
Alignment of Skeletal Muscle Cells Facilitates Acetylcholine Receptor Clustering and Neuromuscular Junction Formation with Co-Cultured Human iPSC-Derived Motor Neurons

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fabrication of Cell Culture Substrates with Linear Patterns
2.2. Cell Culture on the Micropatterned PDMS Substrate
2.3. Fluorescent Staining of Cells Cultured on the PDMS Substrates
2.4. Real-Time PCR
2.5. Co-Culture of Human SkM Myotube and Human iPSC-Derived MNs
2.6. Assessment of Functionality of NMJs
2.7. Statistical Analysis
3. Results and Discussion
3.1. Alignment and AChR Clustering of SkM Cells Cultured on the Surface with Linear Grooves
3.2. Expression Analysis of Key AChR Clustering Genes in SkM Cells Cultured on a Surface with Linear Grooves
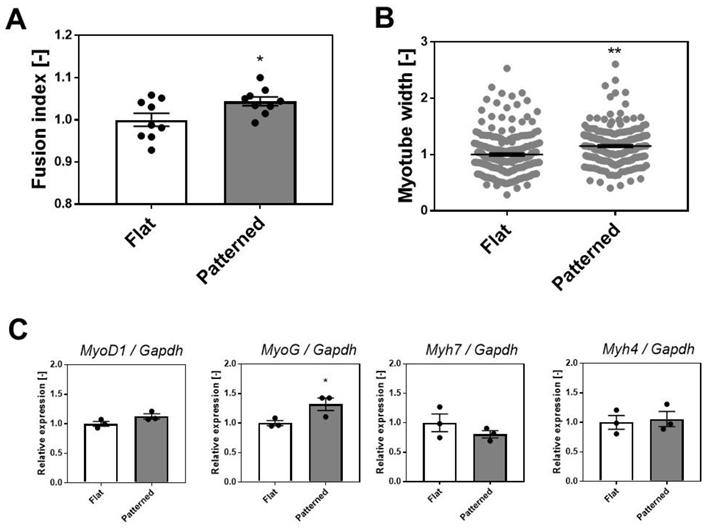
3.3. Effects of Patterned Surface on the Differentiation of SkM Cells
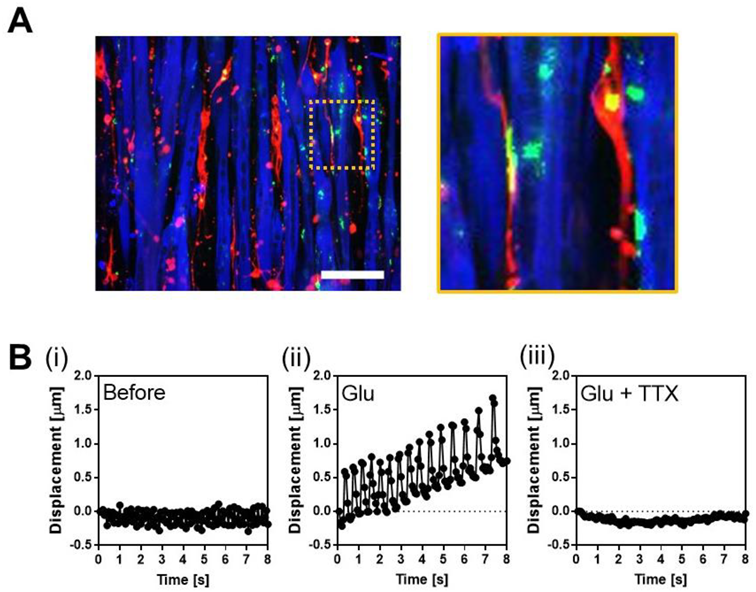
3.4. Co-Culture of Human SkM Cells and Human iPS-Derived Motor Neurons on a Surface with Linear Grooves
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burden, S.J. SnapShot: Neuromuscular Junction. Cell 2011, 144, 826–826.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Yang, F.; Xie, Z.; Lu, B. Intracellular Ca2+ and Ca2+/calmodulin-dependent kinase II mediate acute potentiation of neurotransmitter release by neurotrophin-3. J. Cell Biol. 2000, 149, 783–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.F.; Gonzalez, M.; Stancescu, M.; Vandenburgh, H.H.; Hickman, J.J. Neuromuscular junction formation between human stem cell-derived motoneurons and human skeletal muscle in a defined system. Biomaterials 2011, 32, 9602–9611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, M.; Rumsey, J.W.; Bhargava, N.; Stancescu, M.; Hickman, J.J. A defined long-term in vitro tissue engineered model of neuromuscular junctions. Biomaterials 2010, 31, 4880–4888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cun, X.; Hosta-Rigau, L. Topography: A Biophysical Approach to Direct the Fate of Mesenchymal Stem Cells in Tissue Engineering Applications. Nanomaterials 2020, 10, 2070. [Google Scholar] [CrossRef]
- Gui, N.; Xu, W.; Myers, D.E.; Shukla, R.; Tang, H.P.; Qian, M. The effect of ordered and partially ordered surface topography on bone cell responses: A review. Biomater. Sci. 2018, 6, 250–264. [Google Scholar] [CrossRef]
- Shimizu, K.; Fujita, H.; Nagamori, E. Alignment of skeletal muscle myoblasts and myotubes using linear micropatterned surfaces ground with abrasives. Biotechnol. Bioeng. 2009, 103, 631–638. [Google Scholar] [CrossRef]
- Huang, N.F.; Patel, S.; Thakar, R.G.; Wu, J.; Hsiao, B.S.; Chu, B.; Lee, R.J.; Li, S. Myotube assembly on nanofibrous and micropatterned polymers. Nano Lett. 2006, 6, 537–542. [Google Scholar] [CrossRef]
