
Comparison of the Single-Cell Immune Landscape of Testudines from Different Habitats
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
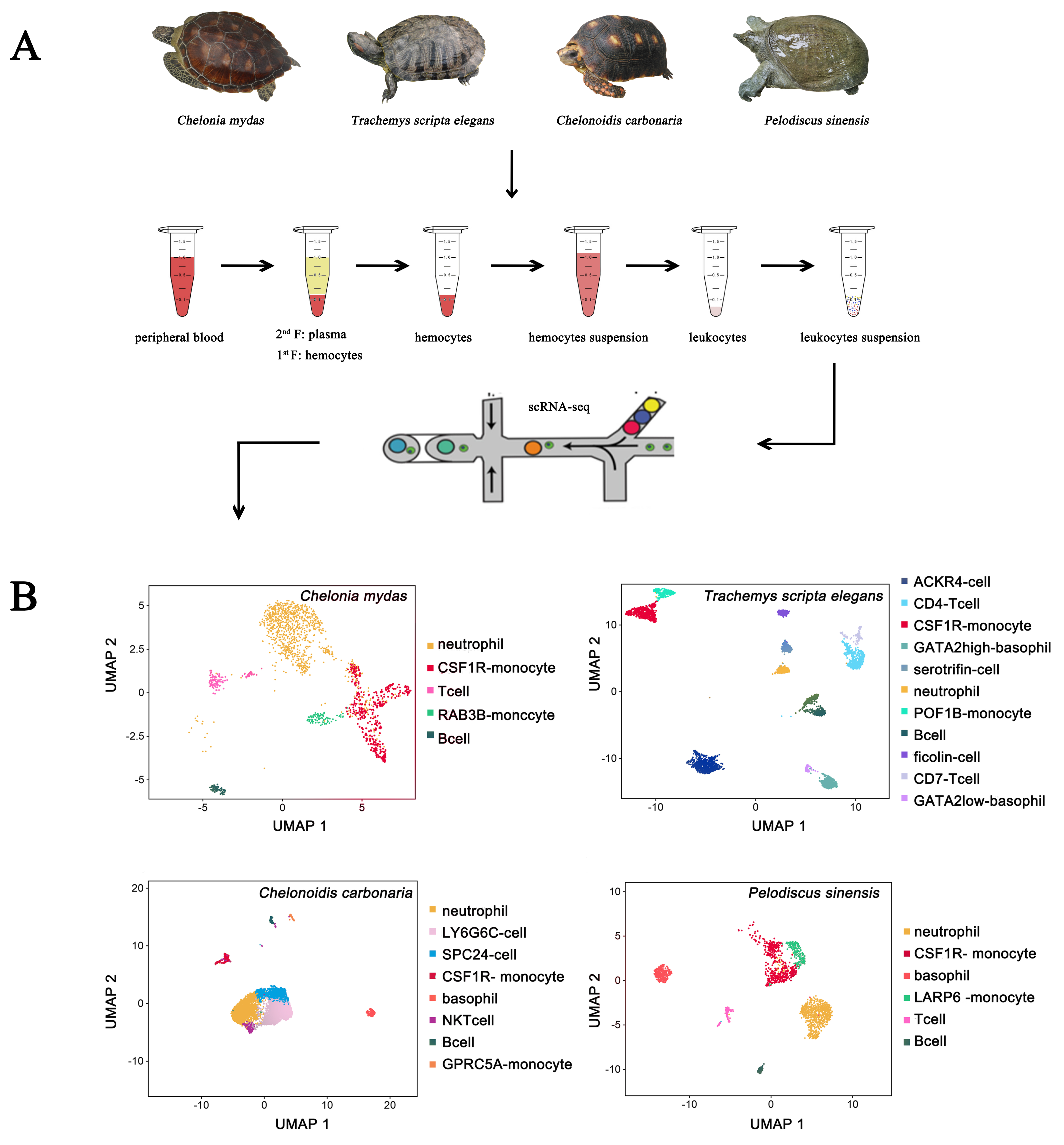
2.1. Sample Collection
2.2. Preparation of Single Cell Suspension
2.3. ScRNA-seq Data Processing and Analysis
3. Results
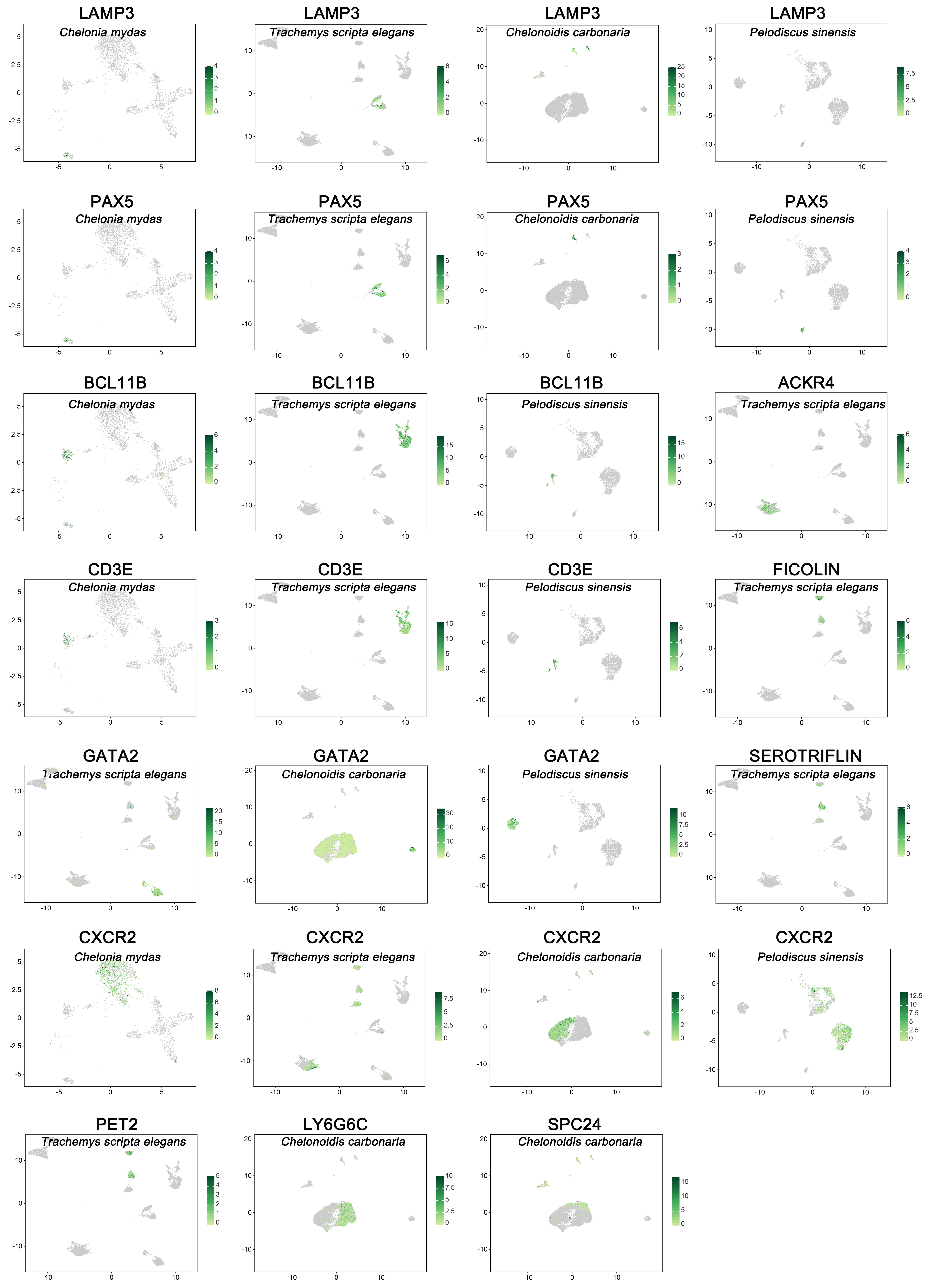
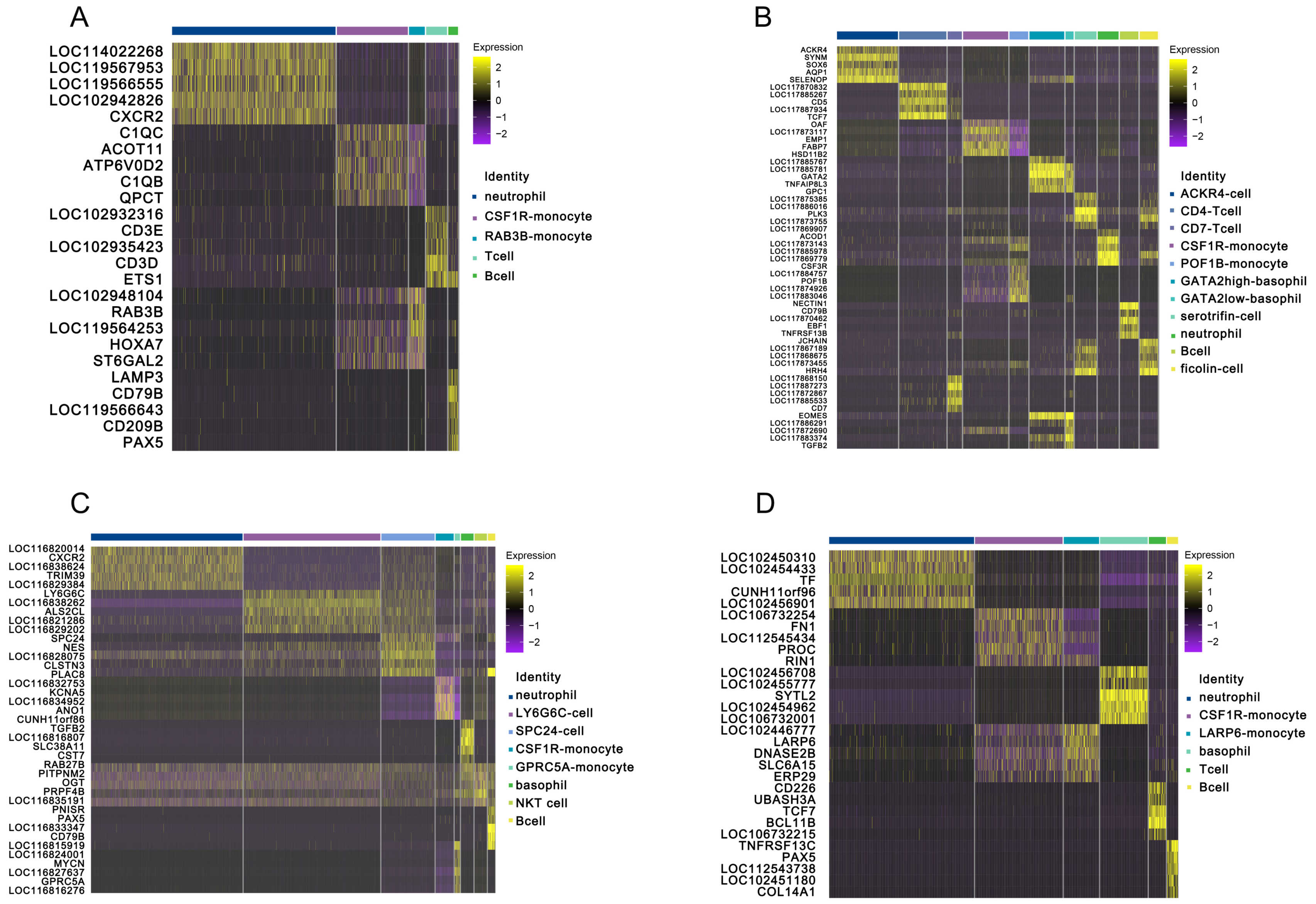
3.1. Landscapes of Peripheral Immune Cells in Testudines
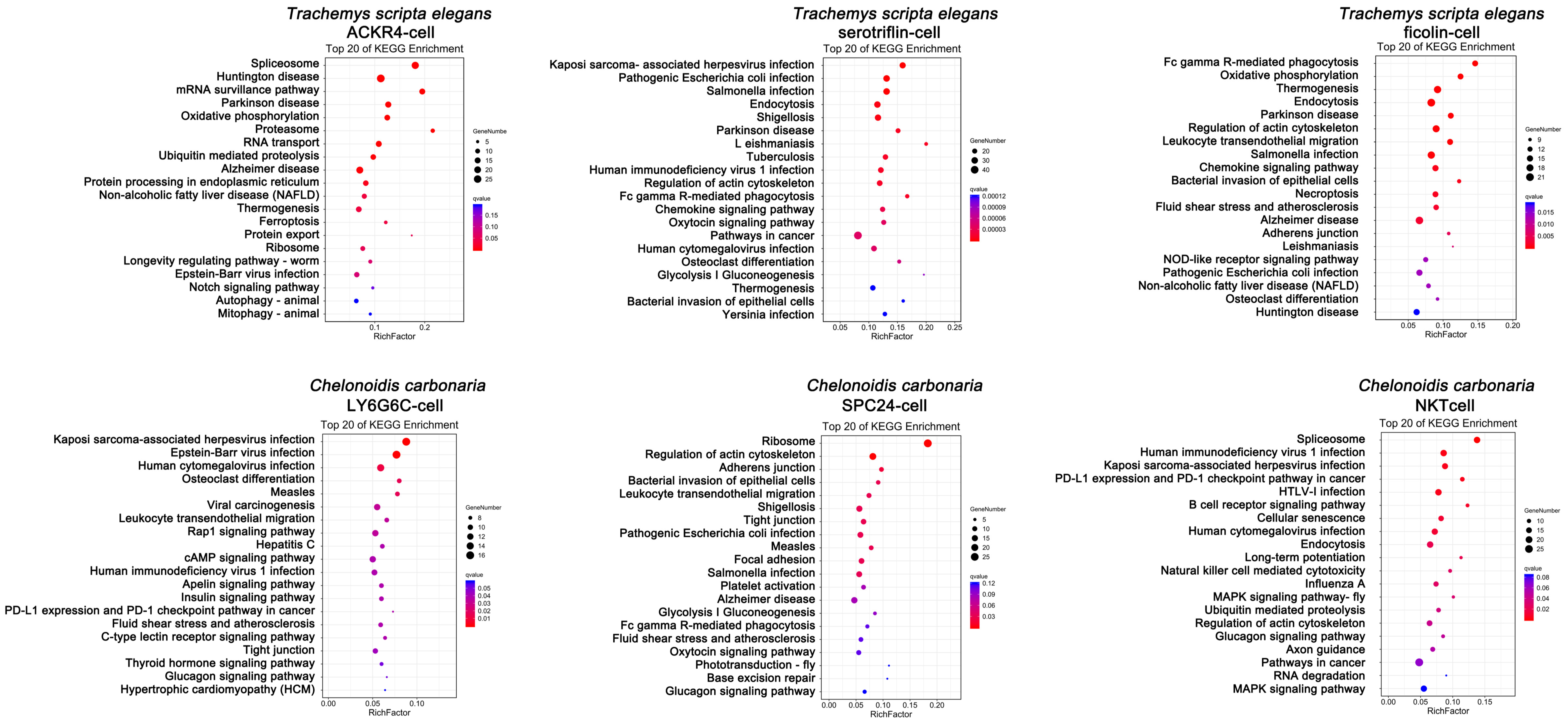
3.2. Innate Immune Function of Peripheral Blood in Testudines
3.3. Adaptive Immune Function of Peripheral Blood in Testudines
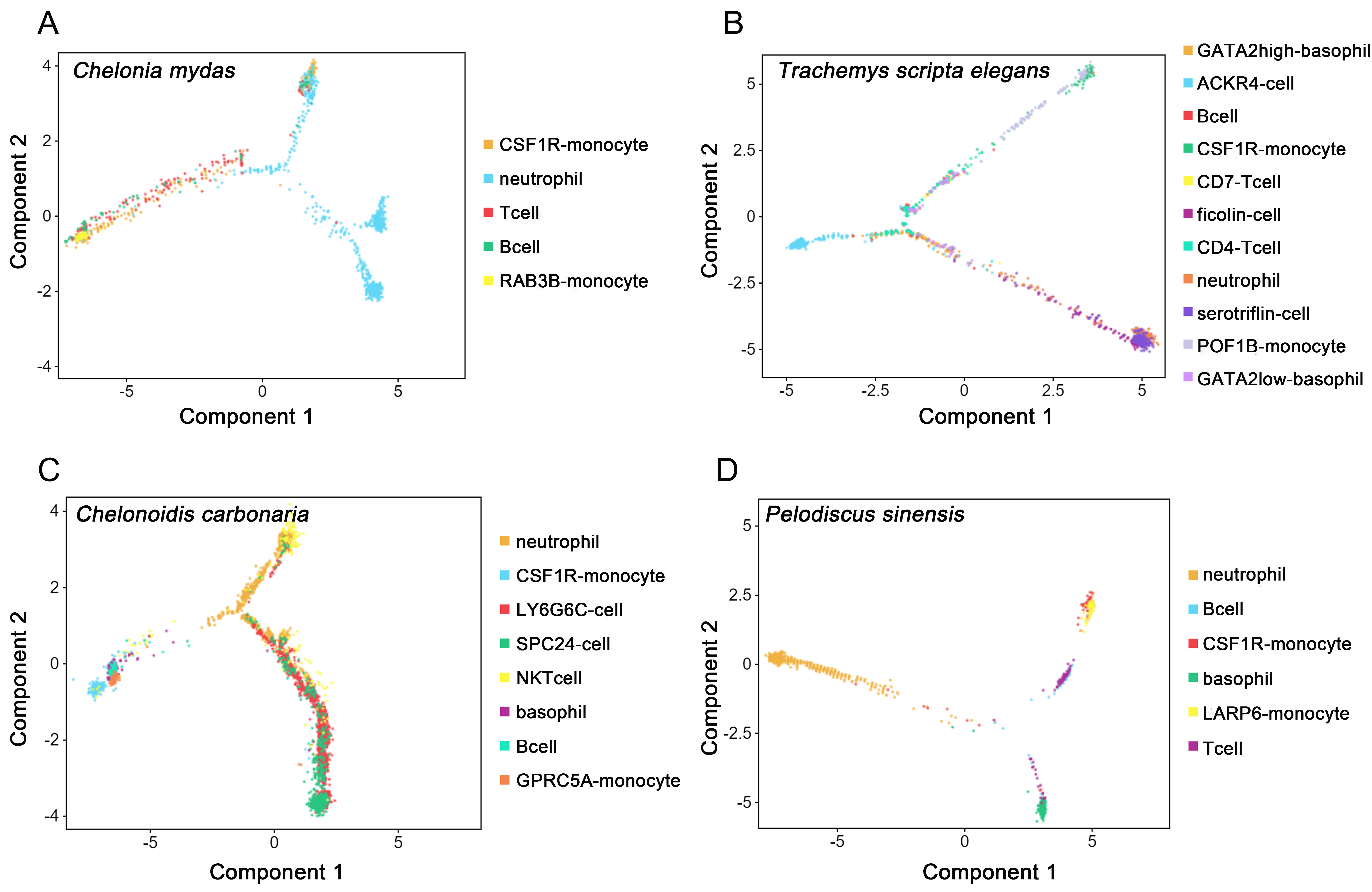
3.4. Developmental Trajectories of Peripheral Immune Cells in Testudines
4. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaffney, E.S. The comparative osteology of the Triassic turtle Proganochelys. Bull. Am. Mus. Nat. Hist. 1990, 194, 1–263. [Google Scholar]
- Krenz, J.G.; Naylor, G.J.P.; Shaffer, H.B.; Janzen, F.J. Molecular phylogenetics and evolution of turtles. Mol. Phylogenetics Evol. 2005, 37, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Rhodin, A.G.J.; Stanford, C.B.; Van Dijk, P.P.; Eisemberg, C.; Luiselli, L.; Mittermeier, R.A.; Hudson, R.; Horne, B.D.; Goode, E.V.; Kuchling, G. Global conservation status of turtles and tortoises (order Testudines). Chelonian Conserv. Biol. 2018, 17, 135–161. [Google Scholar] [CrossRef]
- Thomson, R.C.; Spinks, P.Q.; Shaffer, H.B. A global phylogeny of turtles reveals a burst of climate-associated diversification on continental margins. Proc. Natl. Acad. Sci. USA 2021, 118, e2012215118. [Google Scholar] [CrossRef]
