M.E.; Shehatou, G.S.G.; Kenawy, H.I.; Said, E. Dasatinib mitigates renal fibrosis in a rat model of UUO via inhibition of Src/STAT-3/NF-kappaB signaling. Environ. Toxicol. Pharmacol. 2021, 84, 103625. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.G. Statistical power analyses using G*Power 3.1: Tests for correlation and regression analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Shao, X.; He, J.; Golos, M.; Shi, B. Role of the mTORFOXO1 pathway in obesity-associated renal tubulointerstitial inflammation. Mol. Med. Rep. 2019, 19, 1284–1293. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Lin, S.; Zhang, L.; Li, Y. Resveratrol prevents renal lipotoxicity in high-fat diet-treated mouse model through regulating PPAR-alpha pathway. Mol. Cell. Biochem. 2016, 411, 143–150. [Google Scholar] [CrossRef]
- Xie, X.; Yi, W.; Zhang, P.; Wu, N.; Yan, Q.; Yang, H.; Tian, C.; Xiang, S.; Du, M.; Getachew Assefa, E.; et al. Green Tea Polyphenols, Mimicking the Effects of Dietary Restriction, Ameliorate High-Fat Diet-Induced Kidney Injury via Regulating Autophagy Flux. Nutrients 2017, 9, 497. [Google Scholar] [CrossRef] [Green Version]
- Rosas-Villegas, A.; Sanchez-Tapia, M.; Avila-Nava, A.; Ramirez, V.; Tovar, A.R.; Torres, N. Differential Effect of Sucrose and Fructose in Combination with a High Fat Diet on Intestinal Microbiota and Kidney Oxidative Stress. Nutrients 2017, 9, 393. [Google Scholar] [CrossRef]
- Szeto, H.H.; Liu, S.; Soong, Y.; Alam, N.; Prusky, G.T.; Seshan, S.V. Protection of mitochondria prevents high-fat diet-induced glomerulopathy and proximal tubular injury. Kidney Int. 2016, 90, 997–1011. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Yu, Y.; Deng, J.; Zhang, C.; Zhang, J.; Cheng, Y.; Luo, X.; Han, B.; Yang, H. Sesamin Ameliorates High-Fat Diet-Induced Dyslipidemia and Kidney Injury by Reducing Oxidative Stress. Nutrients 2016, 8, 276. [Google Scholar] [CrossRef] [Green Version]
- Ingvorsen, C.; Karp, N.; Lelliott, C. The role of sex and body weight on the metabolic effects of high-fat diet in C57BL/6N mice. Nutr. Diabetes 2017, 7, e261. [Google Scholar] [CrossRef]
- Enriori, P.J.; Evans, A.E.; Sinnayah, P.; Jobst, E.E.; Tonelli-Lemos, L.; Billes, S.K.; Glavas, M.M.; Grayson, B.E.; Perello, M.; Nillni, E.A. Diet-induced obesity causes severe but reversible leptin resistance in arcuate melanocortin neurons. Cell Metab. 2007, 5, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Adel, M.; Rabei, M.R.; Hazem, N.; Reda Hassan Elsayed, H.; El-Nablawy, M.H. Abscisic Acid Can Protect the Kidney Against Ischemia/Reperfusion Injury Via Antiapoptotic Activity, Downregulation of NOX-4 and Upregulation of Connexin-43. Bull. Egypt. Soc. Physiol. Sci. 2021, 41, 537–552. [Google Scholar] [CrossRef]
- Elsayed, H.R.H.; Anbar, H.S.; Rabei, M.R.; Adel, M.; El-Gamal, R. Eicosapentaenoic and docosahexaenoic acids attenuate methotrexate-induced apoptosis and suppression of splenic T, B-Lymphocytes and macrophages with modulation of expression of CD3, CD20 and CD68. Tissue Cell 2021, 72, 101533. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Gallaghar, S.R.; Smith, J.A. Current protocols in molecular biology, edited by M. Ausubel, R. Brent, R.E. Kingston, D.D. Moore, J.G. Seidman, J.A. Smith, and K. Struhl. Volumes 1 and 2. John Wiley & Sons, Inc., Media, PA, 1988, $165.00. Mol. Reprod. Dev. 1989, 1, 146. [Google Scholar] [CrossRef]
