
Discovery of a Novel Aminocyclopropenone Compound That Inhibits BRD4-Driven Nucleoporin NUP210 Expression and Attenuates Colorectal Cancer Growth

 , , , , and
, , , , and 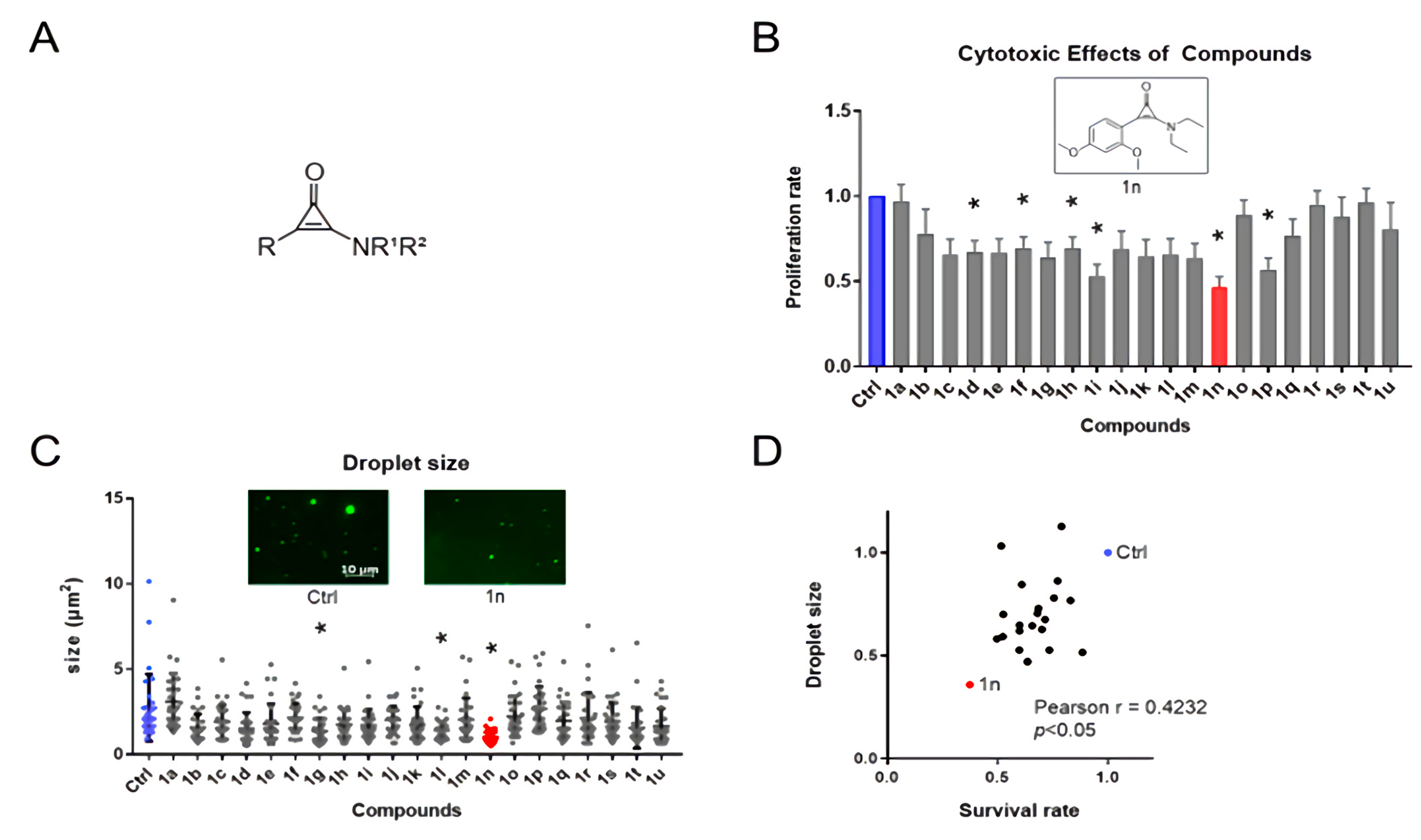{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Western Blotting
2.3. Cell Proliferation Assay
2.4. Colony Formation Assay
2.5. cDNA Preparation and Quantitative Real-Time RT-PCR Assay
2.6. Immunofluorescence Analysis
2.7. Transfections, Viral Particle Production, and Infection
2.8. Data Analysis for ChIP-Seq
2.9. Nuclear Size Measurement
2.10. In Vitro Droplet Assay
2.11. Statistical Analysis
3. Results
3.1. The Effects of Aminocyclopropenone Compounds on Colorectal Cancer Cell Growth and in Vitro Droplets of BRD4
3.2. Aminocyclopropenone Compound ACP-1n Prevented Super-Enhancer-Driven MYC Expression by Blocking BRD4 Assembly in the Nucleus
3.3. Identification of NUP210 as a BRD4-Driven Nuclear Complex Component
3.4. The Expression Levels of NUP210 Determine Cancer Cell Growth and Nucleus Size
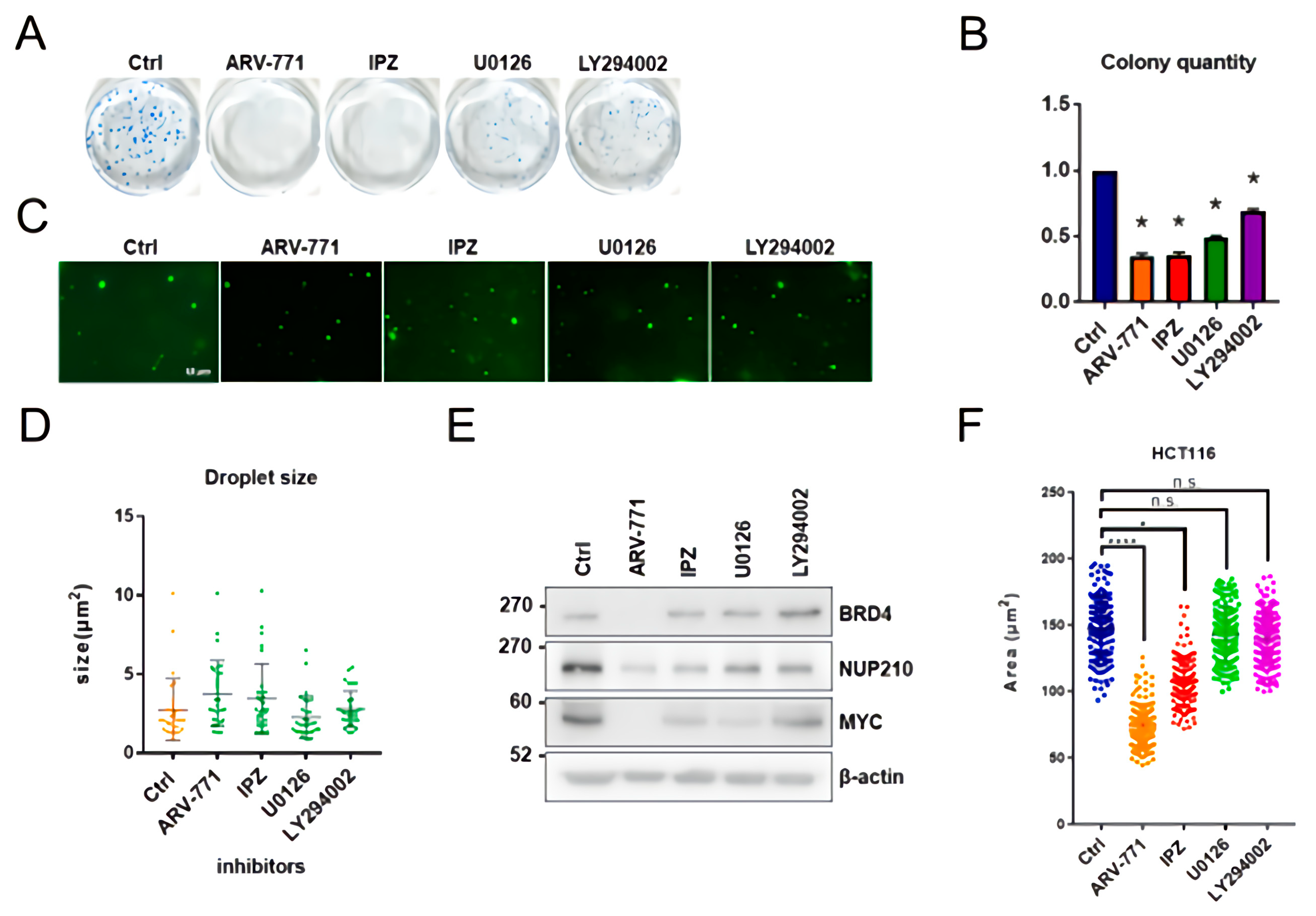
3.5. ARV-771, IPZ, U0126, and LY294002 Are Unable to Inhibit In Vitro BRD4 Assembly
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brenner, H.; Kloor, M.; Pox, C.P. Colorectal cancer. Lancet 2014, 383, 1490–1502. [Google Scholar] [CrossRef]
- Aghabozorgi, A.S.; Ebrahimi, R.; Bahiraee, A.; Tehrani, S.S.; Nabizadeh, F.; Setayesh, L.; Jafarzadeh-Esfehani, R.; Ferns, G.A.; Avan, A.; Rashidi, Z. The genetic factors associated with Wnt signaling pathway in colorectal cancer. Life Sci. 2020, 256, 118006. [Google Scholar] [CrossRef] [PubMed]
- Yi, J. DNA Methylation Change Profiling of Colorectal Disease: Screening towards Clinical Use. Life 2021, 11, 412. [Google Scholar] [CrossRef]
- Welch, H.G.; Robertson, D.J. Colorectal Cancer on the Decline—Why Screening Can’t Explain It All. N. Engl. J. Med. 2016, 374, 1605–1607. [Google Scholar] [CrossRef]
- Minamoto, T. Detection and Characterization of Oncogene Mutations in Preneoplastic and Early Neoplastic Lesions. Adv. Struct. Saf. Stud. 2014, 1105, 381–398. [Google Scholar] [CrossRef] [Green Version]