- Lam, M.T.; Sim, S.; Zhu, X.Y.; Takayama, S. The effect of continuous wavy micropatterns on silicone substrates on the alignment of skeletal muscle myoblasts and myotubes. Biomaterials 2006, 27, 4340–4347. [Google Scholar] [CrossRef]
- Ko, E.; Yu, S.J.; Pagan-Diaz, G.J.; Mahmassani, Z.; Boppart, M.D.; Im, S.G.; Bashir, R.; Kong, H. Matrix Topography Regulates Synaptic Transmission at the Neuromuscular Junction. Adv. Sci. 2019, 6, 1801521. [Google Scholar] [CrossRef]
- Yamaoka, N.; Shimizu, K.; Imaizumi, Y.; Ito, T.; Okada, Y.; Honda, H. Open-Chamber Co-Culture Microdevices for Single-Cell Analysis of Skeletal Muscle Myotubes and Motor Neurons with Neuromuscular Junctions. Biochip J. 2019, 13, 127–132. [Google Scholar] [CrossRef]
- Shimizu, K.; Genma, R.; Gotou, Y.; Nagasaka, S.; Honda, H. Three-Dimensional Culture Model of Skeletal Muscle Tissue with Atrophy Induced by Dexamethasone. Bioengineering 2017, 4, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiomi, K.; Kiyono, T.; Okamura, K.; Uezumi, M.; Goto, Y.; Yasumoto, S.; Shimizu, S.; Hashimoto, N. CDK4 and cyclin D1 allow human myogenic cells to recapture growth property without compromising differentiation potential. Gene Ther. 2011, 18, 857–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagashima, T.; Hadiwidjaja, S.; Ohsumi, S.; Murata, A.; Hisada, T.; Kato, R.; Okada, Y.; Honda, H.; Shimizu, K. In Vitro Model of Human Skeletal Muscle Tissues with Contractility Fabricated by Immortalized Human Myogenic Cells. Adv. Biosyst. 2020, 4, e2000121. [Google Scholar] [CrossRef] [PubMed]
- Enomae, T.; Han, Y.H.; Isogai, A. Nondestructive determination of fiber orientation distribution of paper surface by image analysis. Nord. Pulp Pap. Res. J. 2006, 21, 253–259. [Google Scholar] [CrossRef]
- Bai, Y.; Guo, D.; Sun, X.; Tang, G.; Liao, T.; Peng, Y.; Xu, J.; Shi, L. Balanced Rac1 activity controls formation and maintenance of neuromuscular acetylcholine receptor clusters. J. Cell Sci. 2018, 131, jcs215251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Nagashima, T.; Shimizu, K.; Matsumoto, R.; Honda, H. Selective Elimination of Human Induced Pluripotent Stem Cells Using Medium with High Concentration of L-Alanine. Sci. Rep. 2018, 8, 12427. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Yamaoka, N.; Imaizumi, Y.; Nagashima, T.; Furutani, T.; Ito, T.; Okada, Y.; Honda, H.; Shimizu, K. Development of a human neuromuscular tissue-on-a-chip model on a 24-well-plate-format compartmentalized microfluidic device. Lab Chip 2021, 21, 1897–1907. [Google Scholar] [CrossRef]
- Shimojo, D.; Onodera, K.; Doi-Torii, Y.; Ishihara, Y.; Hattori, C.; Miwa, Y.; Tanaka, S.; Okada, R.; Ohyama, M.; Shoji, M.; et al. Rapid, efficient, and simple motor neuron differentiation from human pluripotent stem cells. Mol. Brain 2015, 8, 79. [Google Scholar] [CrossRef]
