

Bta-miR-199a-3p Inhibits LPS-Induced Inflammation in Bovine Mammary Epithelial Cells via the PI3K/AKT/NF-κB Signaling Pathway


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Inflammation Induction of MAC-T Cells
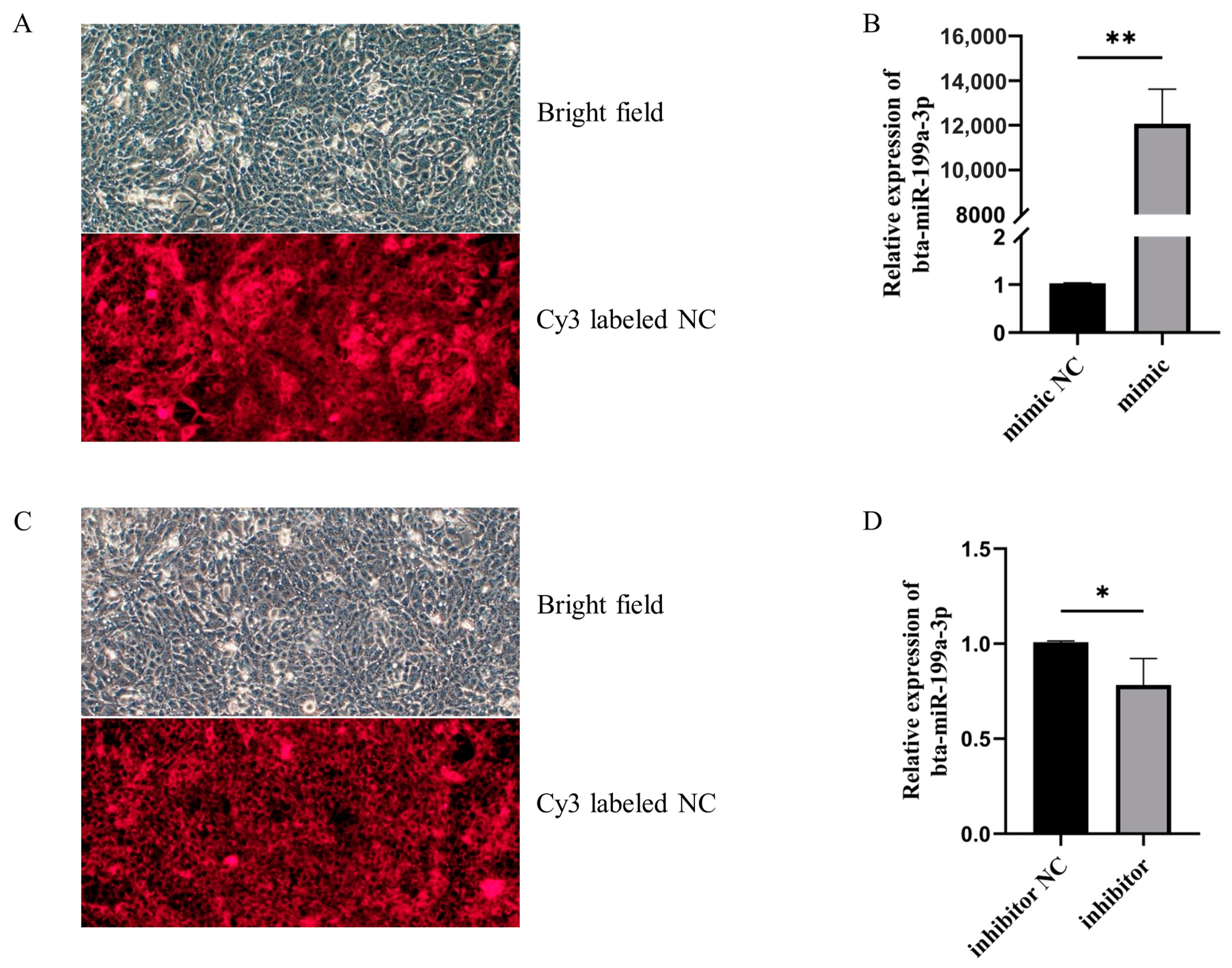
2.3. MAC-T Cell Transfection
2.4. Target Gene Prediction