- Wong, R.W. Nuclear Pore Complex: From structural view to chemical tools. Cell Chem. Biol. 2015, 22, 1285–1287. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, M.S.; Hazawa, M.; Kobayashi, A.; Guillaud, L.; Watanabe-Nakayama, T.; Nakayama, M.; Wang, H.; Kodera, N.; Oshima, M.; Ando, T.; et al. Spatiotemporally tracking of nano-biofilaments inside the nuclear pore complex core. Biomaterials 2020, 256, 120198. [Google Scholar] [CrossRef]
- Mohamed, M.S.; Kobayashi, A.; Taoka, A.; Watanabe-Nakayama, T.; Kikuchi, Y.; Hazawa, M.; Minamoto, T.; Fukumori, Y.; Kodera, N.; Uchihashi, T.; et al. High-Speed Atomic Force Microscopy Reveals Loss of Nuclear Pore Resilience as a Dying Code in Colorectal Cancer Cells. ACS Nano 2017, 11, 5567–5578. [Google Scholar] [CrossRef]
- Dewi, F.R.; Domoto, T.; Hazawa, M.; Kobayashi, A.; Douwaki, T.; Minamoto, T.; Wong, R.W. Colorectal cancer cells require glycogen synthase kinase-3β for sustaining mitosis via translocated promoter region (TPR)-dynein interaction. Oncotarget 2018, 9, 13337–13352. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, A.; Hashizume, C.; Dowaki, T.; Wong, R.W. Therapeutic potential of mitotic interaction between the nucleoporin Tpr and aurora kinase A. Cell Cycle 2015, 14, 1447–1458. [Google Scholar] [CrossRef] [Green Version]
- Funasaka, T.; Tsuka, E.; Wong, R.W. Regulation of autophagy by nucleoporin Tpr. Sci. Rep. 2012, 2, 878. [Google Scholar] [CrossRef] [Green Version]
- Nakano, H.; Funasaka, T.; Hashizume, C.; Wong, R.W. Nucleoporin Translocated Promoter Region (Tpr) Associates with Dynein Complex, Preventing Chromosome Lagging Formation during Mitosis. J. Biol. Chem. 2010, 285, 10841–10849. [Google Scholar] [CrossRef] [Green Version]
- Hazawa, M.; Lin, D.; Kobayashi, A.; Jiang, Y.; Xu, L.; Dewi, F.R.P.; Mohamed, M.S.; Hartono; Nakada, M.; Meguro-Horike, M.; et al. ROCK-dependent phosphorylation of NUP 62 regulates p63 nuclear transport and squamous cell carcinoma proliferation. EMBO Rep. 2017, 19, 73–88. [Google Scholar] [CrossRef] [Green Version]
- Garcia, P.P.; Debo, B.; Aleman, J.R.; Talamas, J.A.; Lan, Y.; Nguyen, N.H.; Won, K.J.; Capelson, M. Metazoan Nuclear Pores Provide a Scaffold for Poised Genes and Mediate Induced Enhancer-Promoter Contacts. Mol. Cell 2017, 66, 63–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascual-Garcia, P.; Capelson, M. Nuclear pores in genome architecture and enhancer