
Gain- and Loss-of-Function CFTR Alleles Are Associated with COVID-19 Clinical Outcomes
 , , , , , , , , , and
, , , , , , , , , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Patients
2.2. Whole Exome Sequencing Analysis (WES)
2.3. Chemicals, Vectors, and Antibodies
2.4. Cell Culture
2.5. Transient Transfection of the Variant CFTR Plasmids
2.6. YFP-Based Assay for the CFTR Activity
2.7. CFTR Half-Life Evaluation
2.8. Western Blot
2.9. Statistical Methods
3. Results
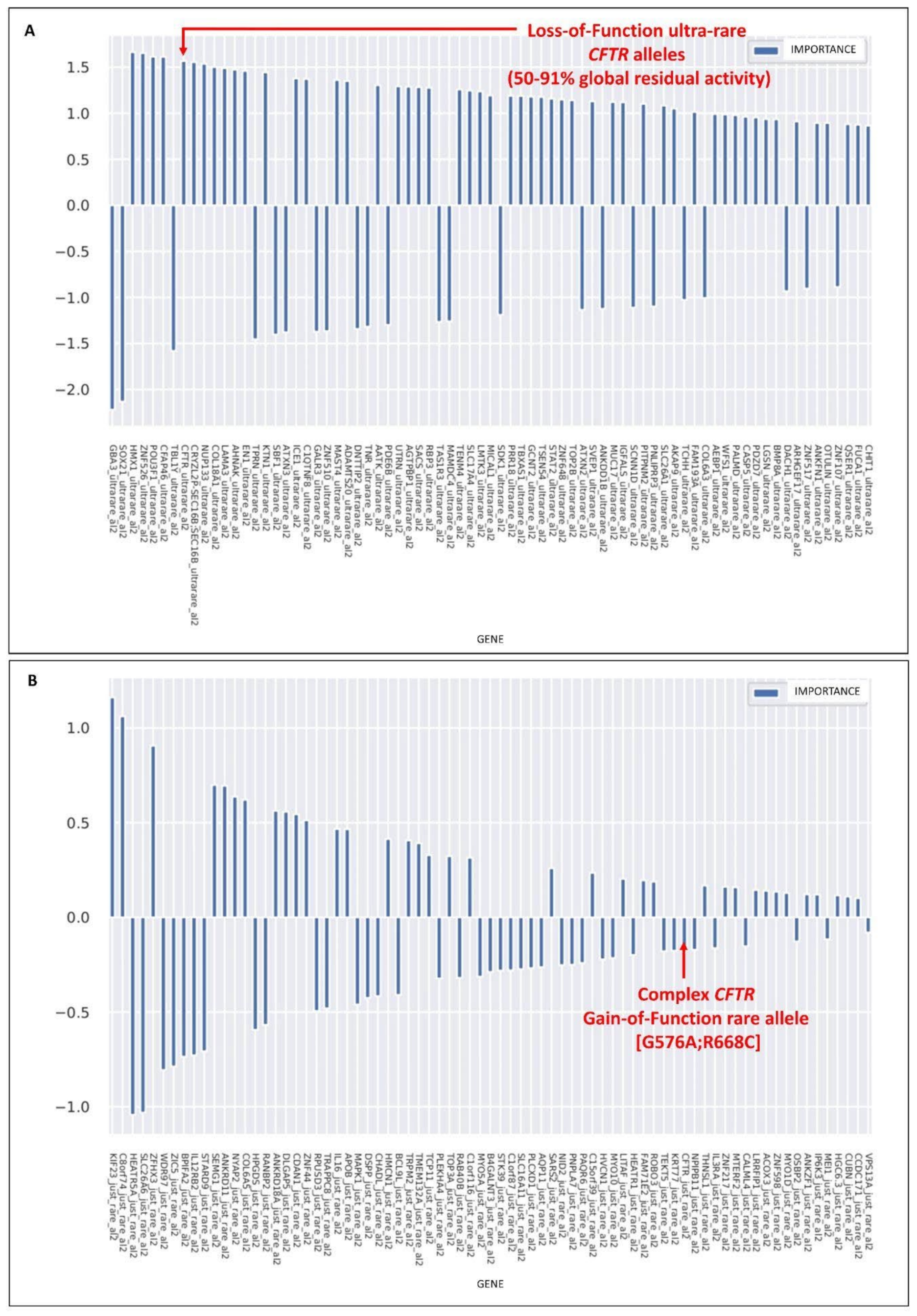
3.1. Rare CFTR Variants Contribute to Mildness While Ultra-Rare Variants Contribute to the Severity of COVID-19
3.2. The Rare CFTR [G576A;R558C] Complex Allele Is a Gain-of-Function Allele
3.3. Ultra-Rare Simple and Complex Alleles with Reduced CFTR Activity
3.4. Association of the CFTR Rare Gain-of-Function Complex Allele and Ultra-Rare Loss-of-Function Alleles with COVID-19 Clinical Outcomes
3.5. COVID-19 as a Mendelian Dominant/Recessive Disorder
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wiersinga, W.J.; Rhodes, A.; Cheng, A.C.; Peacock, S.J.; Prescott, H.C. Pathophysiology, Transmission, Diagnosis, and Treatment of Coronavirus Disease 2019 (COVID-19): A Review. JAMA 2020, 324, 782–793. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Chen, X.; Cai, Y.; Xia, J.; Zhou, X.; Xu, S.; Huang, H.; Zhang, L.; Zhou, X.; Du, C.; et al. Risk Factors Associated With Acute Respiratory Distress Syndrome and Death in Patients With Coronavirus Disease 2019 Pneumonia in Wuhan, China. JAMA Intern. Med. 2020, 180, 934–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grasselli, G.; Greco, M.; Zanella, A.; Albano, G.; Antonelli, M.; Bellani, G.; Bonanomi, E.; Cabrini, L.; Carlesso, E.; Castelli, G.; et al. Risk Factors Associated With Mortality Among Patients With COVID-19 in Intensive Care Units in Lombardy, Italy. JAMA Intern. Med. 2020, 180, 1345–1355. [Google Scholar] [CrossRef] [PubMed]
- Severe COVID-19 GWAS Group; Ellinghaus, D.; Degenhardt, F.; Bujanda, L.; Buti, M.; Albillos, A.; Invernizzi, P.; Fernández, J.; Prati, D.; Baselli, G.; et al. Genomewide Association Study of Severe COVID-19 with Respiratory Failure. N. Engl. J. Med. 2020, 383, 1522–1534. [Google Scholar] [CrossRef] [PubMed]
- Pairo-Castineira, E.; Clohisey, S.; Klaric, L.; Bretherick, A.D.; Rawlik, K.; Pasko, D.; Walker, S.; Parkinson, N.; Fourman, M.H.; Russell, C.D.; et al. Genetic mechanisms of critical illness in COVID-19. Nature 2021, 591, 92–98. [Google Scholar] [CrossRef]
- Kosmicki, J.A.; Horowitz, J.E.; Banerjee, N.; Lanche, R.; Marcketta, A.; Maxwell, E.; Bai, X.; Sun, D.; Backman, J.D.; Sharma, D.; et al. Pan-ancestry exome-wide association analyses of COVID-19 outcomes in 586,157 individuals. Am. J. Hum. Genet. 2021, 108, 1350–1355. [Google Scholar] [CrossRef]
