
Physiological Oxygen Levels Differentially Regulate Adipokine Production in Abdominal and Femoral Adipocytes from Individuals with Obesity Versus Normal Weight
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Upper and Lower Body Adipose Tissue Biopsies
2.2. Human Primary Adipocyte Experiments
2.3. Adipocyte Gene Expression
2.4. Adipokine Secretion
2.5. Statistical Analyses
3. Results
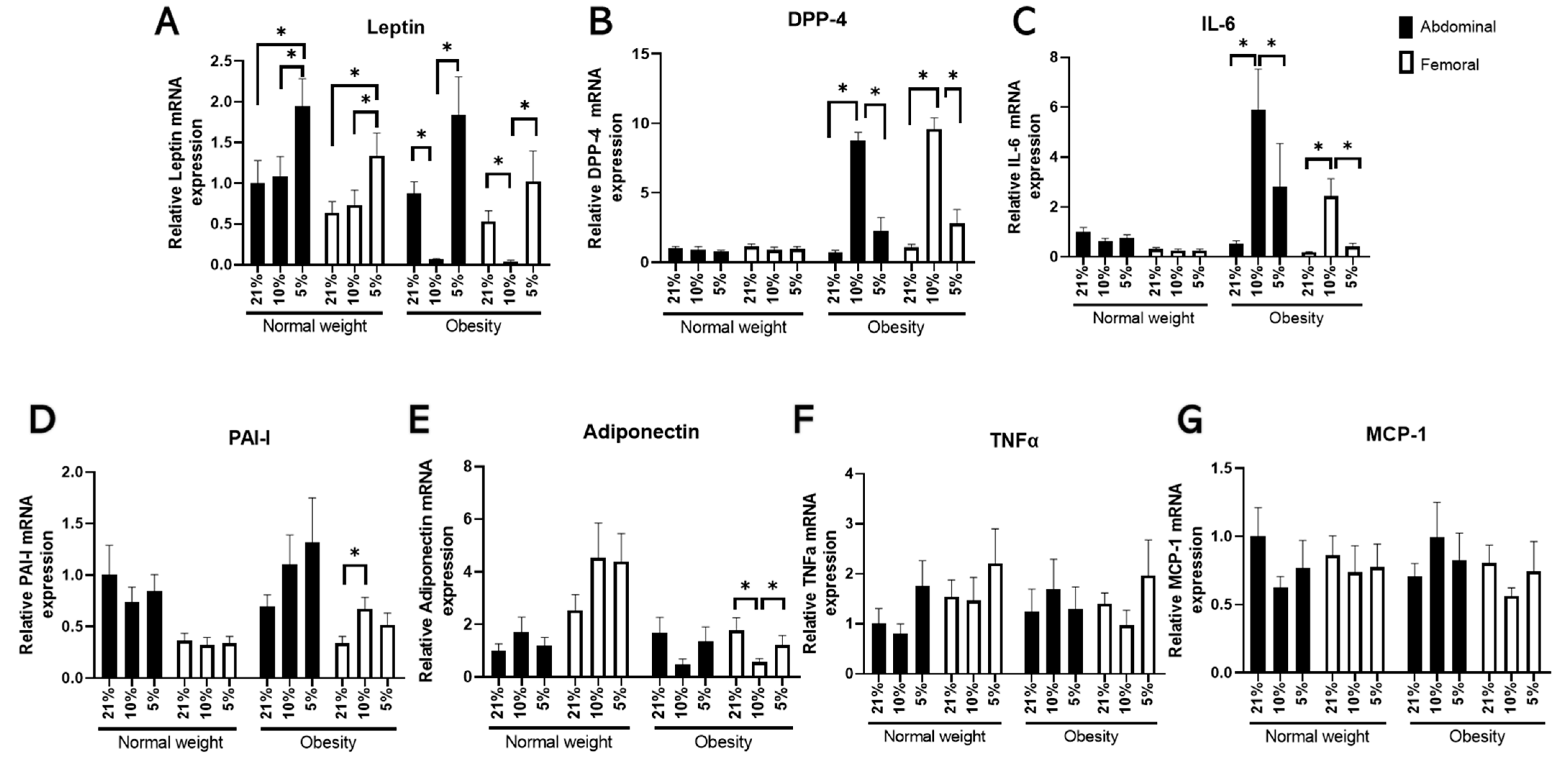
3.1. The Effects of Oxygen Partial Pressure on Adipocyte Gene Expression
3.2. The Effects of Oxygen Partial Pressure on Adipokine Secretion
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kopelman, P.G. Obesity as a medical problem. Nature 2000, 404, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Karpe, F.; Pinnick, K.E. Biology of upper-body and lower-body adipose tissue—Link to whole-body phenotypes. Nat. Rev. Endocrinol. 2014, 11, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Goossens, G.H. The Metabolic Phenotype in Obesity: Fat Mass, Body Fat Distribution, and Adipose Tissue Function. Obes. Facts 2017, 10, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, S.; Hawken, S.; Ôunpuu, S.; Bautista, L.; Franzosi, M.G.; Commerford, P.; Lang, C.C.; Rumboldt, Z.; Onen, C.L.; Lisheng, L.; et al. Obesity and the risk of myocardial infarction in 27 000 participants from 52 countries: A case-control study. Lancet 2005, 366, 1640–1649. [Google Scholar] [CrossRef]
- Seidell, J.; Perusse, L.; Després, J.-P.; Bouchard, C. Waist and hip circumferences have independent and opposite effects on cardiovascular disease risk factors: The Quebec Family Study. Am. J. Clin. Nutr. 2001, 74, 315–321. [Google Scholar] [CrossRef] [Green Version]
- Goossens, G.H.; Jocken, J.W.; Blaak, E.E. Sexual Dimorphism in Cardiometabolic Health: The Role of Adipose Tissue, Muscle and Liver. Nat. Rev. Endocrinol. 2021, 17, 47–66. [Google Scholar] [CrossRef]
- Goossens, G.H. The role of adipose tissue dysfunction in the pathogenesis of obesity-related insulin resistance. Physiol. Behav. 2008, 94, 206–218. [Google Scholar] [CrossRef]
- Rosen, E.D.; Spiegelman, B.M. What We Talk About When We Talk About Fat. Cell 2014, 156, 20–44. [Google Scholar] [CrossRef] [Green Version]
- Lempesis, I.G.; Van Meijel, R.L.J.; Manolopoulos, K.N.; Goossens, G.H. Oxygenation of adipose tissue: A human perspective. Acta Physiol. 2019, 228, e13298. [Google Scholar] [CrossRef]
- Heinonen, S.; Jokinen, R.; Rissanen, A.; Pietiläinen, K.H. White adipose tissue mitochondrial metabolism in health and in obesity. Obes. Rev. 2019, 21, e12958. [Google Scholar] [CrossRef]