function. Curr. Opin. Cell Biol. 2019, 58, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Tan, H.; Wu, J.; He, G.; Liu, B. Dual-target inhibitors of bromodomain-containing protein 4 (BRD4) in cancer therapy: Current situation and future directions. Drug Discov. Today 2021, 27, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Perner, F.; Armstrong, S.A. Targeting Chromatin Complexes in Myeloid Malignancies and Beyond: From Basic Mechanisms to Clinical Innovation. Cells 2020, 9, 2721. [Google Scholar] [CrossRef]
- Shi, Y.; Liu, J.; Zhao, Y.; Cao, J.; Li, Y.; Guo, F. Bromodomain-Containing Protein 4: A Druggable Target. Curr. Drug Targets 2019, 20, 1517–1536. [Google Scholar] [CrossRef]
- Donati, B.; Lorenzini, E.; Ciarrocchi, A. BRD4 and Cancer: Going beyond transcriptional regulation. Mol. Cancer 2018, 17, 164. [Google Scholar] [CrossRef]
- Gen, Y.; Muramatsu, T.; Inoue, J.; Inazawa, J. miR-766-5p Targets Super-Enhancers by Downregulating CBP and BRD4. Cancer Res. 2021, 81, 5190–5201. [Google Scholar] [CrossRef]
- Shi, J.; Vakoc, C.R. The Mechanisms behind the Therapeutic Activity of BET Bromodomain Inhibition. Mol. Cell 2014, 54, 728–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.S.X.; Liu, C.L.C.; Liu, B.L.B.; Chen, J.C.J.; Wu, X.W.X.; Gong, W.G.J. JQ1: A novel potential therapeutic target. Pharmazie 2018, 73, 491–493. [Google Scholar] [PubMed]
- Raina, K.; Lu, J.; Qian, Y.; Altieri, M.; Gordon, D.; Rossi, A.M.K.; Wang, J.; Chen, X.; Dong, H.; Siu, K.; et al. PROTAC-induced BET protein degradation as a therapy for castration-resistant prostate cancer. Proc. Natl. Acad. Sci. USA 2016, 113, 7124–7129. [Google Scholar] [CrossRef] [Green Version]
- Lovén, J.; Hoke, H.A.; Lin, C.Y.; Lau, A.; Orlando, D.A.; Vakoc, C.R.; Bradner, J.E.; Lee, T.I.; Young, R.A. Selective Inhibition of Tumor Oncogenes by Disruption of Super-Enhancers. Cell 2013, 153, 320–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabari, B.R.; Dall’Agnese, A.; Boija, A.; Klein, I.A.; Coffey, E.L.; Shrinivas, K.; Abraham, B.J.; Hannett, N.M.; Zamudio, A.V.; Manteiga, J.C.; et al. Coactivator condensation at super-enhancers links phase separation and gene control. Science 2018, 361, 3958. [Google Scholar] [CrossRef] [Green Version]