- Burden, S.J.; Huijbers, M.G.; Remedio, L. Fundamental Molecules and Mechanisms for Forming and Maintaining Neuromuscular Synapses. Int. J. Mol. Sci. 2018, 19, 490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cetin, H.; Beeson, D.; Vincent, A.; Webster, R. The Structure, Function, and Physiology of the Fetal and Adult Acetylcholine Receptor in Muscle. Front. Mol. Neurosci. 2020, 13, 581097. [Google Scholar] [CrossRef] [PubMed]
- Xing, G.; Xiong, W.C.; Mei, L. Rapsyn as a signaling and scaffolding molecule in neuromuscular junction formation and maintenance. Neurosci. Lett. 2020, 731, 135013. [Google Scholar] [CrossRef] [PubMed]
- Phillips, W.D.; Kopta, C.; Blount, P.; Gardner, P.D.; Steinbach, J.H.; Merlie, J.P. ACh Receptor-Rich Membrane Domains Organized in Fibroblasts by Recombinant 43-Kilodalton Protein. Science 1991, 251, 568–570. [Google Scholar] [CrossRef] [PubMed]
- Mayer, U. Integrins: Redundant or important players in skeletal muscle? J. Biol. Chem. 2003, 278, 14587–14590. [Google Scholar] [CrossRef] [Green Version]
- Burkin, D.J.; Gu, M.; Hodges, B.L.; Campanelli, J.T.; Kaufman, S.J. A Functional Role for Specific Spliced Variants of the α7β1 Integrin in Acetylcholine Receptor Clustering. J. Cell Biol. 1998, 143, 1067–1075. [Google Scholar] [CrossRef] [Green Version]
- Montanaro, F.; Gee, S.H.; Jacobson, C.; Lindenbaum, M.H.; Froehner, S.C.; Carbonetto, S. Laminin and α-Dystroglycan Mediate Acetylcholine Receptor Aggregation via a MuSK-Independent Pathway. J. Neurosci. 1998, 18, 1250–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiyama, J.E.; Glass, D.J.; Yancopoulos, G.D.; Hall, Z.W. Laminin-induced acetylcholine receptor clustering: An alternative pathway. J. Cell Biol. 1997, 139, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Iberite, F.; Gruppioni, E.; Ricotti, L. Skeletal muscle differentiation of human iPSCs meets bioengineering strategies: Perspectives and challenges. NPJ Regen. Med. 2022, 7, 23. [Google Scholar] [CrossRef]
- Brunetti, J.; Koenig, S.; Monnier, A.; Frieden, M. Nanopattern surface improves cultured human myotube maturation. Skelet. Muscle 2021, 11, 12. [Google Scholar] [CrossRef]
- Denes, L.T.; Riley, L.A.; Mijares, J.R.; Arboleda, J.D.; McKee, K.; Esser, K.A.; Wang, E.T. Culturing C2C12 myotubes on micromolded gelatin hydrogels accelerates myotube maturation. Skelet Muscle 2019, 9, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badu-Mensah, A.; Guo, X.; Nimbalkar, S.; Cai, Y.; Hickman, J.J. ALS mutations in both human skeletal muscle and motoneurons differentially affects neuromuscular junction integrity and function. Biomaterials 2022, 289, 121752. [Google Scholar] [CrossRef] [PubMed]