2.5. Construction of Recombinant Plasmid of Double Luciferase Reporter Gene and Identification of Target Gene of Bta-miR-199a-3p
2.6. RNA Extraction and qRT-PCR
2.7. Western Blot
2.8. ELISA
2.9. CCK-8 Assay
2.10. Statistical Analysis
3. Results
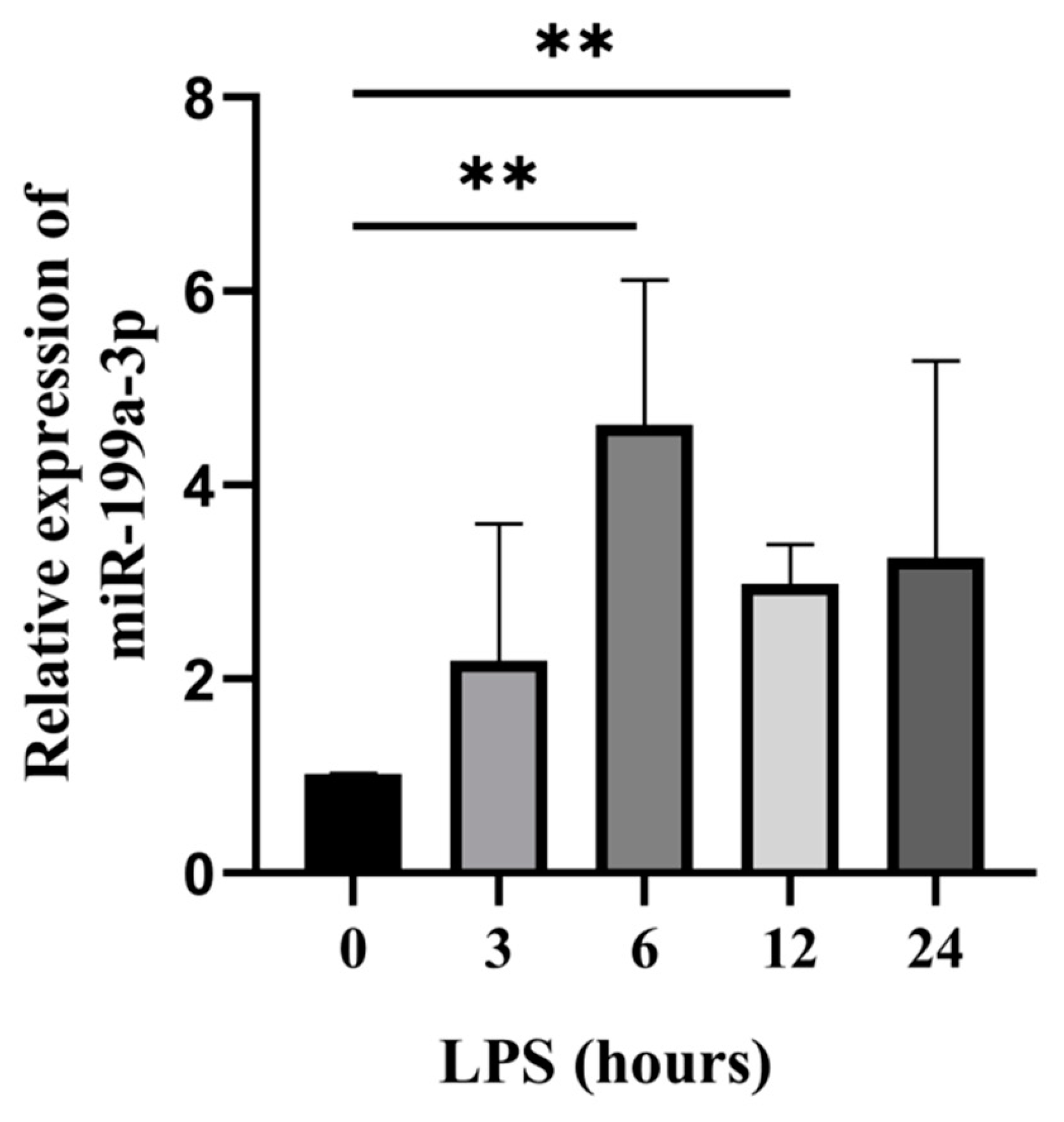
3.1. Bta-miR-199a-3p Was Up-Regulated in LPS-Induced MAC-T Cells