- Hyman, A.; Weber, C.A.; Julicher, F. Liquid-Liquid Phase Separation in Biology. Annu. Rev. Cell Dev. Biol. 2014, 30, 39–58. [Google Scholar] [CrossRef] [Green Version]
- Tögel, L.; Nightingale, R.; Chueh, A.C.; Jayachandran, A.; Tran, H.; Phesse, T.; Wu, R.; Sieber, O.M.; Arango, D.; Dhillon, A.S.; et al. Dual Targeting of Bromodomain and Extraterminal Domain Proteins, and WNT or MAPK Signaling, Inhibits c-MYC Expression and Proliferation of Colorectal Cancer Cells. Mol. Cancer Ther. 2016, 15, 1217–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, R.M.; Huidobro, C.; Urdinguio, R.G.; Mangas, C.; Soldevilla, B.; Domínguez, G.; Bonilla, F.; Fernandez, A.F.; Fraga, M.F. Aberrant epigenetic regulation of bromodomain BRD4 in human colon cancer. J. Mol. Med. 2012, 90, 587–595. [Google Scholar] [CrossRef]
- McCleland, M.L.; Mesh, K.; Lorenzana, E.; Chopra, V.S.; Segal, E.; Watanabe, C.; Haley, B.; Mayba, O.; Yaylaoglu, M.; Gnad, F.; et al. CCAT1 is an enhancer-templated RNA that predicts BET sensitivity in colorectal cancer. J. Clin. Investig. 2016, 126, 639–652. [Google Scholar] [CrossRef]
- Hu, Y.; Zhou, J.; Ye, F.; Xiong, H.; Peng, L.; Zheng, Z.; Xu, F.; Cui, M.; Wei, C.; Wang, X.; et al. BRD4 Inhibitor Inhibits Colorectal Cancer Growth and Metastasis. Int. J. Mol. Sci. 2015, 16, 1928–1948. [Google Scholar] [CrossRef] [Green Version]
- Mishiro, K.; Yushima, Y.; Kunishima, M. Phototriggered Dehydration Condensation Using an Aminocyclopropenone. Org. Lett. 2017, 19, 4912–4915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishiro, K.; Nomura, M.; Furuyama, T.; Kunishima, M. Efficiency Enhancement of a Photocatalytic Decarbonylation of an Aminocyclopropenone by Benzothiophene Substitution. J. Org. Chem. 2021, 86, 3625–3636. [Google Scholar] [CrossRef] [PubMed]
- Dewi, F.R.P.; Jiapaer, S.; Kobayashi, A.; Hazawa, M.; Ikliptikawati, D.K.; Hartono; Sabit, H.; Nakada, M.; Wong, R.W. Nucleoporin TPR (translocated promoter region, nuclear basket protein) upregulation alters MTOR-HSF1 trails and suppresses autophagy induction in ependymoma. Autophagy 2021, 17, 1001–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, L.; Shao, N.; Liu, X.; Nestler, E. ngs.plot: Quick mining and visualization of next-generation sequencing data by integrating genomic databases. BMC Genom. 2014, 15, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazawa, M.; Amemori, S.; Nishiyama, Y.; Iga, Y.; Iwashima, Y.; Kobayashi, A.; Nagatani, H.; Mizuno, M.; Takahashi, K.; Wong, R.W. A light-switching pyrene probe to detect phase-separated biomolecules. iScience 2021, 24, 102865. [Google Scholar] [CrossRef]