- Zhang, G.; Cowled, C.; Shi, Z.; Huang, Z.Y.; Bishop-lilly, K.A.; Fang, X.D.; Wynne, J.W.; Xiong, Z.Q.; Baker, M.L.; Zhao, W. Comparative analysis of bat genomes provides insight into the evolution of flight and immunity. Science 2013, 339, 456–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schountz, T.; Baker, M.L.; Butler, J.; Munster, V. Immunological control of viral infections in bats and the emergence of viruses highly pathogenic to humans. Front. Immunol. 2017, 8, 1098–1117. [Google Scholar] [CrossRef]
- Casal, A.B.; Orós, J. Morphologic and cytochemical characteristics of blood cells of juvenile loggerhead sea turtles (Caretta caretta). Res. Vet. Sci. 2007, 82, 158–165. [Google Scholar] [CrossRef]
- Javanbakht, H.; Vaissi, S.; Parto, P. The morphological characterization of the blood cells in the three species of turtle and tortoise in Iran. Res. Zool. 2013, 3, 38–44. [Google Scholar]
- Hernández, J.D.; Castro, P.; Saavedra, P.; Ramírez, P.; Orós, J. Morphologic and cytochemical characteristics of the blood cells of the yellow-bellied slider (Trachemys scripta scripta). Anat. Histol. Embryol. 2017, 46, 446–455. [Google Scholar] [CrossRef] [Green Version]
- Work, T.M.; Raskin, R.E.; Balazs, G.H.; Whittaker, S.D. Morphologic and cytochemical characteristics of blood cells from Hawaiian green turtles. Am. J. Vet. Res. 1998, 59, 1252–1257. [Google Scholar]
- Koiwai, K.; Koyama, T.; Tsuda, S.; Toyoda, A.; Kikuchi, K.; Suzuki, H.; Kawano, R. Single-cell RNA-seq analysis reveals penaeid shrimp hemocyte subpopulations and cell differentiation process. eLife 2021, 10, e66954. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Chen, Y.; Huang, Z.; Xia, H.D.; Cheng, L.; Wu, H.; Zhang, Y.L.; Wang, F. Single-cell RNA sequencing analysis of shrimp immune cells identifies macrophage-like phagocytes. eLife 2022, 11, e80127. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Zhang, G.; Wang, W.X. Functional heterogeneity of immune defenses in molluscan oysters Crassostrea hongkongensis revealed by high-throughput single-cell transcriptome. Fish Shellfish Immunol. 2022, 120, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Siebert, S.; Farrell, J.A.; Cazet, J.F.; Abeykoon, Y.; Primack, A.S.; Schnitzler, C.E.; Juliano, C.E. Stem cell differentiation trajectories in Hydra resolved at single-cell resolution. Science 2019, 365, eaav9314. [Google Scholar] [CrossRef]
- Dong, F.; Hao, S.; Zhang, S.; Zhu, C.Y.; Cheng, H.; Yang, Z.N.; Hamey, F.K.; Wang, X.F.; Gao, A.; Wang, F.J. Differentiation of transplanted haematopoietic stem cells tracked by single-cell transcriptomic analysis. Nat. Cell Biol. 2020, 22, 630–639. [Google Scholar] [CrossRef]
- Xie, X.W.; Liu, M.Y.; Zhang, Y.W.; Wang, B.R.; Zhu, C.Y.; Wang, C.C.; Li, Q.; Huo, Y.Y.; Guo, J.J.; Xu, C.L. Single-cell transcriptomic landscape of human blood cells. Natl. Sci. Rev. 2021, 8, nwaa180. [Google Scholar] [CrossRef]
- Seminoff, J.A.; Resendiz, A.; Nichols, W.J. Home range of green turtles Chelonia mydas at a coastal foraging area in the Gulf of California, Mexico. Mar. Ecol. Prog. Ser. 2002, 242, 253–265. [Google Scholar] [CrossRef] [Green Version]