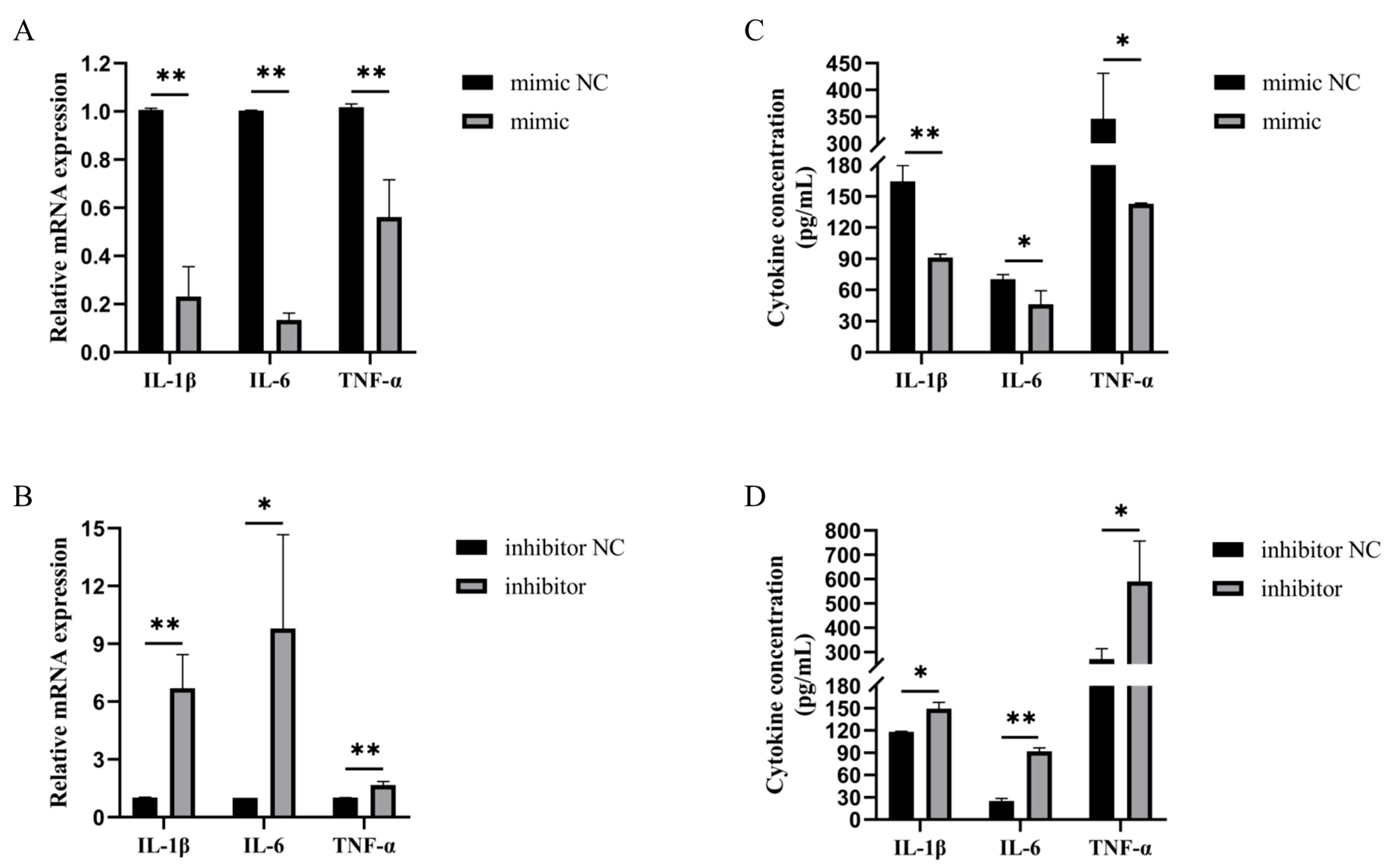
3.2. Bta-miR-199a-3p Inhibited the Inflammatory Response of LPS-Induced MAC-T Cells
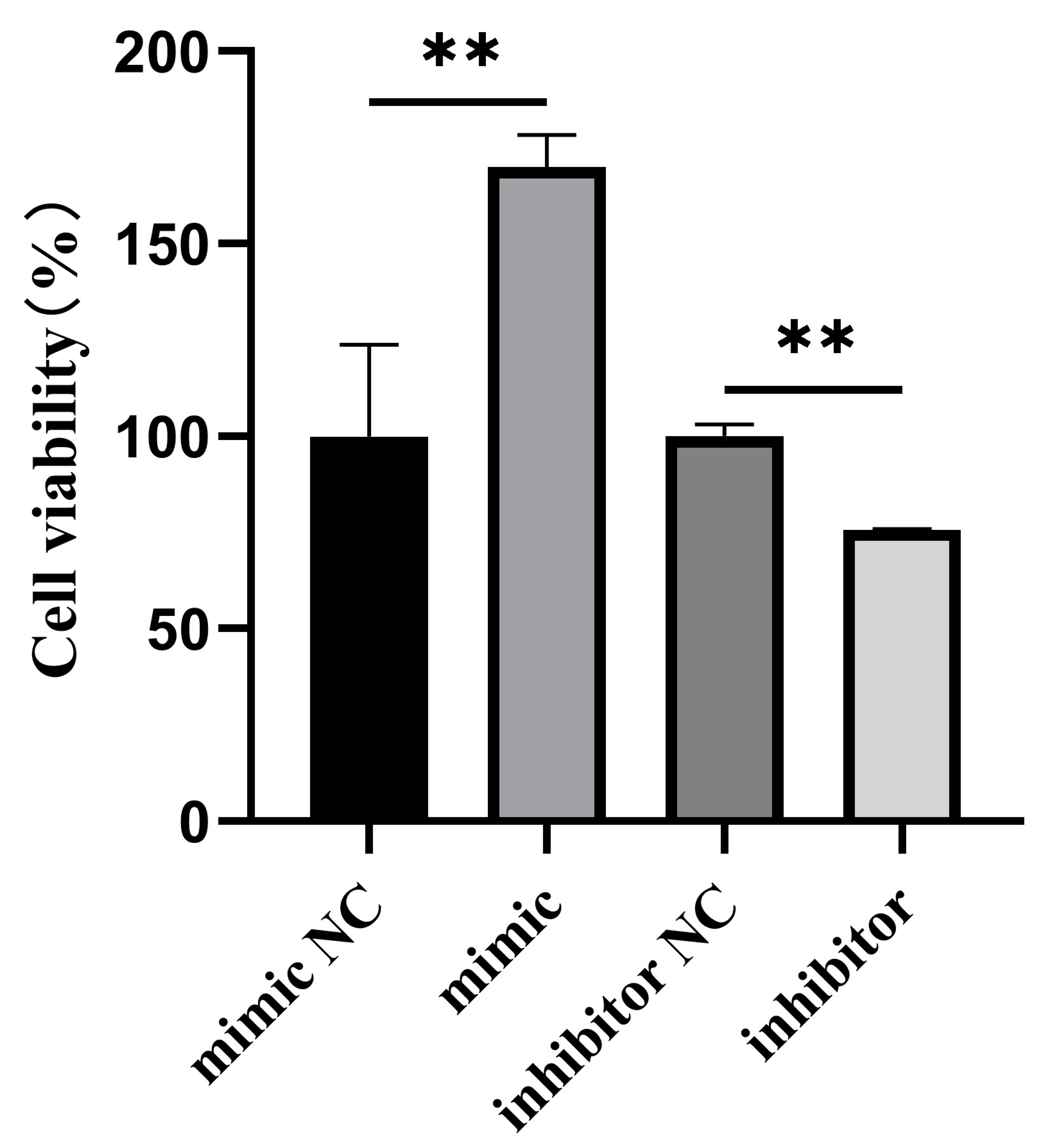