- Ando, R.; Sakaki, T.; Morinaka, Y.; Takahashi, C.; Tamao, Y.; Yoshii, N.; Katayama, S.; Saito, K.-I.; Tokuyama, H.; Isaka, M.; et al. Cyclopropenone-containing cysteine proteinase inhibitors. Synthesis and enzyme inhibitory activities. Bioorg. Med. Chem. 1999, 7, 571–579. [Google Scholar] [CrossRef]
- Ando, R.; Morinaka, Y.; Tokuyama, H.; Isaka, M.; Nakamura, E. A new class of proteinase inhibitor. Cyclopropenone-containing inhibitor of papain. J. Am. Chem. Soc. 1993, 115, 1174–1175. [Google Scholar] [CrossRef]
- Kogen, H.; Kiho, T.; Tago, K.; Miyamoto, S.; Fujioka, T.; Otsuka, N.; Suzuki-Konagai, K.; Ogita, T. Alutacenoic Acids A and B, Rare Naturally Occurring Cyclopropenone Derivatives Isolated from Fungi: Potent Non-Peptide Factor XIIIa Inhibitors. J. Am. Chem. Soc. 2000, 122, 1842–1843. [Google Scholar] [CrossRef]
- Hazawa, M.; Sakai, K.; Kobayashi, A.; Yoshino, H.; Iga, Y.; Iwashima, Y.; Lim, K.S.; Voon, D.C.-C.; Jiang, Y.-Y.; Horike, S.-I.; et al. Disease-specific alteration of karyopherin-α subtype establishes feed-forward oncogenic signaling in head and neck squamous cell carcinoma. Oncogene 2020, 39, 2212–2223. [Google Scholar] [CrossRef] [Green Version]
- Hazawa, M.; Yoshino, H.; Nakagawa, Y.; Shimizume, R.; Nitta, K.; Sato, Y.; Sato, M.; Wong, R.W.; Kashiwakura, I. Karyopherin-β1 Regulates Radioresistance and Radiation-Increased Programmed Death-Ligand 1 Expression in Human Head and Neck Squamous Cell Carcinoma Cell Lines. Cancers 2020, 12, 908. [Google Scholar] [CrossRef] [Green Version]
- Chahine, M.N.; Pierce, G.N. Therapeutic Targeting of Nuclear Protein Import in Pathological Cell Conditions. Pharmacol. Rev. 2009, 61, 358–372. [Google Scholar] [CrossRef]
- Sun, B.; Fiskus, W.; Qian, Y.; Rajapakshe, K.; Raina, K.; Coleman, K.G.; Crew, A.P.; Shen, A.; Saenz, D.T.; Mill, C.P.; et al. BET protein proteolysis targeting chimera (PROTAC) exerts potent lethal activity against mantle cell lymphoma cells. Leukemia 2017, 32, 343–352. [Google Scholar] [CrossRef]
