
Editing of a Novel Cd Uptake-Related Gene CUP1 Contributes to Reducing Cd Accumulations in Arabidopsis thaliana and Brassica napus

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phylogenetic Analysis of AtCUP1
2.2. Cadmium Transport Activities Assay in Yeast
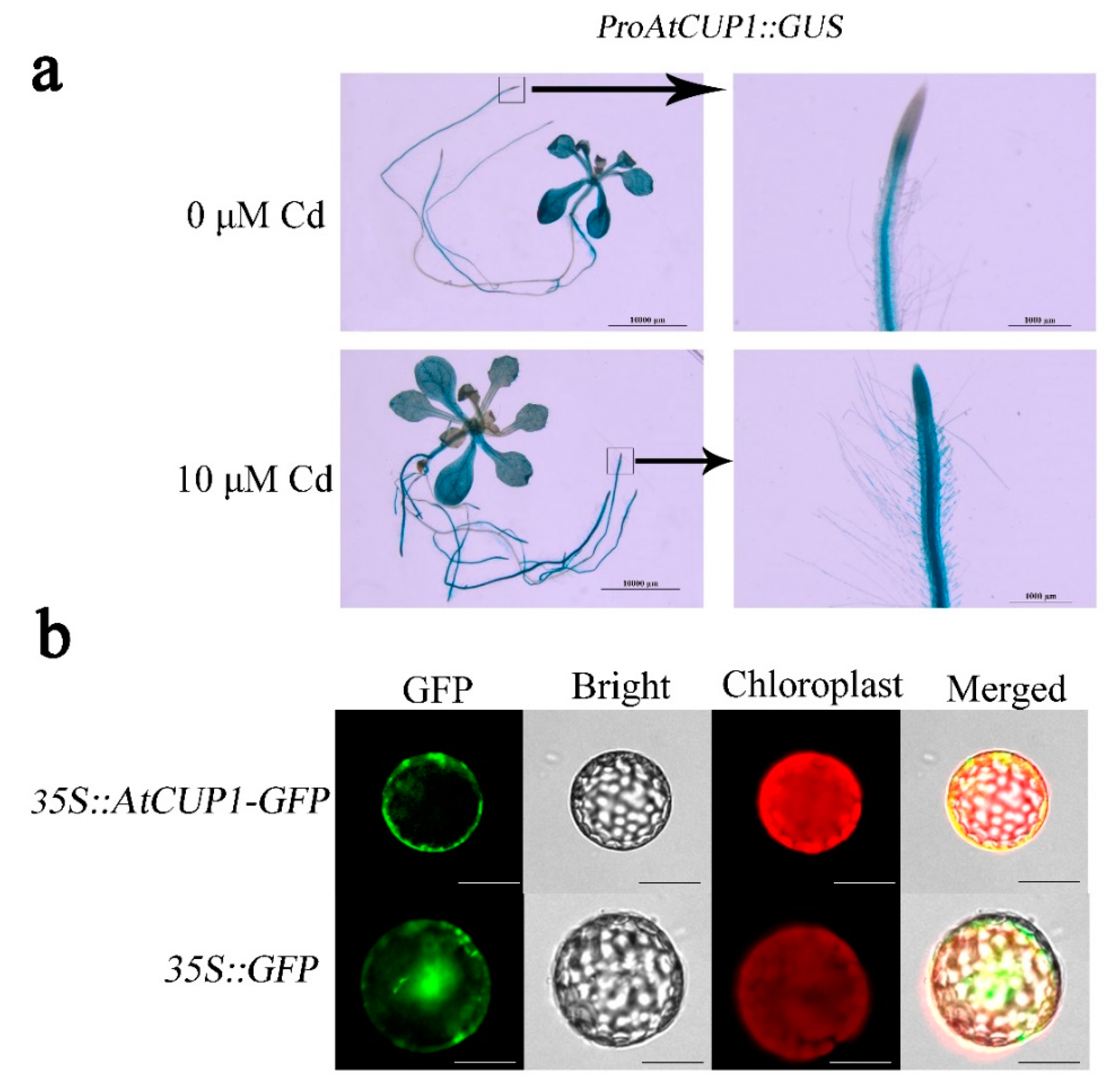
2.3. Subcellular Localization Assay
2.4. Analysis of AtCUP1 Promoter Activity
2.5. Preparation of the AtCUP1-Overexpressing Transgenic Lines, atcup1 Mutant and CRISPR-AtCUP1 Lines
2.6. Cd Treatment Experiment of Different Growth Stages
2.7. Localization Experiment of Cd in Tissues
2.8. Quantitative Real-Time PCR
2.9. Construction and Identification of BnCUP1-Edited Lines
2.10. Hydroponic Experiments of BnCUP1-Edited Lines
2.11. The Field Experiment for CRISPR-BnCUP1 Lines
2.12. Determination of Metal Elements
3. Results
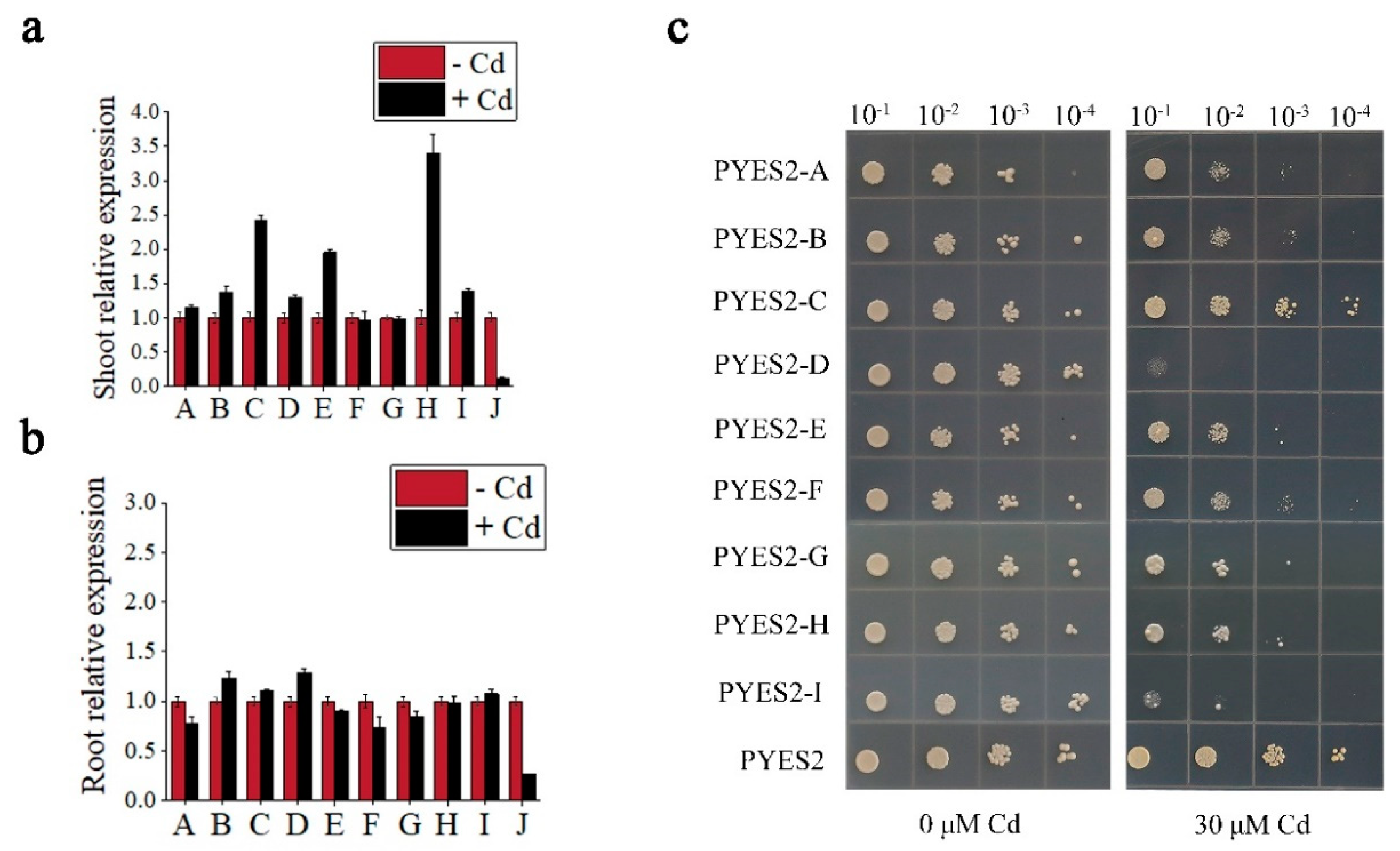
3.1. Screening of Cd Uptake-Related Genes
3.2. Expression Pattern Analysis of AtCUP1
3.3. AtCUP1 Was Confirmed to Be Involved in Cd Uptake in A. thaliana
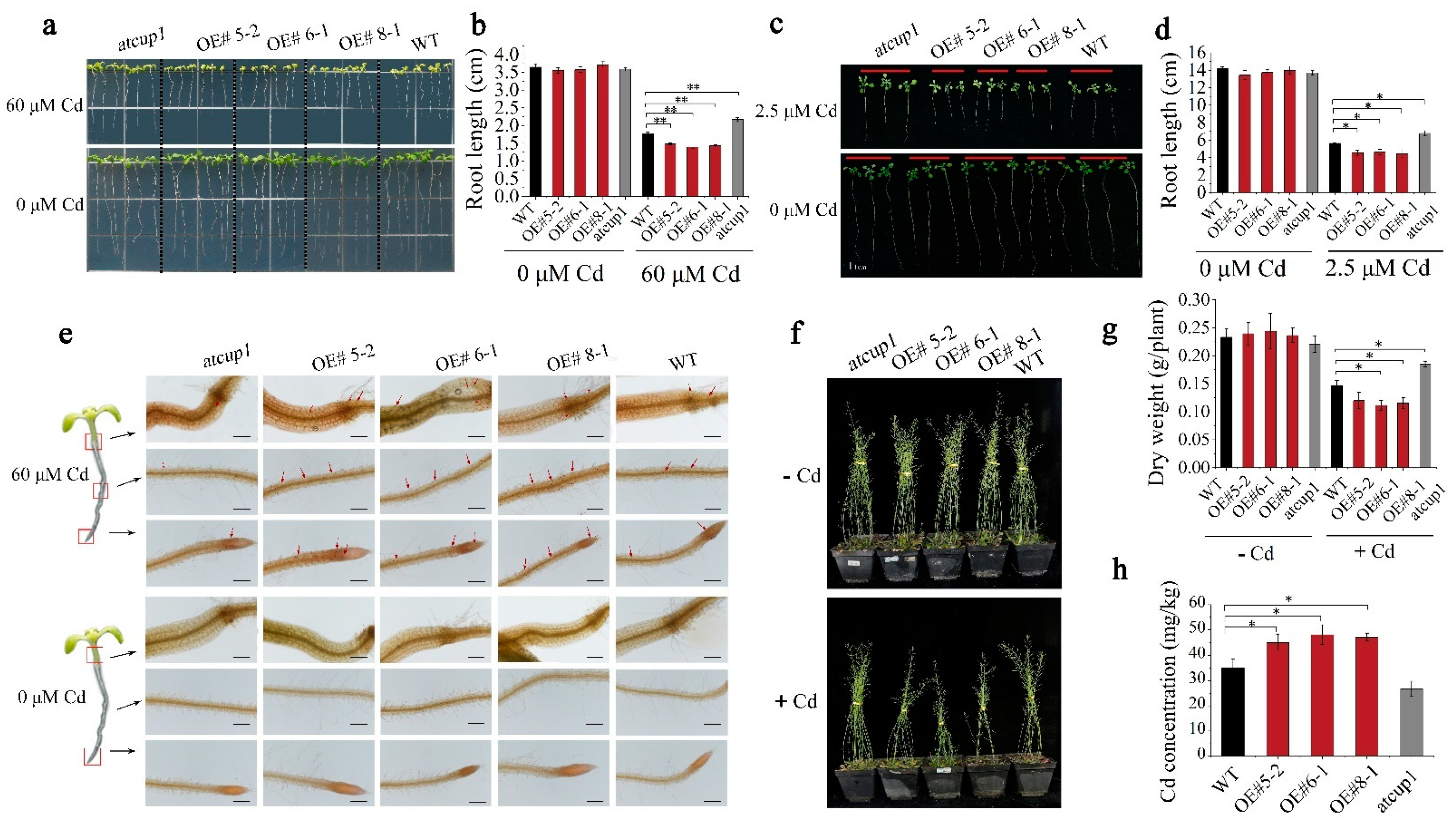
3.4. Various AtCUP1-Edited Lines Showed Divergent Cd Accumulation
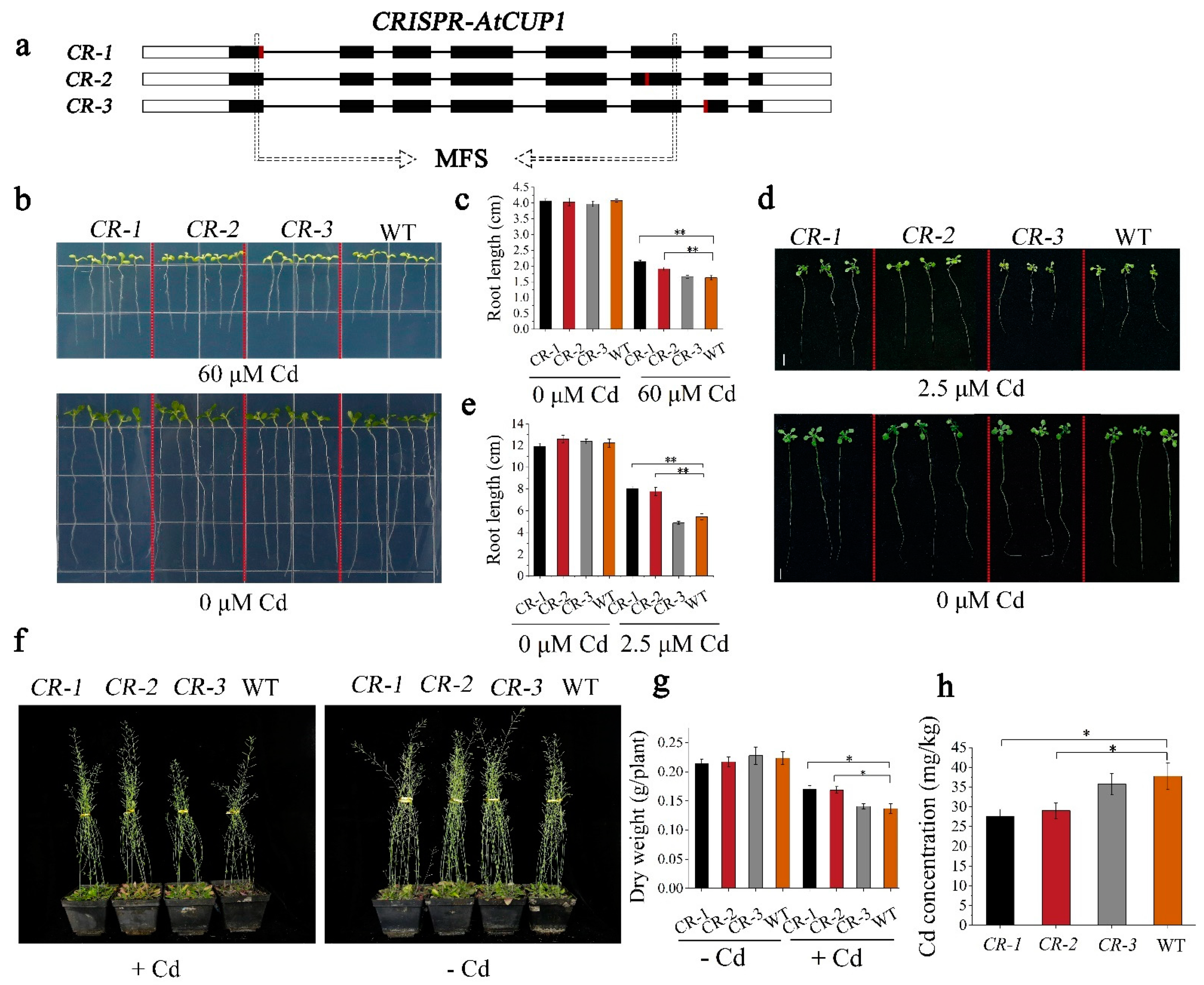
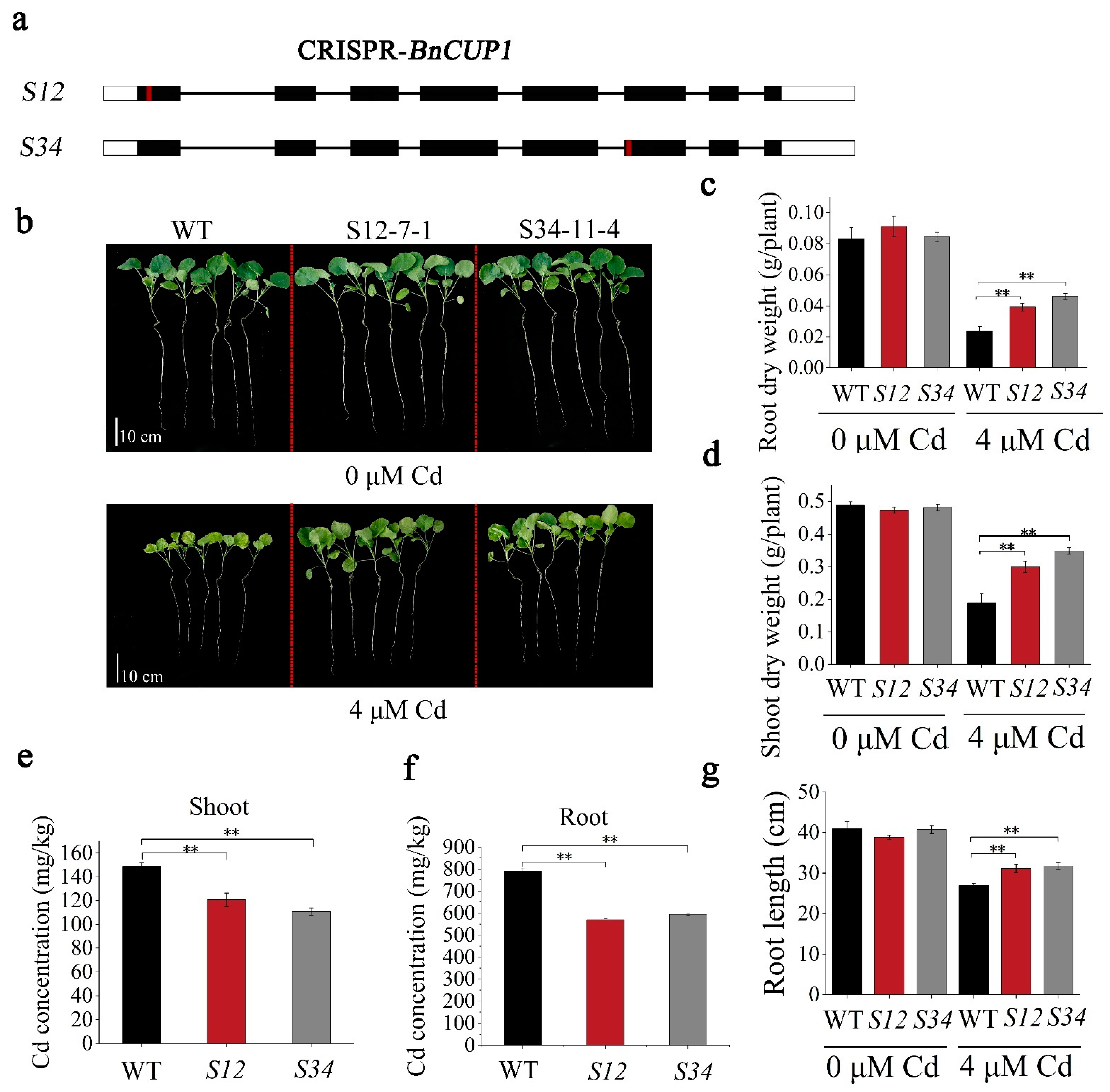
3.5. CRISPR-BnCUP1 Mutation Reduced Cd Accumulation in B. napus
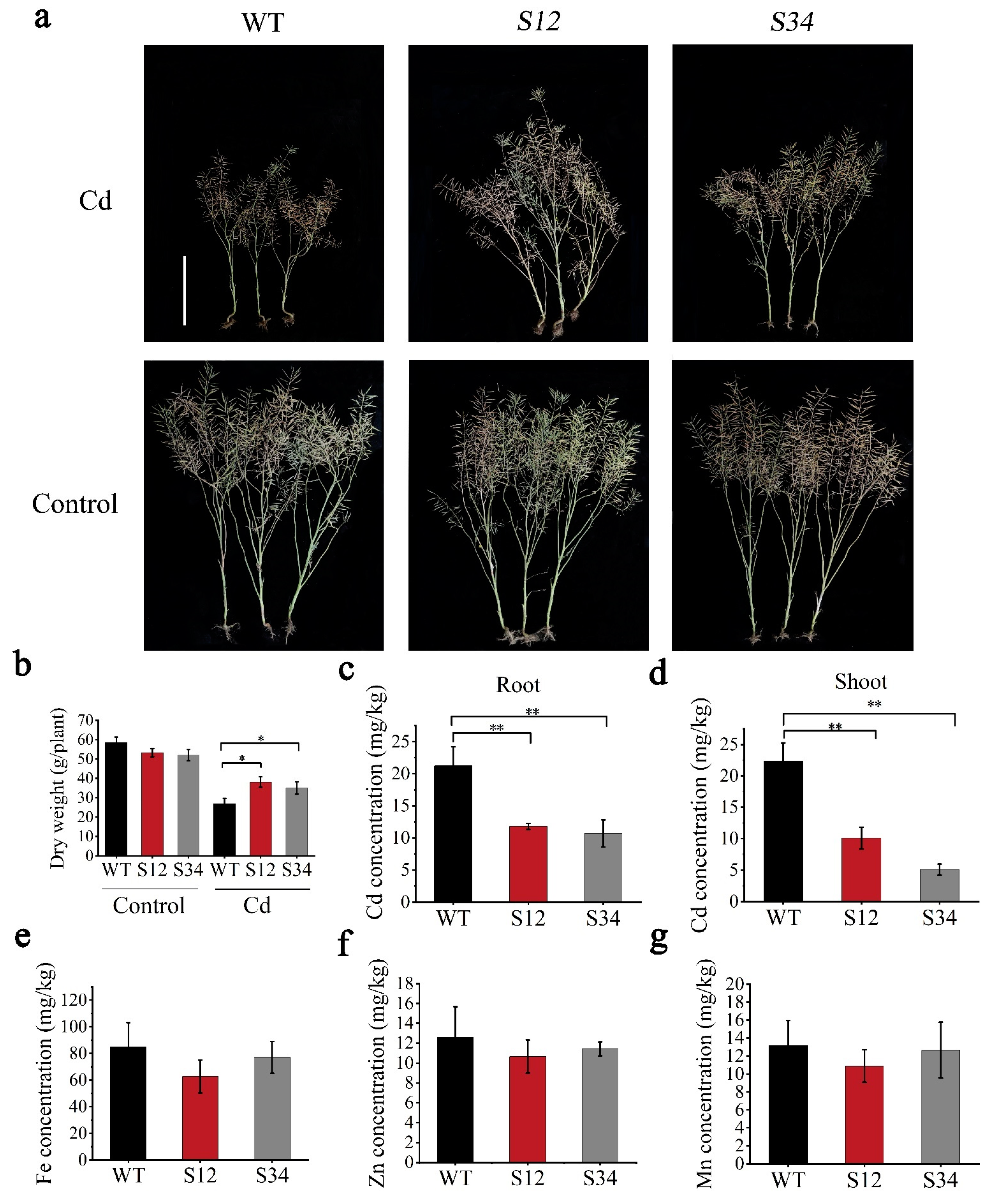
3.6. BnCUP1-Edited B. napus Was Identified as a Compelling Low-Cd Germplasm
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, M.A.; Khan, S.; Khan, A.; Alam, M. Soil contamination with cadmium, consequences and remediation using organic amendments. Sci. Total Environ. 2017, 601, 1591–1605. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Dumat, C.; Khalid, S.; Niazi, N.K.; Antunes, P.M.C. Cadmium Bioavailability, Uptake, Toxicity and Detoxification in Soil-Plant System. Rev. Environ. Contam. Toxicol. 2017, 241, 73–137. [Google Scholar] [CrossRef] [PubMed]