- Arthur, K.E.; Boyle, M.C.; Limpus, C.J. Ontogenetic changes in diet and habitat use in green sea turtle (Chelonia mydas) life history. Mar. Ecol. Prog. Ser. 2008, 362, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Tran, S.L.; Moorhead, D.L.; McKenna, K.C. Habitat selection by native turtles in a Lake Erie wetland, USA. Am. Midl. Nat. 2007, 158, 16–28. [Google Scholar] [CrossRef]
- Castanet, J. Age estimation and longevity in reptiles. Gerontology 1994, 40, 174–192. [Google Scholar] [CrossRef]
- McRobert, S.P.; Hopkins, D.T. The effects of dietary vitamin C on growth rates of juvenile slider turtles (Trachemys scripta elegans). J. Zoo Wildl. Med. 1998, 29, 419–422. [Google Scholar] [PubMed]
- Legler, J.M. Tortoises (Geochelone carbonaria) in Panama: Distribution and variation. Am. Midl. Nat. 1963, 70, 490–503. [Google Scholar] [CrossRef]
- Moskovits, D.K.; Bjorndal, K.A. Diet and food preferences of the tortoises Geochelone carbonaria and G. denticulata in northwestern Brazil. Herpetologica 1990, 46, 207–218. [Google Scholar]
- Adnagulov, E.V.; Maslova, I.V. On the distribution of Pelodiscus sinensis (Wiegmann, 1834) (Testudines: Trionychidae) in the Russian Far East. Herpetol. Petropolitana 2003, 12, 117–119. [Google Scholar]
- Works, A.J.; Olson, D.H. Diets of two nonnative freshwater turtle species (Trachemys scripta and Pelodiscus sinensis) in Kawai Nui Marsh, Hawaii. J. Herpetol. 2018, 52, 444–452. [Google Scholar] [CrossRef]
- Xiong, G.; Wang, X.Q.; Zhou, X.W.; Zeng, D.; Chen, Z.N.; Wang, P.; Kang, L. Genetic variation in the Chinese soft-shell turtles (Pelodiscus spp.) revealed by sequences of mitochondrial Cytb gene. Mitochondrial DNA Part A 2019, 30, 874–879. [Google Scholar] [CrossRef]