- Ganna, A. COVID-19 Host Genetics Initiative. Mapping the human genetic architecture of COVID-19. Nature 2021, 600, 472–477. [Google Scholar] [CrossRef]
- Kousathanas, A.; Pairo-Castineira, E.; Rawlik, K.; Stuckey, A.; Odhams, C.A.; Walker, S.; Russell, C.D.; Malinauskas, T.; Wu, Y.; Millar, J.; et al. Whole-genome sequencing reveals host factors underlying critical COVID-19. Nature 2022, 607, 97–103. [Google Scholar] [CrossRef]
- Baldassarri, M.; Picchiotti, N.; Fava, F.; Fallerini, C.; Benetti, E.; Daga, S.; Valentino, F.; Doddato, G.; Furini, S.; Giliberti, A.; et al. Shorter androgen receptor polyQ alleles protect against life-threatening COVID-19 disease in European males. eBioMedicine 2021, 65, 103246. [Google Scholar] [CrossRef]
- Croci, S.; Venneri, M.A.; Mantovani, S.; Fallerini, C.; Benetti, E.; Picchiotti, N.; Campolo, F.; Imperatore, F.; Palmieri, M.; Daga, S.; et al. The polymorphism L412F in TLR3 inhibits autophagy and is a marker of severe COVID-19 in males. Autophagy 2021, 18, 1662–1672. [Google Scholar] [CrossRef]
- Fallerini, C.; Daga, S.; Benetti, E.; Picchiotti, N.; Zguro, K.; Catapano, F.; Baroni, V.; Lanini, S.; Bucalossi, A.; Marotta, G.; et al. SELP Asp603Asn and severe thrombosis in COVID-19 males. J. Hematol. Oncol. 2021, 14, 123. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Bastard, P.; Liu, Z.; Le Pen, J.; Moncada-Velez, M.; Chen, J.; Ogishi, M.; Sabli, I.K.D.; Hodeib, S.; Korol, C.; et al. Inborn errors of type I IFN immunity in patients with life-threatening COVID-19. Science 2020, 370, eabd4570. [Google Scholar] [CrossRef] [PubMed]
- Van Der Made, C.I.; Simons, A.; Schuurs-Hoeijmakers, J.; Heuvel, G.V.D.; Mantere, T.; Kersten, S.; Van Deuren, R.C.; Steehouwer, M.; Van Reijmersdal, S.V.; Jaeger, M.; et al. Presence of Genetic Variants Among Young Men With Severe COVID-19. JAMA 2020, 324, 663. [Google Scholar] [CrossRef] [PubMed]
- Fallerini, C.; Daga, S.; Mantovani, S.; Benetti, E.; Picchiotti, N.; Francisci, D.; Paciosi, F.; Schiaroli, E.; Baldassarri, M.; Fava, F.; et al. Association of Toll-like receptor 7 variants with life-threatening COVID-19 disease in males: Findings from a nested case-control study. eLife 2021, 10, e67569. [Google Scholar] [CrossRef]
- Solanich, X.; Vargas-Parra, G.; van der Made, C.I.; Simons, A.; Schuurs-Hoeijmakers, J.; Antolí, A.; del Valle, J.; Rocamora-Blanch, G.; Setién, F.; Esteller, M.; et al. Genetic Screening for TLR7 Variants in Young and Previously Healthy Men With Severe COVID-19. Front. Immunol. 2021, 12, 2965. [Google Scholar] [CrossRef]