- Lohani, N.; Jain, D.; Singh, M.B.; Bhalla, P.L. Engineering Multiple Abiotic Stress Tolerance in Canola, Brassica napus. Front. Plant Sci. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Chen, L.; Wan, H.; Qian, J.; Guo, J.; Sun, C.; Wen, J.; Yi, B.; Ma, C.; Tu, J.; Song, L.; et al. Genome-Wide Association Study of Cadmium Accumulation at the Seedling Stage in Rapeseed (Brassica napus L.). Front. Plant Sci. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodziak-Dopierała, B.; Kwapuliński, J.; Sobczyk, K.; Wiechuła, D. Analysis of the Content of Cadmium and Zinc in Parts of the Human Hip Joint. Biol. Trace Elem. Res. 2015, 163, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Shu, Y. Cadmium Transporters in the Kidney and Cadmium-Induced Nephrotoxicity. Int. J. Mol. Sci. 2015, 16, 1484–1494. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S.-I.; Suzui, N.; Nagasaka, T.; Komatsu, F.; Ishioka, N.S.; Ito-Tanabata, S.; Kawachi, N.; Rai, H.; Hattori, H.; Chino, M.; et al. Application of glutathione to roots selectively inhibits cadmium transport from roots to shoots in oilseed rape. J. Exp. Bot. 2013, 64, 1073–1081. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Yin, X.; Bañuelos, G.S.; Lin, Z.Q.; Liu, Y.; Li, M.; Yuan, L. Indications of Selenium Protection against Cadmium and Lead Toxicity in Oilseed Rape (Brassica napus L.). Front. Plant Sci. 2016, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Zhu, Z.; Wu, X.; Liu, Z.; Zou, J.; Chen, Y.; Su, N.; Cui, J. Lower