- Hames, B.D.; Rickwood, D. Gel Electrophoresis of Proteins: A Practical Approach, 2nd ed.; Oxford University Press: Oxford, UK, 1990; pp. 1–147. [Google Scholar]
- Bisen, P.S. Laboratory Protocols in Applied Life Sciences; Taylor & Francis: Abingdon, UK, 2014. [Google Scholar] [CrossRef]
- Glastras, S.J.; Chen, H.; Tsang, M.; Teh, R.; McGrath, R.T.; Zaky, A.; Chen, J.; Wong, M.G.; Pollock, C.A.; Saad, S. The renal consequences of maternal obesity in offspring are overwhelmed by postnatal high fat diet. PLoS ONE 2017, 12, e0172644. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Vara, J.; Miller, M. When tissue antigens and antibodies get along: Revisiting the technical aspects of immunohistochemistry—the red, brown, and blue technique. Vet. Pathol. 2014, 51, 42–87. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Wang, Z.; Proctor, G.; Moskowitz, S.; Liebman, S.E.; Rogers, T.; Lucia, M.S.; Li, J.; Levi, M. Diet-induced obesity in C57BL/6J mice causes increased renal lipid accumulation and glomerulosclerosis via a sterol regulatory element-binding protein-1c-dependent pathway. J. Biol. Chem. 2005, 280, 32317–32325. [Google Scholar] [CrossRef] [Green Version]
- Zoja, C.; Abbate, M.; Remuzzi, G. Progression of renal injury toward interstitial inflammation and glomerular sclerosis is dependent on abnormal protein filtration. Nephrol. Dial. Transplant. 2015, 30, 706–712. [Google Scholar] [CrossRef] [Green Version]
- Whaley-Connell, A.; Sowers, J.R. Obesity and kidney disease: From population to basic science and the search for new therapeutic targets. Kidney Int. 2017, 92, 313–323. [Google Scholar] [CrossRef]
- Nasri, R.; Abdelhedi, O.; Jemil, I.; Daoued, I.; Hamden, K.; Kallel, C.; Elfeki, A.; Lamri-Senhadji, M.; Boualga, A.; Nasri, M. Ameliorating effects of goby fish protein hydrolysates on high-fat-high-fructose diet-induced hyperglycemia, oxidative stress and deterioration of kidney function in rats. Chem.-Biol. Interact. 2015, 242, 71–80. [Google Scholar] [CrossRef]
- Li, C.; Culver, S.A.; Quadri, S.; Ledford, K.L.; Al-Share, Q.Y.; Ghadieh, H.E.; Najjar, S.M.; Siragy, H.M. High-fat diet amplifies renal renin angiotensin system expression, blood pressure elevation, and renal dysfunction caused by Ceacam1 null deletion. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E802–E810. [Google Scholar] [CrossRef] [Green Version]
- Hu, M.; Fan, M.; Zhen, J.; Lin, J.; Wang, Q.; Lv, Z.; Wang, R. FAK contributes to proteinuria in hypercholesterolaemic rats and modulates podocyte F-actin re-organization via activating p38 in response to ox-LDL. J. Cell. Mol. Med. 2017, 21, 552–567. [Google Scholar] [CrossRef]
- Demoulin, J.-B.; Ericsson, J.; Kallin, A.; Rorsman, C.; Rönnstrand, L.; Heldin, C.-H. Platelet-derived growth factor stimulates membrane lipid synthesis through activation of phosphatidylinositol 3-kinase and sterol regulatory element-binding proteins. J. Biol. Chem. 2004, 279, 35392–35402. [Google Scholar] [CrossRef] [Green Version]
- Elsayed, H.R.H.; El-Nablaway, M.; Khattab, B.A.; Sherif, R.N.; Elkashef, W.F.; Abdalla, A.M.; El Nashar, E.M.; Abd-Elmonem, M.M.; El-Gamal, R. Independent of Calorie Intake, Short-term Alternate-day Fasting Alleviates NASH, With Modulation of Markers of Lipogenesis, Autophagy, Apoptosis, and Inflammation in Rats. J. Histochem. Cytochem. 2021, 69, 575–596. [Google Scholar] [CrossRef]