- Mantovani, S.; Daga, S.; Fallerini, C.; Baldassarri, M.; Benetti, E.; Picchiotti, N.; Fava, F.; Gallì, A.; Zibellini, S.; Bruttini, M.; et al. Rare variants in Toll-like receptor 7 results in functional impairment and downregulation of cytokine-mediated signaling in COVID-19 patients. Genes Immun. 2021, 23, 51–56. [Google Scholar] [CrossRef]
- Baldassarri, M.; Fava, F.; Fallerini, C.; Daga, S.; Benetti, E.; Zguro, K.; Amitrano, S.; Valentino, F.; Doddato, G.; Giliberti, A.; et al. Severe COVID-19 in Hospitalized Carriers of Single CFTR Pathogenic Variants. J. Pers. Med. 2021, 11, 558. [Google Scholar] [CrossRef] [PubMed]
- Gabbi, C.; Renieri, A.; Strandvik, B. Geographical distribution of cystic fibrosis carriers as population genetic determinant of COVID-19 spread and fatality in 37 countries. J. Infect. 2022, 85, 318–321. [Google Scholar] [CrossRef]
- Picchiotti, N.; Benetti, E.; Fallerini, C.; Daga, S.; Baldassarri, M.; Fava, F.; Zguro, K.; Valentino, F.; Doddato, G.; Giliberti, A.; et al. Post-Mendelian genetic model in COVID-19. Cardiol. Cardiovasc. Med. 2021, 5, 673–694. [Google Scholar] [CrossRef]
- Fallerini, C.; Picchiotti, N.; Baldassarri, M.; Zguro, K.; Daga, S.; Fava, F.; Benetti, E.; Amitrano, S.; Bruttini, M.; Palmieri, M.; et al. Common, low-frequency, rare, and ultra-rare coding variants contribute to COVID-19 severity. Qual. Life Res. 2021, 141, 147–173. [Google Scholar] [CrossRef]
- Elborn, J.S. Cystic fibrosis. Lancet 2016, 388, 2519–2531. [Google Scholar] [CrossRef] [PubMed]
- Caohuy, H.; Eidelman, O.; Chen, T.; Yang, Q.; Walton, N.I.; Pollard, H.B. Inflammation in the COVID-19 airway is due to inhibition of CFTR signaling by the SARS-CoV-2 Spike protein. bioRxiv 2022. [CrossRef]
- Chen, L.; Guan, W.-J.; Qiu, Z.-E.; Xu, J.-B.; Bai, X.; Hou, X.-C.; Sun, J.; Qu, S.; Huang, Z.-X.; Lei, T.-L.; et al. SARS-CoV-2 nucleocapsid protein triggers hyperinflammation via protein-protein interaction-mediated intracellular Cl− accumulation in respiratory epithelium. Signal Transduct. Target. Ther. 2022, 7, 255. [Google Scholar] [CrossRef] [PubMed]
- Morales, L.; Oliveros, J.C.; Enjuanes, L.; Sola, I. Contribution of Host miRNA-223-3p to SARS-CoV-Induced Lung Inflammatory Pathology. mBio 2022, 13, e03135-21. [Google Scholar] [CrossRef]
- Daga, S.; Fallerini, C.; Baldassarri, M.; Fava, F.; Valentino, F.; Doddato, G.; Benetti, E.; Furini, S.; Giliberti, A.; Tita, R.; et al. Employing a systematic approach to biobanking and analyzing clinical and genetic data for advancing COVID-19 research. Eur. J. Hum. Genet. 2021, 29, 745–759. [Google Scholar] [CrossRef]
- COVID-19 Therapeutic Trial Synopsis. WHO R&D Blueprint Novel Coronavirus. COVID 19 Therapeutic Trial Synopsis. R&D Blueprint. 2020. Available online: https://www.who.int/publications/i/item/covid-19-therapeutic-trial-synopsis (accessed on 3 November 2022).