- Sun, J.; Shi, Y.; Yildirim, E. The Nuclear Pore Complex in Cell Type-Specific Chromatin Structure and Gene Regulation. Trends Genet. 2019, 35, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Jahangiri, L.; Tsaprouni, L.; Trigg, R.M.; Williams, J.A.; Gkoutos, G.V.; Turner, S.D.; Pereira, J. Core regulatory circuitries in defining cancer cell identity across the malignant spectrum. Open Biol. 2020, 10, 200121. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Yu, Y.; Ma, Z. Novel strategies targeting bromodomain-containing protein 4 (BRD4) for cancer drug discovery. Eur. J. Med. Chem. 2020, 200, 112426. [Google Scholar] [CrossRef] [PubMed]
- Mayer, S.V.; Murnauer, A.; Von Wrisberg, M.-K.; Jokisch, M.; Lang, K. Photo-induced and Rapid Labeling of Tetrazine-Bearing Proteins via Cyclopropenone-Caged Bicyclononynes. Angew. Chem. Int. Ed. 2019, 58, 15876–15882. [Google Scholar] [CrossRef] [Green Version]
- Jevtić, P.; Schibler, A.C.; Wesley, C.C.; Pegoraro, G.; Misteli, T.; Levy, D.L. The nucleoporin ELYS regulates nuclear size by controlling NPC number and nuclear import capacity. EMBO Rep. 2019, 20, e47283. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kondo, H.; Mishiro, K.; Iwashima, Y.; Qiu, Y.; Kobayashi, A.; Lim, K.; Domoto, T.; Minamoto, T.; Ogawa, K.; Kunishima, M.; et al. Discovery of a Novel Aminocyclopropenone Compound That Inhibits BRD4-Driven Nucleoporin NUP210 Expression and Attenuates Colorectal Cancer Growth. Cells 2022, 11, 317. https://doi.org/10.3390/cells11030317
Kondo H, Mishiro K, Iwashima Y, Qiu Y, Kobayashi A, Lim K, Domoto T, Minamoto T, Ogawa K, Kunishima M, et al. Discovery of a Novel Aminocyclopropenone Compound That Inhibits BRD4-Driven Nucleoporin NUP210 Expression and Attenuates Colorectal Cancer Growth. Cells. 2022; 11(3):317. https://doi.org/10.3390/cells11030317
Chicago/Turabian StyleKondo, Hiroya, Kenji Mishiro, Yuki Iwashima, Yujia Qiu, Akiko Kobayashi, Keesiang Lim, Takahiro Domoto, Toshinari Minamoto, Kazuma Ogawa, Munetaka Kunishima, and et al. 2022. "Discovery of a Novel Aminocyclopropenone Compound That Inhibits BRD4-Driven Nucleoporin NUP210 Expression and Attenuates Colorectal Cancer Growth" Cells 11, no. 3: 317. https://doi.org/10.3390/cells11030317