cadmium accumulation and higher antioxidative capacity in edible parts of Brassica campestris L. seedlings applied with glutathione under cadmium toxicity. Environ. Sci. Pollut. Res. Int. 2019, 26, 13235–13245. [Google Scholar] [CrossRef]
- Qi, W.Y.; Li, Q.; Chen, H.; Liu, J.; Xing, S.F.; Xu, M.; Yan, Z.; Song, C.; Wang, S.G. Selenium nanoparticles ameliorate Brassica napus L. cadmium toxicity by inhibiting the respiratory burst and scavenging reactive oxygen species. J. Hazard Mater. 2021, 417, 1–10. [Google Scholar] [CrossRef]
- Yan, H.; Xu, W.; Xie, J.; Gao, Y.; Wu, L.; Sun, L.; Feng, L.; Chen, X.; Zhang, T.; Dai, C.; et al. Variation of a major facilitator superfamily gene contributes to differential cadmium accumulation between rice subspecies. Nat. Commun. 2019, 10, 2562–2574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.H. Reduction in cadmium accumulation in japonica rice grains by CRISPR/Cas9-mediated editing of OsNRAMP5. J. Integr. Agr. 2019, 18, 210–219. [Google Scholar] [CrossRef]
- Verret, F.; Gravot, A.; Auroy, P.; Leonhardt, N.; David, P.; Nussaume, L.; Vavasseur, A.; Richaud, P. Overexpression of AtHMA4 enhances root-to-shoot translocation of zinc and cadmium and plant metal tolerance. FEBS Lett. 2004, 576, 306–3012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morel, M.; Crouzet, J.; Gravot, A.; Auroy, P.; Leonhardt, N.; Vavasseur, A.; Richaud, P. AtHMA3, a P1B-ATPase allowing Cd/Zn/Co/Pb vacuolar storage in Arabidopsis. Plant Physiol. 2009, 149, 894–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, R.; Ishimaru, Y.; Senoura, T.; Shimo, H.; Ishikawa, S.; Arao, T.; Nakanishi, H.; Nishizawa, N.K. The OsNRAMP1 iron transporter is involved in Cd accumulation in rice. J. Exp. Bot. 2011, 62, 4843–4850. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, S.; Gonzalez-Nieto, D.; Ghiaur, G.; Dunn, S.K.; Ficker, A.M.; Murali, B.; Madhu, M.; Gutstein, D.E.; Fishman, G.I.; Barrio, L.C.; et al. Ion-beam irradiation, gene identification, and marker-assisted breeding in the development of low-cadmium rice. Proc. Natl. Acad. Sci. USA 2012, 109, 19166–19171. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Li, Y.; Fu, Y.; Xie, H.; Song, S.; Qiu, M.; Wen, J.; Chen, M.; Chen, G.; Tian, Y.; et al. Mutation at Different Sites of Metal Transporter Gene OsNramp5 Affects Cd Accumulation and Related Agronomic Traits in Rice (Oryza sativa L.). Front. Plant Sci. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Chang, J.D.; Huang, S.; Yamaji, N.; Zhang, W.; Ma, J.F.; Zhao, F.J. OsNRAMP1 transporter contributes to cadmium and manganese uptake in rice. Plant Cell Environ. 2020, 43, 2476–2491. [Google Scholar] [CrossRef]
- Chu, C.; Huang, R.; Liu, L.; Tang, G.; Xiao, J.; Yoo, H.; Yuan, M. The rice heavy metal transporter OsNRAMP1 regulates disease resistance by modulating ROS homeostasis. Plant Cell Environ. 2022, 45, 1109–1126. [Google Scholar] [CrossRef]
- Wong, C.K.E.; Cobbett, C.S. HMA P-type ATPases are the major mechanism for root-to-shoot Cd translocation in Arabidopsis thaliana. New Phytol. 2009, 181, 71–78. [Google Scholar] [CrossRef]
- Cun, P.; Sarrobert, C.; Richaud, P.; Chevalier, A.; Soreau, P.; Auroy, P.; Gravot, A.; Baltz, A.; Leonhardt, N.; Vavasseur, A. Modulation of Zn/Cd P(1B2)-ATPase activities in Arabidopsis impacts differently on Zn and Cd contents in shoots and seeds. Metallomics 2014, 6, 2109–2116. [Google Scholar] [CrossRef] [PubMed]