- Sondo, E.; Tomati, V.; Caci, E.; Esposito, A.I.; Pfeffer, U.; Pedemonte, N.; Galietta, L.J.V. Rescue of the mutant CFTR chloride channel by pharmacological correctors and low temperature analyzed by gene expression profiling. Am. J. Physiol. Physiol. 2011, 301, C872–C885. [Google Scholar] [CrossRef] [Green Version]
- Pedemonte, N.; Tomati, V.; Sondo, E.; Caci, E.; Millo, E.; Armirotti, A.; Damonte, G.; Zegarra-Moran, O.; Galietta, L.J.V. Dual Activity of Aminoarylthiazoles on the Trafficking and Gating Defects of the Cystic Fibrosis Transmembrane Conductance Regulator Chloride Channel Caused by Cystic Fibrosis Mutations. J. Biol. Chem. 2011, 286, 15215–15226. [Google Scholar] [CrossRef] [Green Version]
- Pedemonte, N.; Bertozzi, F.; Caci, E.; Sorana, F.; Di Fruscia, P.; Tomati, V.; Ferrera, L.; Rodríguez-Gimeno, A.; Berti, F.; Pesce, E.; et al. Discovery of a picomolar potency pharmacological corrector of the mutant CFTR chloride channel. Sci. Adv. 2020, 6, eaay9669. [Google Scholar] [CrossRef] [Green Version]
- Sondo, E.; Falchi, F.; Caci, E.; Ferrera, L.; Giacomini, E.; Pesce, E.; Tomati, V.; Bertozzi, S.M.; Goldoni, L.; Armirotti, A.; et al. Pharmacological Inhibition of the Ubiquitin Ligase RNF5 Rescues F508del-CFTR in Cystic Fibrosis Airway Epithelia. Cell Chem. Biol. 2018, 25, 891–905.e8. [Google Scholar] [CrossRef]
- Oliver, K.E.; Rauscher, R.; Mijnders, M.; Wang, W.; Wolpert, M.J.; Maya, J.; Sabusap, C.M.; Kesterson, R.A.; Kirk, K.L.; Rab, A.; et al. Slowing ribosome velocity restores folding and function of mutant CFTR. J. Clin. Investig. 2019, 129, 5236–5253. [Google Scholar] [CrossRef]
- Veit, G.; Oliver, K.; Apaja, P.M.; Perdomo, D.; Bidaud-Meynard, A.; Lin, S.-T.; Guo, J.; Icyuz, M.; Sorscher, E.J.; Iv, J.L.H.; et al. Ribosomal Stalk Protein Silencing Partially Corrects the ΔF508-CFTR Functional Expression Defect. PLoS Biol. 2016, 14, e1002462, Erratum in PLoS Biol. 2016, 14, e1002574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meriin, A.; Mense, M.; Colbert, J.D.; Liang, F.; Bihler, H.; Zaarur, N.; Rock, K.L.; Sherman, M.Y. A Novel Approach to Recovery of Function of Mutant Proteins by Slowing Down Translation. J. Biol. Chem. 2012, 287, 34264–34272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.K.; Schultz, B.D.; Katzenellenbogen, J.A.; Price, E.M.; Bridges, R.J.; Bradbury, N.A. Estrogen inhibition of cystic fibrosis transmembrane conductance regulator-mediated chloride secretion. J. Pharmacol. Exp. Ther. 2000, 295, 195–204. [Google Scholar]
- Ajonuma, L.C.; Tsang, L.L.; Zhang, G.H.; Wong, C.H.Y.; Lau, M.C.; Ho, L.S.; Rowlands, D.K.; Zhou, C.X.; Ng, C.P.; Chen, J.; et al. Estrogen-Induced Abnormally High Cystic Fibrosis Transmembrane Conductance Regulator Expression Results in Ovarian Hyperstimulation Syndrome. Mol. Endocrinol. 2005, 19, 3038–3044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saint-Criq, V.; Kim, S.H.; Katzenellenbogen, J.A.; Harvey, B.J. Non-Genomic Estrogen Regulation of Ion Transport and Airway Surface Liquid Dynamics in Cystic Fibrosis Bronchial Epithelium. PLoS ONE 2013, 8, e78593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rochwerger, L.; Dho, S.; Parker, L.; Foskett, J.K.; Buchwald, M. Estrogen-dependent expression of the cystic fibrosis transmembrane regulator gene in a novel uterine epithelial cell line. J. Cell Sci. 1994, 107, 2439–2448. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, A.; Kaufman, J.M.; Goemaere, S.; van Pottelberg, I. Estradiol in elderly men. Aging Male 2002, 5, 98–102. [Google Scholar] [CrossRef]