- Butler, A.; Hoffman, P.; Smibert, P.; Papalexi, E.; Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 2018, 36, 411–420. [Google Scholar] [CrossRef]
- Korsunsky, I.; Millard, N.; Fan, J.; Slowikowski, K.; Zhang, F.; Wei, K.; Baglaenko, Y.; Brenner, M.; Loh, P.; Raychaudhuri, S. Fast, sensitive and accurate integration of single-cell data with Harmony. Nat. Methods 2019, 16, 1289–1296. [Google Scholar] [CrossRef]
- Camp, J.G.; Sekine, K.; Gerber, T.; Loeffler-Wirth, H.; Binder, H.; Gac, M.; Kanton, S.; Kageyama, J.; Damm, G.; Seehofer, D. Multilineage communication regulates human liver bud development from pluripotency. Nature 2017, 546, 533–538. [Google Scholar] [CrossRef]
- Qiu, X.; Mao, Q.; Tang, Y.; Wang, L.; Chawla, R.; Pliner, H.A.; Trapnell, C. Reversed graph embedding resolves complex single-cell trajectories. Nat. Methods 2017, 14, 979–982. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.Y.; Li, P.P.; Gu, H.X.; Ye, M.B. Hematology, morphology, and ultrastructure of blood cells of juvenile olive ridley sea turtles (Lepidochelys olivacea). Chelonian Conserv. Biol. 2011, 10, 250–256. [Google Scholar] [CrossRef]
- da Cunha, H.F.; Carrijo, T.F.; Prestes, A.C.; Arruda, L.S.; Rezende, P.B.; Santos, T.; Brandão, D. Morphological analysis of peripheral blood cells of Chelonoidis carbonaria (Spix, 1824). Biosci. J. 2015, 31, 242–247. [Google Scholar]
- Knotková, Z.; Doubek, J.; Knotek, Z.; Hájková, P. Blood cell morphology and plasma biochemistry in Russian tortoises (Agrionemys horsfieldi). Acta Vet. Brno 2002, 71, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.Y.; Jia, W.T.; Fan, M.J.; Shao, X.; Li, Z.L.; Liu, Y.J.; Ma, Y.L.; Li, Y.X.; Li, R.; Tu, Q. Single-cell immune landscape of human recurrent miscarriage. Genom. Proteom. Bioinform. 