- Madej, M.G.; Dang, S.Y.; Yan, N.E.; Kaback, H.R. Evolutionary mix-and-match with MFS transporters. Proc. Natl. Acad. Sci. USA 2013, 110, 5870–5874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haydon, M.J.; Cobbett, C.S. A Novel Major Facilitator Superfamily Protein at the Tonoplast Influences Zinc Tolerance and Accumulation in Arabidopsis. Plant Physiol. 2007, 143, 1705–1719. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.H. EvolView, an online tool for visualizing, annotating and managing phylogenetic trees. Nucleic Acids Res. 2012, 40, W569–W572. [Google Scholar] [CrossRef]
- Roberts, C. Methods in Yeast Genetics. Methods Med. Res. 1950, 3, 37–50. [Google Scholar]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [Green Version]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chen, H. A novel CRISPR/Cas9 system for efficiently generating Cas9-free multiplex mutants in Arabidopsis. aBIOTECH 2020, 1, 6–14. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Bai, J.; Wang, J.; Le, S.; Wang, M.; Zhao, Y. Variations in cadmium accumulation and distribution among different oilseed rape cultivars in Chengdu Plain in China. Environ. Sci. Pollut. R. 2019, 26, 3415–3427. [Google Scholar] [CrossRef]
- Clabeaux, B.L.; Navarro, D.A.; Aga, D.S.; Bisson, M.A. Cd tolerance and accumulation in the aquatic macrophyte, Chara australis: Potential use for charophytes in phytoremediation. Environ. Sci. Technol. 2011, 45, 5332–5338. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.L.; Dong, L.; Wang, Z.P.; Zhang, H.Y.; Han, C.Y.; Liu, B.; Wang, X.C.; Chen, Q.J. A CRISPR/Cas9 toolkit for multiplex genome editing in plants. BMC Plant Biol. 2014, 14, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, T.; Yu, X.; Yang, H.; Gao, Q.; Ji, H.; Wang, Y.; Yan, G.; Peng, Y.; Luo, H.; Liu, K.; et al. Development and Validation of an Effective CRISPR/Cas9 Vector for Efficiently Isolating Positive Transformants and Transgene-Free Mutants in a Wide Range of Plant Species. Front. Plant Sci. 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, Y.; Yu, K.; Cai, S.; Hu, L.; Amoo, O.; Xu, L.; Yang, Y.; Ma, B.; Jiao, Y.; Zhang, C.; et al. Targeted mutagenesis of BnTT8 homologs controls yellow seed coat development for effective oil production in Brassica napus L. Plant Biotechnol. J. 2020, 18, 1153–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, D.; Jiang, L.; Mason, A.S.; Xiao, M.; Zhu, L.; Li, L.; Zhou, Q.; Shen, C.; Huang, C. Research progress and strategies for multifunctional rapeseed: A case study of China. J. Int. Agric. 2016, 15, 1673–1684. [Google Scholar] [CrossRef] [Green Version]
- Su, D.; Xing, J.; Jiao, W.; Wong, W. Cadmium uptake and speciation changes in the rhizosphere of cadmium accumulator and non-accumulator oilseed rape varieties. J. Environ. Sci. 2009, 21, 1125–1128. [Google Scholar] [CrossRef]
- Cao, X.; Luo, J.; Wang, X.; Chen, Z.; Liu, G.; Khan, M.B.; Kang, K.J.; Feng, Y.; He, Z.; Yang, X. Responses of soil bacterial community and Cd phytoextraction to a Sedum alfredii-oilseed rape (Brassica napus L. and Brassica juncea L.) intercropping system. Sci. Total. Environ. 2020, 723, 138152. [Google Scholar] [CrossRef]