- Schmidt, A.; Peters, S.; Knaus, A.; Sabir, H.; Hamsen, F.; Maj, C.; Fazaal, J.; Sivalingam, S.; Savchenko, O.; Mantri, A.; et al. TBK1 and TNFRSF13B mutations and an autoinflammatory disease in a child with lethal COVID-19. NPJ Genom. Med. 2021, 6, 55. [Google Scholar] [CrossRef] [PubMed]
- Rivera, E.G.; Patnaik, A.; Salvemini, J.; Jain, S.; Lee, K.; Lozeau, D.; Yao, Q. SARS-CoV-2/COVID-19 and its relationship with NOD2 and ubiquitination. Clin. Immunol. 2022, 238, 109027. [Google Scholar] [CrossRef]
- Masood, K.I.; Yameen, M.; Ashraf, J.; Shahid, S.; Mahmood, S.F.; Nasir, A.; Nasir, N.; Jamil, B.; Ghanchi, N.K.; Khanum, I.; et al. Upregulated type I interferon responses in asymptomatic COVID-19 infection are associated with improved clinical outcome. Sci. Rep. 2021, 11, 22958. [Google Scholar] [CrossRef]
- Malaquias, M.A.S.; Gadotti, A.C.; Motta-Junior, J.D.S.; Martins, A.P.C.; Azevedo, M.L.V.; Benevides, A.P.K.; Cézar-Neto, P.; Carmo, L.A.P.D.; Zeni, R.C.; Raboni, S.M.; et al. The role of the lectin pathway of the complement system in SARS-CoV-2 lung injury. Transl. Res. J. Lab. Clin. Med. 2020, 231, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Sahebnasagh, A.; Nabavi, S.M.; Kashani, H.R.K.; Abdollahian, S.; Habtemariam, S.; Rezabakhsh, A. Anti-VEGF agents: As appealing targets in the setting of COVID-19 treatment in critically ill patients. Int. Immunopharmacol. 2021, 101, 108257. [Google Scholar] [CrossRef] [PubMed]
- Kuek, L.E.; Lee, R.J. First contact: The role of respiratory cilia in host-pathogen interactions in the airways. Am. J. Physiol. Cell. Mol. Physiol. 2020, 319, L603–L619. [Google Scholar] [CrossRef] [PubMed]
- Coté, J.J.; Granger, P.; Mishra, A.; Sorini, G. COVID-19 in a pregnant cystic fibrosis carrier with myasthenia gravis: A case report. Case Rep. Women’s Health 2022, 34, e00406. [Google Scholar] [CrossRef] [PubMed]
- Wellmerling, J.H.; Chang, S.-W.; Kim, E.; Osman, W.H.; Boyaka, P.; Borchers, M.T.; Cormet-Boyaka, E. Reduced expression of the Ion channel CFTR contributes to airspace enlargement as a consequence of aging and in response to cigarette smoke in mice. Respir. Res. 2019, 20, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiemer, L.; Lund, O.; Brunak, S.; Blom, N.S. Coronavirus 3CLpro proteinase cleavage sites: Possible relevance to SARS virus pathology. BMC Bioinform. 2004, 5, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabriel, S.E.; Brigman, K.N.; Koller, B.H.; Boucher, R.C.; Stutts, M.J. Cystic Fibrosis Heterozygote Resistance to Cholera Toxin in the Cystic Fibrosis Mouse Model. Science 1994, 266, 107–109. [Google Scholar] [CrossRef]
- Trapnell, B.C.; Chu, C.S.; Paakko, P.K.; Banks, T.C.; Yoshimura, K.; Ferrans, V.J.; Chernick, M.S.; Crystal, R.G. Expression of the cystic fibrosis transmembrane conductance regulator gene in the respiratory tract of normal individuals and individuals with cystic fibrosis. Proc. Natl. Acad. Sci. USA 1991, 88, 6565–6569. [Google Scholar] [CrossRef] [Green Version]