2021, 19, 208–222. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Xie, L.J.; Ding, G.H.; Song, S.R.; Chen, L.Q.; Li, G.; Xia, M.; Han, D.D.; Zheng, Y.; Liu, J.; et al. Single-cell RNA sequencing of peripheral blood mononuclear cells from acute Kawasaki disease patients. Nat. Commun. 2021, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.G.; Lei, L.; Yang, X.F.; Ma, K.L.; Zheng, H.Q.; Su, Y.H.; Jiao, A.J.; Wang, X.; Liu, H.Y.; Zou, Y.J.; et al. Single-cell sequencing reveals antitumor characteristics of intratumoral immune cells in old mice. J. Immunother. Cancer 2021, 9, e002809. [Google Scholar] [CrossRef]
- Aguilar-Ruiz, S.R.; Torres-Aguilar, H.; González-Domínguez, É.; González-Pérez, J.N.G.; Vargas-Ayala, G.; Meraz-Ríos, M.A.; García-Zepeda, E.A.; Sánchez-Torres, C. Human CD16+ and CD16–monocyte subsets display unique effector properties in inflammatory conditions in vivo. J. Leukoc. Biol. 2011, 90, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Dentener, M.A.; Francot, G.J.M.; Buurman, W.A. Bactericidal/permeability-increasing protein, a lipopolysaccharide-specific protein on the surface of human peripheral blood monocytes. J. Infect. Dis. 1996, 173, 252–255. [Google Scholar] [CrossRef] [Green Version]
- Krautter, F.; Recio, C.; Hussain, M.T.; Lezama, D.R.; Maione, F.; Chimen, M.; Iqbal, A.J. Characterisation of endogenous Galectin-1 and-9 expression in monocyte and macrophage subsets under resting and inflammatory conditions. Biomed. Pharmacother. 2020, 130, 110595. [Google Scholar] [CrossRef]