- Cao, X.; Wang, X.; Tong, W.; Gurajala, H.K.; He, Z.; Yang, X. Accumulation and distribution of cadmium and lead in 28 oilseed rape cultivars grown in a contaminated field. Environ. Sci. Pollut. Res. Int. 2020, 27, 2400–2411. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, M.; Tian, S.; Lu, L.; Shohag, M.J.; Yang, X. Metallothionein 2 (SaMT2) from Sedum alfredii Hance confers increased Cd tolerance and accumulation in yeast and tobacco. PLoS ONE 2014, 9, e102750. [Google Scholar] [CrossRef]
- Takahashi, R.; Ishimaru, Y.; Nakanishi, H.; Nishizawa, N.K. Role of the iron transporter OsNRAMP1 in cadmium uptake and accumulation in rice. Plant Signal. Behav. 2011, 6, 1813–1816. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Ogo, Y.; Senoura, T.; Nishizawa, N.K.; Nakanishi, H. The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice. Plant Cell Environ. 2012, 35, 1948–1957. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Wang, P.; Wang, P.; Yang, M.; Lian, X.; Tang, Z.; Huang, C.F.; Salt, D.E.; Zhao, F.J. A loss-of-function allele of OsHMA3 associated with high cadmium accumulation in shoots and grain of Japonica rice cultivars. Plant Cell Environ. 2016, 39, 1941–1954. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Mao, B.; Li, Y.; Lv, Q.; Zhang, L.; Chen, C.; He, H.; Wang, W.; Zeng, X.; Shao, Y.; et al. Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield. Sci. Rep. 2017, 7, 14438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, R.; Acarkan, A.; Boivin, K. Comparative structural genomics in the Brassicaceae family. Plant Physiol. Biochem. 2001, 39, 253–262. [Google Scholar] [CrossRef]