- Mathew, H.R.; Choi, M.Y.; Parkins, M.D.; Fritzler, M.J. Systematic review: Cystic fibrosis in the SARS-CoV-2/COVID-19 pandemic. BMC Pulm. Med. 2021, 21, 173. [Google Scholar] [CrossRef]
- Carr, S.B.; McClenaghan, E.; Elbert, A.; Faro, A.; Cosgriff, R.; Abdrakhmanov, O.; Brownlee, K.; Burgel, P.-R.; Byrnes, C.A.; Cheng, S.Y.; et al. Factors associated with clinical progression to severe COVID-19 in people with cystic fibrosis: A global observational study. J. Cyst. Fibros. Off. J. Eur. Cyst. Fibros. Soc. 2021, 21, e221–e231. [Google Scholar] [CrossRef]
- Bezzerri, V.; Gentili, V.; Api, M.; Finotti, A.; Papi, C.; Tamanini, A.; Olioso, D.; Duca, M.; Tedesco, E.; Leo, S.; et al. ACE2 expression and localization are regulated by CFTR: Implications beyond cystic fibrosis. bioRxiv 2021. [Google Scholar] [CrossRef]
- Lader, A.S.; Prat, A.G.; Jackson, G.R.; Chervinsky, K.L.; Lapey, A.; Kinane, T.B.; Cantiello, H.F. Increased circulating levels of plasma ATP in cystic fibrosis patients. Clin. Physiol. 2000, 20, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Abraham, E.H.; Guidotti, G.; Rapaport, E.; Bower, D.; Brown, J.; Griffin, R.J.; Donnelly, A.; Waitzkin, E.D.; Qamar, K.; Thompson, M.A.; et al. Cystic fibrosis improves COVID-19 survival and provides clues for treatment of SARS-CoV-2. Purinergic Signal. 2021, 17, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Abraham, E.H.; Vos, P.; Kahn, J.; Grubman, S.A.; Jefferson, D.M.; Ding, I.; Okunieff, P. Cystic fibrosis hetero–and homozygosity is associated with inhibition of breast cancer growth. Nat. Med. 1996, 2, 593–596. [Google Scholar] [CrossRef]
- Becq, F. CFTR channels and adenosine triphosphate release: The impossible rendez-vous revisited in skeletal muscle. J. Physiol. 2010, 588, 4605–4606. [Google Scholar] [CrossRef]
- Caracciolo, M.; Correale, P.; Mangano, C.; Foti, G.; Falcone, C.; Macheda, S.; Cuzzola, M.; Conte, M.; Falzea, A.C.; Iuliano, E.; et al. Efficacy and Effect of Inhaled Adenosine Treatment in Hospitalized COVID-19 Patients. Front. Immunol. 2021, 12, 613070. [Google Scholar] [CrossRef]
- Correale, P.; Caracciolo, M.; Bilotta, F.; Conte, M.; Cuzzola, M.; Falcone, C.; Mangano, C.; Falzea, A.C.; Iuliano, E.; Morabito, A.; et al. Therapeutic effects of adenosine in high flow 21% oxygen aereosol in patients with COVID-19-pneumonia. PLoS ONE 2020, 15, e0239692. [Google Scholar] [CrossRef]
- Falcone, C.; Caracciolo, M.; Correale, P.; Macheda, S.; Vadalà, E.; La Scala, S.; Tescione, M.; Danieli, R.; Ferrarelli, A.; Tarsitano, M.; et al. Can Adenosine Fight COVID-19 Acute Respiratory Distress Syndrome? J. Clin. Med. 2020, 9, 3045. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
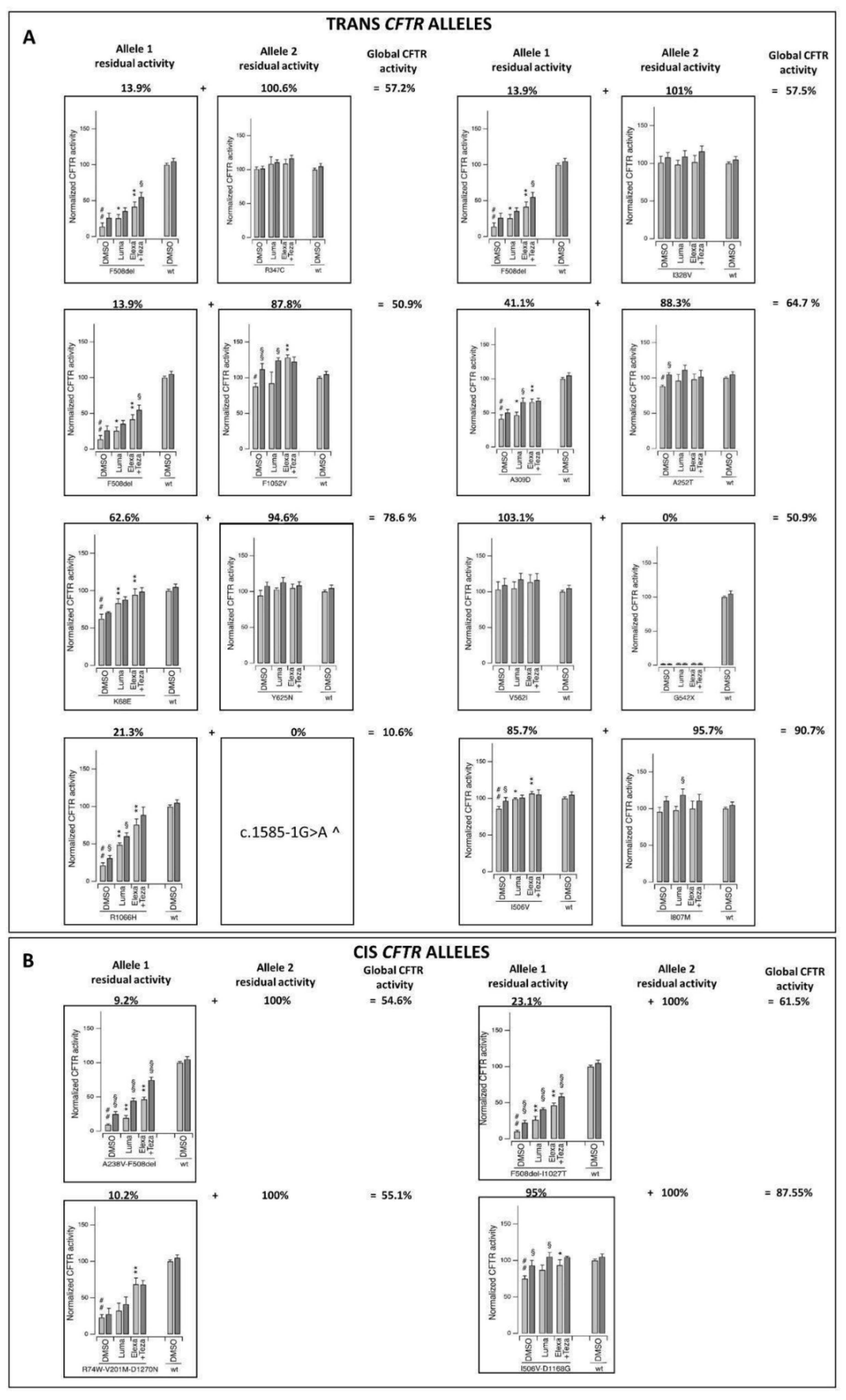
| Nucleotide Change | Amino Acid Change | dbSNP | CADD | gnomAD_ NFE | Cis/ Trans | Alleles’ Residual Activity (%) | Global Residual Activity (%) | N. of Patients | Clinical Category | Age | Sex | Long COVID (Yes/No) | Patient ID | IPGS |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| c.1039C>T | p.R347C | rs39750814 | 34 | 0.0000088 | trans | 100.6 | 57.2 | 1 | 2 | 32 | M | Yes | P1 | 180 |