- Kwok, I.; Becht, E.; Xia, Y.; Ng, M.; Teh, Y.C.; Tan, L.; Evrard, M.; Li, J.L.Y.; Tran, H.T.N.; Tan, Y.; et al. Combinatorial single-cell analyses of granulocyte-monocyte progenitor heterogeneity reveals an early uni-potent neutrophil progenitor. Immunity 2020, 53, 303–318.e5. [Google Scholar] [CrossRef]
- Hume, D.A.; Caruso, M.; Ferrari-Cestari, M.; Summers, K.M.; Pridans, C.; Irvine, K.M. Phenotypic impacts of CSF1R deficiencies in humans and model organisms. J. Leukoc. Biol. 2020, 107, 205–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, H.H.; Wang, L.; Li, J.J.; Luo, S.; Han, Q.Q.; Su, F.; Wei, J.; Wei, X.N.; Wu, J.P.; Li, B.; et al. Single-cell RNA-sequencing reveals distinct immune cell subsets and signaling pathways in IgA nephropathy. Cell Biosci. 2021, 11, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Sunderkötter, C.; Nikolic, T.; Dillon, M.J.; Van Rooijen, N.; Stehling, M.; Drevets, D.A.; Leenen, P.J. Subpopulations of mouse blood monocytes differ in maturation stage and inflammatory response. J. Immunol. 2004, 172, 4410–4417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukund, K.; Nayak, P.; Ashokkumar, C.; Rao, S.; Almeda, J.; Betancourt-Garcia, M.M.; Sindhi, R.; Subramaniam, S. Immune response in severe and non-severe coronavirus disease 2019 (COVID-19) infection: A mechanistic landscape. Front. Immunol. 2021, 12, 738073. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Qi, X.; Liu, B.; Huang, H. The STAT5–GATA2 pathway is critical in basophil and mast cell differentiation and maintenance. J. Immunol. 2015, 194, 4328–4338. [Google Scholar] [CrossRef] [Green Version]