- Machado, C.B.; Pluchon, P.; Harley, P.; Rigby, M.; Sabater, V.G.; Stevenson, D.C.; Hynes, S.; Lowe, A.; Burrone, J.; Viasnoff, V.; et al. In Vitro Modeling of Nerve-Muscle Connectivity in a Compartmentalized Tissue Culture Device. Adv. Biosyst. 2019, 3, 1800307. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Orientation | Sequence (5′-3′) |
|---|---|---|
| Gapdh | Forward | TCAACAGCAACTCCCACTCTTCCA |
| Reverse | ACCACCCTGTTGCTGTACCGTATT | |
| Lrp4 | Forward | GGAGGGTCAGTGAAGATGTAAAG |
| Reverse | CTGGCTGCTGATCTCTGAATAG | |
| Musk | Forward | CATGGCAGAGTTTGACAACCC |
| Reverse | TTCGGAGGAACTCATTGAGGTC | |
| Dok7 | Forward | TGAGCTTCCTGTTTGACTGCA |
| Reverse | GCAACACGCTCTTCTGAGGC | |
| Itgα5 | Forward | ACCTGGACCAAGACGGCTACAA |
| Reverse | CTGGGAAGGTTTAGTGCTCAGTC | |
| Itgα7 | Forward | AACCAATGGCTGGGAGTCAG |
| Reverse | ATCCCGAGTCTCCAAAGCCT | |
| Achrα | Forward | ACCTGGACCTATGACGGCTCT |
| Reverse | AGTTACTCAGGTCGGGCTGGT | |
| Achrγ | Forward | CTTGTGGCTAAGAAGGTGCCTG |
| Reverse | GCAAGGACACATTGAGCACGAC | |
| Achrε | Forward | AGACCTGAGGACACTGTCACCA |
| Reverse | TCGTCCTTGCTGTAGTTGAGCC | |
| MyoD | Forward | AGGACACGACTGCTTTCTTCACCA |
| Reverse | TTAACTTTCTGCCACTCCGGAACC | |
| MyoG | Forward | CCAACCCAGGAGATCATTTG |
| Reverse | ACGATGGACGTAAGGGAGTG | |
| Myh7 | Forward | ATGCTGACAGATCGGGAGAA |
| Reverse | GGTTGGCTTGGATGATTTGA | |
| Myh4 | Forward | GGCACCCTTGAGGATCAAAT |
| Reverse | GCTATCAATGTCCGCAGAGG |
| Total Number of Observations [-] | Number of Observations Including Myotubes Contracted by Glutamate Addition [-] | |
|---|---|---|
| Flat | 12 | 2 |
| Patterned | 12 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimizu, K.; Kassai, H.; Kamei, Y.; Yamamoto, K.; Nagashima, T.; Maekawa, T.; Akiyama, H.; Honda, H. Alignment of Skeletal Muscle Cells Facilitates Acetylcholine Receptor Clustering and Neuromuscular Junction Formation with Co-Cultured Human iPSC-Derived Motor Neurons. Cells 2022, 11, 3760. https://doi.org/10.3390/cells11233760
Shimizu K, Kassai H, Kamei Y, Yamamoto K, Nagashima T, Maekawa T, Akiyama H, Honda H. Alignment of Skeletal Muscle Cells Facilitates Acetylcholine Receptor Clustering and Neuromuscular Junction Formation with Co-Cultured Human iPSC-Derived Motor Neurons. Cells. 2022; 11(23):3760. https://doi.org/10.3390/cells11233760
Chicago/Turabian StyleShimizu, Kazunori, Haruo Kassai, Yuhei Kamei, Kazuki Yamamoto, Takunori Nagashima, Tadayoshi Maekawa, Hirokazu Akiyama, and Hiroyuki Honda. 2022. "Alignment of Skeletal Muscle Cells Facilitates Acetylcholine Receptor Clustering and Neuromuscular Junction Formation with Co-Cultured Human iPSC-Derived Motor Neurons" Cells 11, no. 23: 3760. https://doi.org/10.3390/cells11233760