- Chen, K.; Zhang, J.; Beeraka, N.M.; Tang, C.; Babayeva, Y.V.; Sinelnikov, M.Y.; Zhang, X.; Zhang, J.; Liu, J.; Reshetov, I.V.; et al. Advances in the Prevention and Treatment of Obesity-Driven Effects in Breast Cancers. Front. Oncol. 2022, 12, 2663. [Google Scholar] [CrossRef] [PubMed]
- Goossens, G.H.; Blaak, E.E. Adipose Tissue Dysfunction and Impaired Metabolic Health in Human Obesity: A Matter of Oxygen? Front. Endocrinol. 2015, 6, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goossens, G.H.; Blaak, E.E. Adipose Tissue Oxygen Tension: Implications for Chronic Metabolic and Inflammatory Diseases. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 539–546. [Google Scholar] [CrossRef]
- Trayhurn, P. Hypoxia and Adipose Tissue Function and Dysfunction in Obesity. Physiol. Rev. 2013, 93, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lempesis, I.G.; Goossens, G.H.; Manolopoulos, K.N. Measurement of human abdominal and femoral intravascular adipose tissue blood flow using percutaneous Doppler ultrasound. Adipocyte 2021, 10, 119–123. [Google Scholar] [CrossRef]
- Vogel, M.A.; Jocken, J.W.; Sell, H.; Hoebers, N.; Essers, Y.; Rouschop, K.M.; Čajlaković, M.; Blaak, E.E.; Goossens, G.H. Differences in Upper and Lower-Body Adipose Tissue Oxygen Tension Contribute to the Adipose Tissue Phenotype in Humans. J. Clin. Endocrinol. Metab. 2018, 103, 3688–3697. [Google Scholar] [CrossRef] [Green Version]
- Rausch, M.E.; Weisberg, S.; Vardhana, P.; Tortoriello, D.V. Obesity in C57BL/6J mice is characterized by adipose tissue hypoxia and cytotoxic T-cell infiltration. Int. J. Obes. 2007, 32, 451–463. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Gao, Z.; Yin, J.; He, Q. Hypoxia Is a Potential Risk Factor for Chronic Inflammation and Adiponectin Reduction in Adipose Tissue of Ob/Ob and Dietary Obese Mice. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1118–E1128. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Gao, Z.; He, Q.; Zhou, D.; Guo, Z.; Ye, J. Role of hypoxia in obesity-induced disorders of glucose and lipid metabolism in adipose tissue. Am. J. Physiol. Metab. 2009, 296, E333–E342. [Google Scholar] [CrossRef] [Green Version]
- Hodson, L.; Humphreys, S.M.; Karpe, F.; Frayn, K.N. Metabolic Signatures of Human Adipose Tissue Hypoxia in Obesity. Diabetes 2013, 62, 1417–1425. [Google Scholar] [CrossRef]