- Mun, J.H.; Yu, H.J.; Park, S.; Park, B.S. Genome-wide identification of NBS-encoding resistance genes in Brassica rapa. Mol. Genet. Genom. 2009, 282, 617–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, A.S.; Snowdon, R.J. Oilseed rape: Learning about ancient and recent polyploid evolution from a recent crop species. Plant Biol. 2016, 18, 883–892. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material | Branch Number | Plant Height (cm) | Silique Number per Plant | Number of Seeds per Silique | 1000-Seed Weight (g) | Yield/Plant (g) |
|---|---|---|---|---|---|---|
| Westar | 7.47 ± 0.47 | 138.38 ± 4.90 | 246.40 ± 16.09 | 13.40 ± 0.52 | 3.70 ± 0.10 | 12.07 ± 0.89 |
| S12 | 8.80 ± 0.70 | 147.93 ± 3.38 | 299.67 ± 21.30 | 16.64 ± 0.48 ** | 3.43 ± 0.08 | 17.04 ± 1.27 ** |
| S34 | 8.67 ± 0.77 | 150.65 ± 2.50 * | 303.13 ± 26.30 | 17.59 ± 0.40 ** | 3.42 ± 0.10 * | 17.68 ± 1.29 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, J.; Bai, J.; Liu, S.; Fu, J.; Zhang, Y.; Luo, T.; Ren, H.; Wang, R.; Zhao, Y. Editing of a Novel Cd Uptake-Related Gene CUP1 Contributes to Reducing Cd Accumulations in Arabidopsis thaliana and Brassica napus. Cells 2022, 11, 3888. https://doi.org/10.3390/cells11233888
Yao J, Bai J, Liu S, Fu J, Zhang Y, Luo T, Ren H, Wang R, Zhao Y. Editing of a Novel Cd Uptake-Related Gene CUP1 Contributes to Reducing Cd Accumulations in Arabidopsis thaliana and Brassica napus. Cells. 2022; 11(23):3888. https://doi.org/10.3390/cells11233888
Chicago/Turabian StyleYao, Junyu, Jiuyuan Bai, Sha Liu, Jingyan Fu, Ying Zhang, Tianshun Luo, Hongpei Ren, Rui Wang, and Yun Zhao. 2022. "Editing of a Novel Cd Uptake-Related Gene CUP1 Contributes to Reducing Cd Accumulations in Arabidopsis thaliana and Brassica napus" Cells 11, no. 23: 3888. https://doi.org/10.3390/cells11233888