3.3. Effect of Bta-miR-199a-3p on the Proliferation of MAC-T Cells
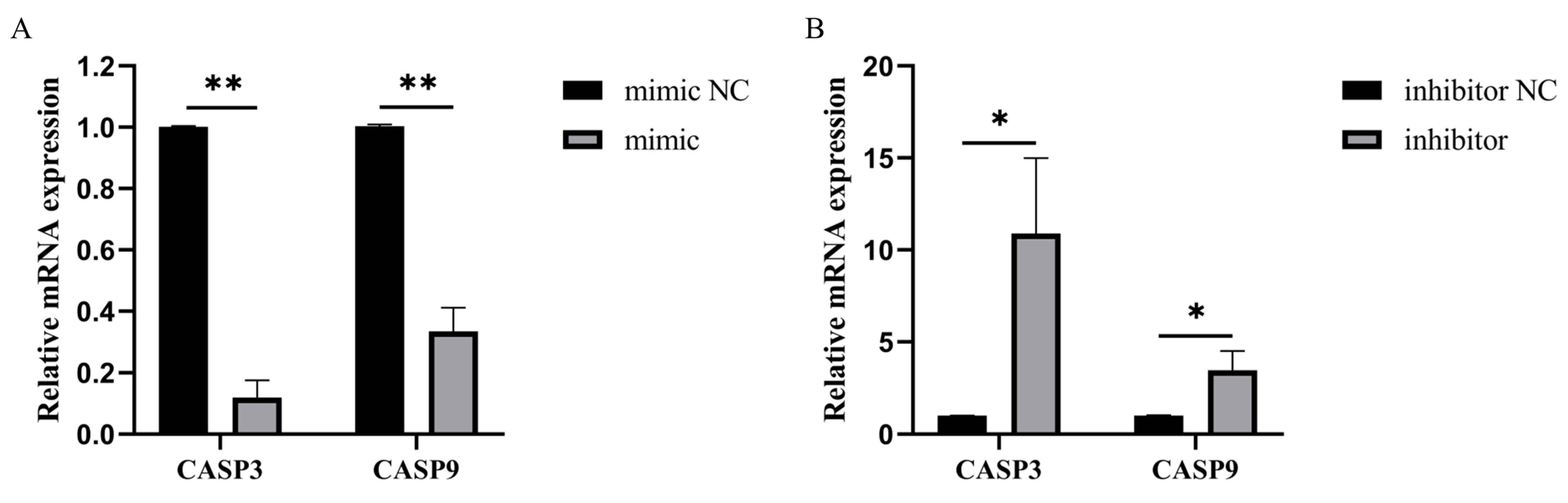
3.4. Bta-miR-199a-3p Inhibited the Expression of Apoptotic Genes