- Pasarica, M.; Sereda, O.R.; Redman, L.M.; Albarado, D.C.; Hymel, D.T.; Roan, L.E.; Rood, J.C.; Burk, D.H.; Smith, S.R. Reduced Adipose Tissue Oxygenation in Human Obesity: Evidence for Rarefaction, Macrophage Chemotaxis, and Inflammation without an Angiogenic Response. Diabetes 2009, 58, 718–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goossens, G.H.; Bizzarri, A.; Venteclef, N.; Essers, Y.; Cleutjens, J.P.; Konings, E.; Jocken, J.W.E.; Čajlaković, M.; Ribitsch, V.; Clément, K.; et al. Increased Adipose Tissue Oxygen Tension in Obese Compared with Lean Men Is Accompanied by Insulin Resistance, Impaired Adipose Tissue Capillarization, and Inflammation. Circulation 2011, 124, 67–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawler, H.M.; Underkofler, C.M.; Kern, P.A.; Erickson, C.; Bredbeck, B.; Rasouli, N. Adipose Tissue Hypoxia, Inflammation, and Fibrosis in Obese Insulin-Sensitive and Obese Insulin-Resistant Subjects. J. Clin. Endocrinol. Metab. 2016, 101, 1422–1428. [Google Scholar] [CrossRef] [Green Version]
- Cifarelli, V.; Beeman, S.C.; Smith, G.I.; Yoshino, J.; Morozov, D.; Beals, J.W.; Kayser, B.D.; Watrous, J.D.; Jain, M.; Patterson, B.W.; et al. Decreased adipose tissue oxygenation associates with insulin resistance in individuals with obesity. J. Clin. Investig. 2020, 130, 6688–6699. [Google Scholar] [CrossRef] [PubMed]
- Goossens, G.H.; Vogel, M.A.A.; Vink, R.G.; Mariman, E.C.; Van Baak, M.A.; Blaak, E.E. Adipose tissue oxygenation is associated with insulin sensitivity independently of adiposity in obese men and women. Diabetes Obes. Metab. 2018, 20, 2286–2290. [Google Scholar] [CrossRef] [PubMed]
- Trayhurn, P. Hypoxia and Adipocyte Physiology: Implications for Adipose Tissue Dysfunction in Obesity. Annu. Rev. Nutr. 2014, 34, 207–236. [Google Scholar] [CrossRef] [PubMed]
- Famulla, S.; Schlich, R.; Sell, H.; Eckel, J. Differentiation of Human Adipocytes at Physiological Oxygen Levels Results in Increased Adiponectin Secretion and Isoproterenol-Stimulated Lipolysis. Adipocyte 2012, 1, 132–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jocken, J.W.E.; Goossens, G.H.; Popeijus, H.; Essers, Y.; Hoebers, N.; Blaak, E.E. Contribution of lipase deficiency to mitochondrial dysfunction and insulin resistance in hMADS adipocytes. Int. J. Obes. 2015, 40, 507–513. [Google Scholar] [CrossRef]
- Rouschop, K.M.; Ramaekers, C.H.; Schaaf, M.B.; Keulers, T.G.; Savelkouls, K.G.; Lambin, P.; Koritzinsky, M.; Wouters, B. Autophagy is required during cycling hypoxia to lower production of reactive oxygen species. Radiother. Oncol. 2009, 92, 411–416. [Google Scholar] [CrossRef]