- Sohn, M.; Kim, K.; Uddin, M.J.; Lee, G.; Hwang, I.; Kang, H.; Kim, H.; Lee, J.H.; Ha, H. Delayed treatment with fenofibrate protects against high-fat diet-induced kidney injury in mice: The possible role of AMPK autophagy. Am. J. Physiol. Ren. Physiol. 2017, 312, F323–F334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartleben, B.; Godel, M.; Meyer-Schwesinger, C.; Liu, S.; Ulrich, T.; Kobler, S.; Wiech, T.; Grahammer, F.; Arnold, S.J.; Lindenmeyer, M.T.; et al. Autophagy influences glomerular disease susceptibility and maintains podocyte homeostasis in aging mice. J. Clin. Investig. 2010, 120, 1084–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawakami, T.; Gomez, I.G.; Ren, S.; Hudkins, K.; Roach, A.; Alpers, C.E.; Shankland, S.J.; D’Agati, V.D.; Duffield, J.S. Deficient Autophagy Results in Mitochondrial Dysfunction and FSGS. J. Am. Soc. Nephrol. 2015, 26, 1040–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, F.F.; Horta, L.F.B.; de Albuquerque Maia, L.; Lopes-Pacheco, M.; da Silva, A.B.; Morales, M.M.; Gonçalves-de-Albuqueque, C.F.; Takiya, C.M.; de Castro-Faria-Neto, H.C.; Rocco, P.R.M. Dasatinib reduces lung inflammation and fibrosis in acute experimental silicosis. PLoS ONE 2016, 11, e0147005. [Google Scholar] [CrossRef]
- Mohammadalipour, A.; Karimi, J.; Khodadadi, I.; Solgi, G.; Hashemnia, M.; Sheikh, N.; Bahabadi, M. Dasatinib prevent hepatic fibrosis induced by carbon tetrachloride (CCl4) via anti-inflammatory and antioxidant mechanism. Immunopharmacol. Immunotoxicol. 2017, 39, 19–27. [Google Scholar] [CrossRef]
- Li, C.; Shen, Y.; Huang, L.; Liu, C.; Wang, J. Senolytic therapy ameliorates renal fibrosis postacute kidney injury by alleviating renal senescence. FASEB J. 2021, 35, e21229. [Google Scholar] [CrossRef]
- Islam, M.; Tuday, E.; Kim, J.; Dutson, T.; Phuong, T.; Donato, A.; Lesniewski, L. Senolytic Drugs, Dasatinib and Quercetin, Attenuate Adipose Tissue T Lymphocyte Infiltration and Improve Metabolic Function in Old Mice. FASEB J. 2021, 35. [Google Scholar] [CrossRef]
- Bouley, R.; Cheung, P.W.; Terlouw, A.; Brown, D. Dasatinib, a non-receptor tyrosine kinase inhibitor, induces aquaporin-2 membrane accumulation in a phospho-serine 256 independent manner. FASEB J. 2017, 31, 855.18. [Google Scholar] [CrossRef]
- Owens, W.A.; Walaszczyk, A.; Spyridopoulos, I.; Dookun, E.; Richardson, G.D. Senescence and senolytics in cardiovascular disease: Promise and potential pitfalls. Mech. Ageing Dev. 2021, 198, 111540. [Google Scholar] [CrossRef]
- Yi, J.S.; Perla, S.; Huang, Y.; Mizuno, K.; Giordano, F.J.; Vinks, A.A.; Bennett, A.M. Low-dose Dasatinib Ameliorates Hypertrophic Cardiomyopathy in Noonan Syndrome with Multiple Lentigines. Cardiovasc. Drugs Ther. 2021, 1–16. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Studied Parameter | Reference | Means | Standard Deviations | Effect Sizes | Sample Sizes |
|---|---|---|---|---|---|
| CD68 | [21] | 2, 11, 11 and 27 | 2 | 4.5052 | 8 |
| iNOS | [22] | 1.1, 2.2, 2.2, and 5.2 | 0.6 | 2.5424 | 8 |
| LC3 | [23] | 0.8, 1.3, 1.3, and 1.4 | 0.19 | 1.2343 | 16 |
| P62 | [23] | 0.22, 0.25, 0.25 and 0.31 | 0.025 | 1.3077 | 16 |
| SREBP | [24] | 0.4, 1, 1, and 1.2 | 0.08 | 3.75 | 8 |
| TGF-b | [25] | 1, 1.1, 1.1 and 1.5 | 0.15 | 1.2802 | 16 |
| ASMA | [26] | 8, 11, 17 and 26 | 2 | 3.4369 | 8 |
| Parameter | Group | F Value | p Value | |||