- Yamanishi, Y.; Miyake, K.; Iki, M.; Tsutsui, H.; Karasuyama, H. Recent advances in understanding basophil-mediated Th2 immune responses. Immunol. Rev. 2017, 278, 237–245. [Google Scholar] [CrossRef]
- Merluzzi, S.; Betto, E.; Ceccaroni, A.A.; Magris, R.; Giunta, M.; Mion, F. Mast cells, basophils and B cell connection network. Mol. Immunol. 2015, 63, 94–103. [Google Scholar] [CrossRef]
- Lantz, O.; Bendelac, A. An invariant T cell receptor alpha chain is used by a unique subset of major histocompatibility complex class I-specific CD4+ and CD4-8-T cells in mice and humans. J. Exp. Med. 1994, 180, 1097–1106. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.K.; Tripathi, P.; Cardell, S.L. Type II NKT cells: An elusive population with immunoregulatory properties. Front. Immunol. 2018, 9, 1969. [Google Scholar] [CrossRef]
- Medvedovic, J.; Ebert, A.; Tagoh, H.; Busslinger, M. Pax5: A master regulator of B cell development and leukemogenesis. Adv. Immunol. 2011, 111, 179–206. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, R.; Ma, G.; Zhai, X.; Shi, H.; Wang, J. Comparison of the Single-Cell Immune Landscape of Testudines from Different Habitats. Cells 2022, 11, 4023. https://doi.org/10.3390/cells11244023
Guo R, Ma G, Zhai X, Shi H, Wang J. Comparison of the Single-Cell Immune Landscape of Testudines from Different Habitats. Cells. 2022; 11(24):4023. https://doi.org/10.3390/cells11244023
Chicago/Turabian StyleGuo, Rui, Guangwei Ma, Xiaofei Zhai, Haitao Shi, and Jichao Wang. 2022. "Comparison of the Single-Cell Immune Landscape of Testudines from Different Habitats" Cells 11, no. 24: 4023. https://doi.org/10.3390/cells11244023