- Dardeno, T.A.; Chou, S.H.; Moon, H.-S.; Chamberland, J.P.; Fiorenza, C.G.; Mantzoros, C.S. Leptin in human physiology and therapeutics. Front. Neuroendocr. 2010, 31, 377–393. [Google Scholar] [CrossRef]
- Fasshauer, M.; Blüher, M. Adipokines in health and disease. Trends Pharmacol. Sci. 2015, 36, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Celik, O.; Aydin, S.; Celik, N.; Yilmaz, M. Peptides: Basic determinants of reproductive functions. Peptides 2015, 72, 34–43. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Parameter | Normal Weight (n = 9) | Obesity (n = 9) | p Value |
|---|---|---|---|
| Age (years) | 56.7 ± 1.8 | 56 ± 1.3 | 0.566 |
| BMI (kg/m2) | 22.8 ± 0.4 | 34.8 ± 1.3 | <0.001 |
| Waist circumference (cm) | 79.4 ± 3.1 | 105.2 ± 3.8 | <0.001 |
| Hip circumference (cm) | 94.4 ± 2.8 | 119.9 ± 4.8 | <0.001 |
| Waist-to-hip ratio | 0.84 ± 0.02 | 0.88 ± 0.04 | 0.127 |
| Visceral fat mass (g) | 402.5 ± 118 | 1,325 ± 153.3 | 0.003 |
| Abdominal fat mass (kg) | 10.01 ± 1.48 | 24.4 ± 2.37 | <0.001 |
| Leg fat mass (kg) | 7.67 ± 0.86 | 16.03 ± 1.43 | 0.001 |
| Fasting glucose (mmol/L) | 4.91 ± 0.10 | 5.10 ± 0.23 | 0.416 |
| 2-hour glucose (mmol/L) | 4.90 ± 0.34 | 4.70 ± 0.33 | 0.684 |
| Fasting insulin (pmol/L) | 28.40 ± 5.80 | 43.30 ± 10.20 | 0.202 |
| HOMA2 IR | 0.46 ± 0.10 | 0.72 ± 0.20 | 0.187 |
| SBP (mmHg) | 119.6 ± 4.4 | 133.0 ± 3.1 | 0.039 |
| DBP (mmHg) | 73.6 ± 4.3 | 81.7 ± 1.9 | 0.153 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lempesis, I.G.; Hoebers, N.; Essers, Y.; Jocken, J.W.E.; Rouschop, K.M.A.; Blaak, E.E.; Manolopoulos, K.N.; Goossens, G.H. Physiological Oxygen Levels Differentially Regulate Adipokine Production in Abdominal and Femoral Adipocytes from Individuals with Obesity Versus Normal Weight. Cells 2022, 11, 3532. https://doi.org/10.3390/cells11223532
Lempesis IG, Hoebers N, Essers Y, Jocken JWE, Rouschop KMA, Blaak EE, Manolopoulos KN, Goossens GH. Physiological Oxygen Levels Differentially Regulate Adipokine Production in Abdominal and Femoral Adipocytes from Individuals with Obesity Versus Normal Weight. Cells. 2022; 11(22):3532. https://doi.org/10.3390/cells11223532
Chicago/Turabian StyleLempesis, Ioannis G., Nicole Hoebers, Yvonne Essers, Johan W. E. Jocken, Kasper M. A. Rouschop, Ellen E. Blaak, Konstantinos N. Manolopoulos, and Gijs H. Goossens. 2022. "Physiological Oxygen Levels Differentially Regulate Adipokine Production in Abdominal and Femoral Adipocytes from Individuals with Obesity Versus Normal Weight" Cells 11, no. 22: 3532. https://doi.org/10.3390/cells11223532