|---|---|---|---|---|---|---|
| Vehicle (n = 6) | Dasatinib Control (n = 6) | Obese (n = 6) | Obese + Dasatinib (n = 6) | |||
| Body weight (gm) | 34 ± 1.83 A | 33 ± 1.63 A | 41.25 ± 1.11 B | 37.25 ± 1.65 AB | 5.565 | 0.013 |
| Kidney weight (gm) | 0.24 ± 0.012 A | 0.24 ± 0.014 A | 0.32 ± 0.009 B | 0.27 ± 0.009 A | 10.114 | 0.001 |
| Parameter | Group | F Value | p Value | |||
|---|---|---|---|---|---|---|
| Vehicle (n = 6) | Dasatinib Control (n = 6) | Obese (n = 6) | Obese + Dasatinib (n = 6) | |||
| Glucose (mg/dL) | 83.25 ± 3.12 A | 85.50 ± 1.71 A | 121.50 ± 1.55 B | 96.00 ± 2.74 C | 44.780 | <0.0005 |
| Creatinine (mg/dL) | 0.088 ± 0.012 A | 0.082 ± 0.015 A | 0.188 ± 0.018 B | 0.098 ± 0.009 A | 13.058 | <0.0005 |
| Urea (mg/dL) | 32.98 ± 4.12 A | 38.05 ± 4.52 A | 72.54 ± 6.02 B | 56.10 ± 5.90 C | 12.032 | 0.001 |
| Parameter | Group | F Value | p Value | |||
|---|---|---|---|---|---|---|
| Vehicle (n = 6) | Dasatinib Control (n = 6) | Obese (n = 6) | Obese + Dasatinib (n = 6) | |||
| Total cholesterol (mg/dL) | 70.88 ± 6.99 A | 72.62 ± 3.15 A | 201.75 ± 8.19 B | 124.64 ± 4.66 C | 102.481 | <0.0005 |
| Triglyceride (mg/dL) | 90.34 ± 4.17 A | 93.92 ± 6.12 A | 167.23 ± 6.72 B | 130.22 ± 1.56 C | 50.705 | <0.0005 |
| LDL (mg/dL) | 32.76 ± 5.83 A | 34.58 ± 5.04 A | 127.53 ± 6.95 B | 65.27 ± 3.58 C | 65.032 | <0.0005 |
| HDL-C (mg/dL) | 27.45 ± 1.39 A | 27.98 ± 1.46 A | 42.98 ± 2.88 B | 39.82 ± 1.80 B | 16.491 | <0.0005 |
| Parameter | Group | F Value | p Value | |||
|---|---|---|---|---|---|---|
| Vehicle (n = 6) | Dasatinib Control (n = 6) | Obese (n = 6) | Obese + Dasatinib (n = 6) | |||
| Glomerulosclerosis | 0(0–1) A | 0(0–1) A | 4(2–4) B | 2(1–2) C | 50.523 | <0.0005 |
| Tubular dilatation | 0(0–1) A | 0(0–1) A | 3.5(2–4) B | 1.5(1–2) C | 51.044 | <0.0005 |
| Cast formation | 0(0–0.75) A | 0(0–1) A | 4(2.25–4) B | 1.5(1–2) C | 54.227 | <0.0005 |
| Tubular vacuolation | 0(0–1) A | 0(0–1) A | 4(2–4) B | 1.5(1–2) C | 46.877 | <0.0005 |
| Inflammatory cellular infiltrate | 0(0–1) A | 0(0–1) A | 3(2–3.75) B | 1(1–2) C | 48.078 | <0.0005 |
| Tubulointerstitial fibrosis | 0(0–1) A | 0(0–1) A | 3(2–4) B | 1.5(1–2) C | 52.222 | <0.0005 |
| Score | 3 (2–3.75) A | 3(2–4) A | 19.5(17–21) B | 9(7.25–10) C | 67.305 | <0.0005 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elsayed, H.R.H.; El-Gamal, R.; Rabei, M.R.; Elhadidy, M.G.; Hamed, S.; Othman, B.H.; Elshaer, M.M.A.; Sedky, M.K.; Hassan, A.T.A.E.; El-Nablaway, M. Enhanced Autophagic Flux, Suppressed Apoptosis and Reduced Macrophage Infiltration by Dasatinib in Kidneys of Obese Mice. Cells 2022, 11, 746. https://doi.org/10.3390/cells11040746
Elsayed HRH, El-Gamal R, Rabei MR, Elhadidy MG, Hamed S, Othman BH, Elshaer MMA, Sedky MK, Hassan ATAE, El-Nablaway M. Enhanced Autophagic Flux, Suppressed Apoptosis and Reduced Macrophage Infiltration by Dasatinib in Kidneys of Obese Mice. Cells. 2022; 11(4):746. https://doi.org/10.3390/cells11040746
Chicago/Turabian StyleElsayed, Hassan Reda Hassan, Randa El-Gamal, Mohammed R. Rabei, Mona G. Elhadidy, Shereen Hamed, Basma H. Othman, Mohamed Mahmoud Abdelraheem Elshaer, Mostafa Khaled Sedky, Ahmed Tarek Abd Elbaset Hassan, and Mohammad El-Nablaway. 2022. "Enhanced Autophagic Flux, Suppressed Apoptosis and Reduced Macrophage Infiltration by Dasatinib in Kidneys of Obese Mice" Cells 11, no. 4: 746. https://doi.org/10.3390/cells11040746