| c.1520_1522del | p.F508del | rs113993960 | n.a. | 0.012 | 13.9 | |||||||||
| c.[713C>T;1520_1522del] | p.[A238V; F508del] | rs39750878; rs113993960 | 26; n.a. | n.a; 0.012 | cis | 9.2 | 54.6 | 1 | 2 | 30 | M | Yes | P2 | 250 |
| WT | WT | n.a. | n.a. | n.a. | 100 ^ | |||||||||
| c.3154T>G | p.F1052V | rs15021278 | 31 | 0.009 | trans | 87.8 | 50.9 | 1 | 4 | 52 | M | Yes | P3 | −80 |
| c.1520_1522del | p.F508del | rs113993960 | n.a. | 0.012 | 13.9 | |||||||||
| c.202A>G | p.K68E | rs39750833 | 24 | 0.0001 | trans | 62.6 | 78.6 | 1 | 4 | 42 | M | n.a. | P4 | 32 |
| c.1873T>A | p.Y625N | rs760390633 | 24 | 0.000018 | 94.6 | |||||||||
| c.A982A>G | p.I328V | n.a. | 29 | n.a. | trans | 101 | 57.5 | 1 | 3 | 63 | M | Yes | P5 | 215 |
| c.1520_1522del | p.F508del | rs113993960 | n.a. | 0.012 | 13.9 | |||||||||
| 215c.754G>A | p.A252T | n.a. | 24 | n.a. | trans | 88.3 | 64.7 | 1 | 4 | 91 | F | Yes | P6 | −174 |
| c.926C>A | p.A309D | n.a. | 32 | n.a. | 41.1 | |||||||||
| c.1624G>T | p.G542 * | rs11399395 | 39 | 0.0004 | trans | 0 | 51.55 | 1 | 3 | 77 | F | n.a. | P7 ° | 176 |
| c.1684G>A | p.V562I | rs1800097 | 24 | 0.0002 | 103.1 | |||||||||
| c.3197G>A | p.R1066H | rs121909019 | 34 | 0.000076 | trans | 21.3 | 10.6 | 1 | 4 | 51 | M | Yes | P8 | −15 |
| c.1585-1G>A | n.a. | rs76713772 | 26 | 0.0001 | 0 | |||||||||
| c.[1516A>G;3503A>G] | p.[I506V;D1168G] | rs1800091; rs150326506 | 24; 28 | 0.0005; na | cis | 95 | 87.55 | 2 | 1 | 65 | M | No | P9 | 243 |
| WT | WT | n.a. | n.a. | n.a. | 100 ^ | 2 | 69 | M | n.a. | P10 | 276 |
| Nucleotide Change | Amino Acid Change | dbSNP | CADD | gnomAD_ NFE | Cis/ Trans | Residual Allele Activity (%) | Global Residual Activity (%) | N. of Patients | Clinical Category | Age | Sex | Long-COVID (Yes/No) | Patient ID | IPGS |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Complex Allele 1 * | Complex Allele 1 * | rs115545701; rs11971167; rs138338446 | 24; 34; 27 | 0.0003; 0.0001; 0.0002 | cis | 23.1 | 61.6 | 1 | 6 | 38 | M | NA | P11 | −170 |
| WT | WT | n.a. | n.a. | n.a. | 100 ^ | |||||||||
| Complex allele 2 # | Complex allele 2 # | rs1800112; rs113993960 | 17; na | 0.003; 0.0123 | cis | 10.2 | 55.1 | 1 | 6 | 21 | F | Yes | P12 | −22 |
| WT | WT | n.a. | n.a. | n.a | 100 ^ | |||||||||
| c.1516A>G | p.I506V | rs1800091 | 24 | 0.0005 | trans | 85.7 | 90.7 | 1 | 6 | 43 | F | NA | P13 | −32 |
| c.2421A>G | p.I807M | rs1800103 | 22 | 0.0006 | 95.7 |
| Clinical Category | Females (n.) | Median Age (IQR) | Total n. Patients |
|---|---|---|---|
| Cat. 1 | F (1) | 84 y | 1 |
| Cat. 2 | F (1) | 57 y | 2 |
| M (1) | 75 y | ||
| Cat. 3 | 6(F) | 81.5 y (69.3–85.5) | 15 |
| 9 (M) | 71 y (56.3–76.5) | ||
| Cat. 4 | 2 (F) | 77;82 y | 11 |
| 9(M) | 62 y (54–72.5) | ||
| Cat. 5 | 2 (F) | 43;53 y | 4 |
| 2(M) | 57;64 y | ||
| Cat. 6 | 2 (F) | 44;45 y | 8 |
| 6 (M) | 57.5 y (42.5–63.5) |
| Female | Male | ||||
|---|---|---|---|---|---|
| Total (n = 2585) | Age < 50 y (n = 289) | Age ≥ 50 y (n = 765) | Age < 50 y (n = 317) | Age ≥ 50 y (n = 1214) | |
| COVID Outcome Scale, No. (%) | |||||
| Cat. 1 (Death) | 164 (6.34) | 2 (0.69) | 68 (8.89) ** | 1 (0.32) | 93 (7.66) ** |
| Cat. 2 (Invasive mechanical ventilation) | 180 (6.96) | 6 (2.08) | 36 (4.71) * | 23 (7.26) ° | 115 (9.47) ° |
| Cat. 3 (CPAP/BiPAP) | 584 (22.59) | 24 (8.30) | 163 (21.31) ** | 55 (17.35) ° | 342 (28.17) ** |
| Cat. 4 (Hospitalized, receiving supplemental low-flow oxygen) | 850 (32.88) | 37 (12.8) | 291 (38.04) ** | 79 (24.92) ° | 443 (36.49) ** |
| Cat. 5 (Hospitalized, not receiving supplemental oxygen) | 347 (13.42) | 58 (20.07) | 96 (12.55) * | 63 (19.87) | 130 (10.71) ** |
| Cat. 6 (Not hospitalized) | 460 (17.79) | 162 (56.6) | 111 (14.49) ** | 96 (30.28) ° | 91 (7.5) **,° |
| Female | Male | ||||
|---|---|---|---|---|---|
| Total | Age < 50 y (n = 289) | Age ≥ 50 y (n = 765) | Age < 50 y (n = 317) | Age ≥ 50 y (n = 1214) | |
| Individuals | 41 | 3 (1.04) | 11 (1.44) | 4 (1.26) | 23 (1.89) |
| COVID Outcome Scale, No. (%) | |||||
| Cat. 1 (Death; n = 164) | 1 (0.61) | 0 | 1 (1.47) | 0 | 0 |
| Cat. 2 (Invasive mechanical ventilation; n = 180) | 2 (1.11) | 0 | 1 (2.78) | 0 | 1 (0.87) |
| Cat. 3 (CPAP/BiPAP; n = 584) | 15 (2.57) | 0 | 6 (3.68) ° | 1 (1.82) | 8 (2.34) |
| Cat. 4 (Hospitalized, receiving supplemental low-flow oxygen; n = 850) | 11 (1.29) | 0 | 2 (0.69) | 1 (1.27) | 8 (1.81) |
| Cat. 5 (Hospitalized, not receiving supplemental oxygen; n = 347) | 4 (1.15) | 1 (1.72) | 1 (1.04) | 0 | 2 (1.54) |
| Cat. 6 (Not hospitalized; n = 460) | 8 (1.74) | 2 (1.23) | 0 | 2 (2.08) | 4 (4.4) * |
| Female | Male | ||||
|---|---|---|---|---|---|
| Total (n = 2585) | Age < 50 y (n = 289) | Age ≥ 50 y (n = 765) | Age < 50 y (n = 317) | Age ≥ 50 y (n = 1214) | |
| Individuals | 114 | (n = 13) | (n = 25) | (n = 20) | (n = 56) |
| COVID Outcome Scale, No. (%) | |||||
| Cat. 1 (Death; n = 164) | 13 (7.93) *,° | 0 | 4 (5.88) | 0 | 9 (9.68) * |
| Cat. 2 (Invasive mechanical ventilation; n = 180) | 10 (5.56) | 0 | 3 (8.33) * | 3 (13.04) | 4 (3.48) |
| Cat. 3 (CPAP/BiPAP; n = 584) | 27 (4.62) | 1 (4.17) | 7 (4.29) | 3 (5.45) | 16 (4.68) |
| Cat. 4 (Hospitalized, receiving supplemental low-flow oxygen; n = 850) | 29 (3.41) | 2 (5.41) | 4 (1.37) | 5 (6.33) | 18 (4.06) |
| Cat. 5 (Hospitalized, not receiving supplemental oxygen; n = 347) | 13 (3.75) | 2 (3.45) | 3 (3.12) | 2 (3.17) | 6 (4.62) |
| Cat. 6 (Not hospitalized; n = 460) | 22 (4.78) | 8 (4.94) | 4 (3.6) | 7 (7.29) | 3 (3.30) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldassarri, M.; Zguro, K.; Tomati, V.; Pastorino, C.; Fava, F.; Croci, S.; Bruttini, M.; Picchiotti, N.; Furini, S.; GEN-COVID Multicenter Study; et al. Gain- and Loss-of-Function CFTR Alleles Are Associated with COVID-19 Clinical Outcomes. Cells 2022, 11, 4096. https://doi.org/10.3390/cells11244096
Baldassarri M, Zguro K, Tomati V, Pastorino C, Fava F, Croci S, Bruttini M, Picchiotti N, Furini S, GEN-COVID Multicenter Study, et al. Gain- and Loss-of-Function CFTR Alleles Are Associated with COVID-19 Clinical Outcomes. Cells. 2022; 11(24):4096. https://doi.org/10.3390/cells11244096
Chicago/Turabian StyleBaldassarri, Margherita, Kristina Zguro, Valeria Tomati, Cristina Pastorino, Francesca Fava, Susanna Croci, Mirella Bruttini, Nicola Picchiotti, Simone Furini, GEN-COVID Multicenter Study, and et al. 2022. "Gain- and Loss-of-Function CFTR Alleles Are Associated with COVID-19 Clinical Outcomes" Cells 11, no. 24: 4096. https://doi.org/10.3390/cells11244096