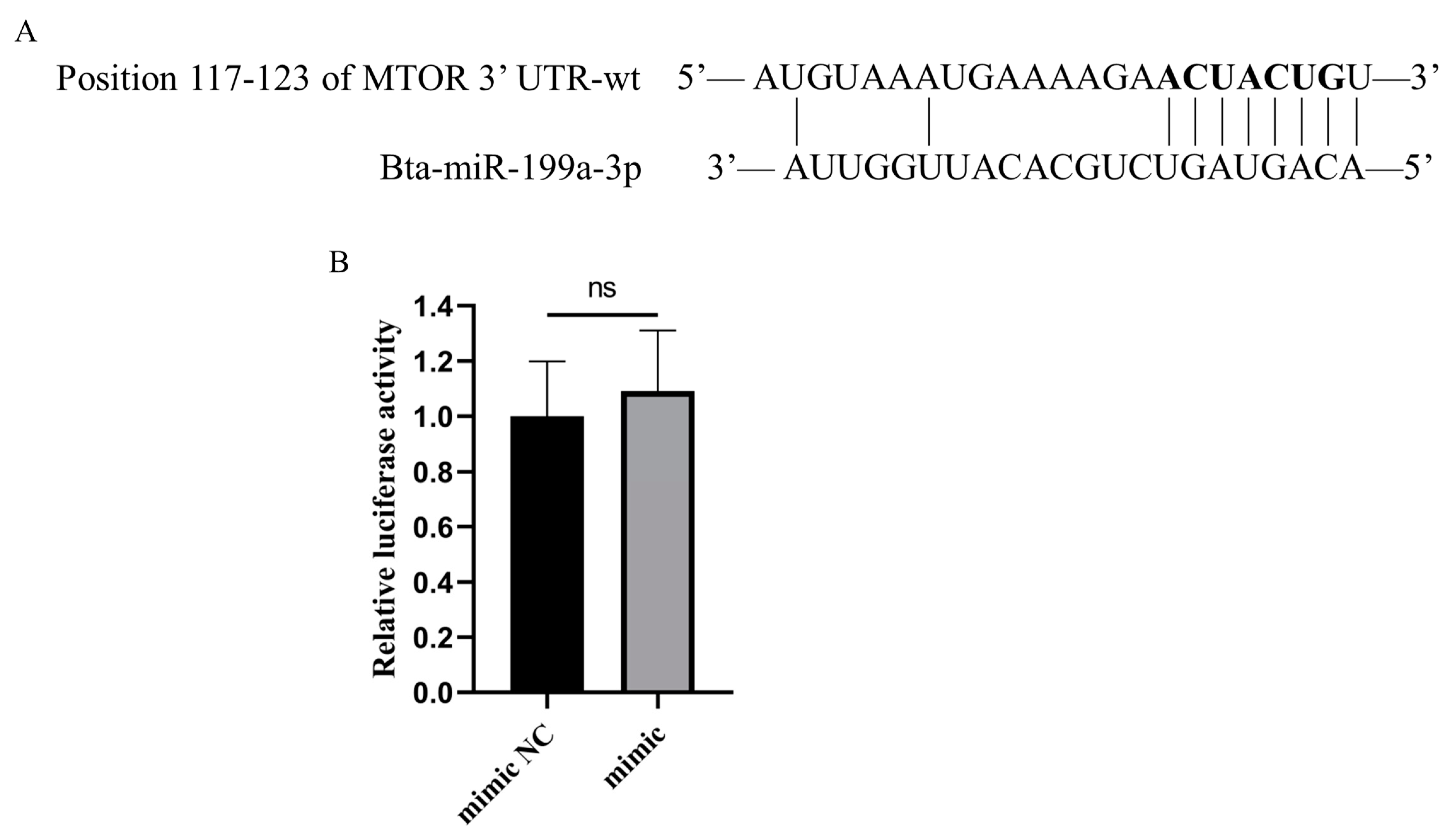
3.5. Bta-miR-199a-3p Does Not Directly Target MTOR
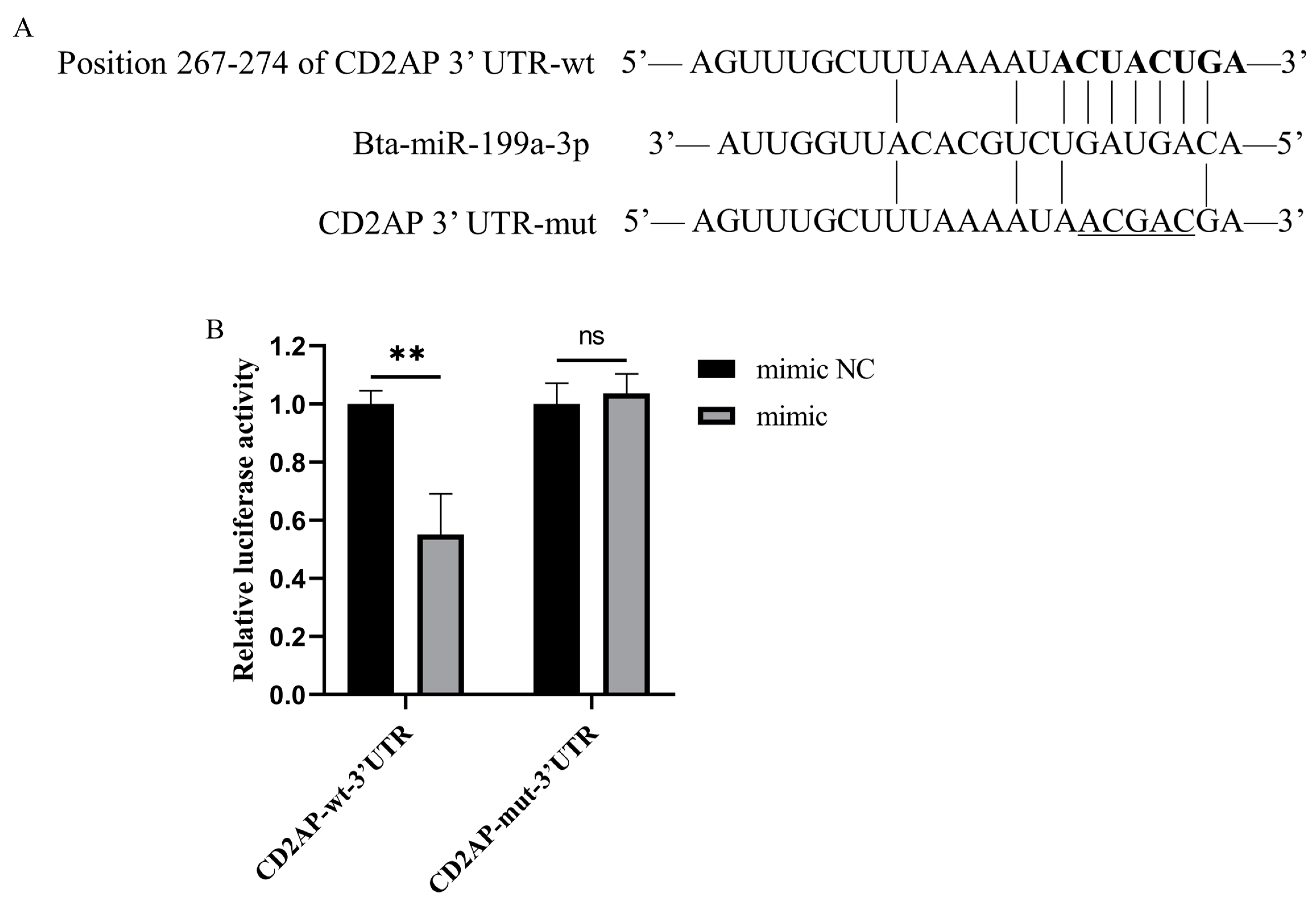
3.6. CD2AP Is a Direct Molecular Target of Bta-miR-199a-3p
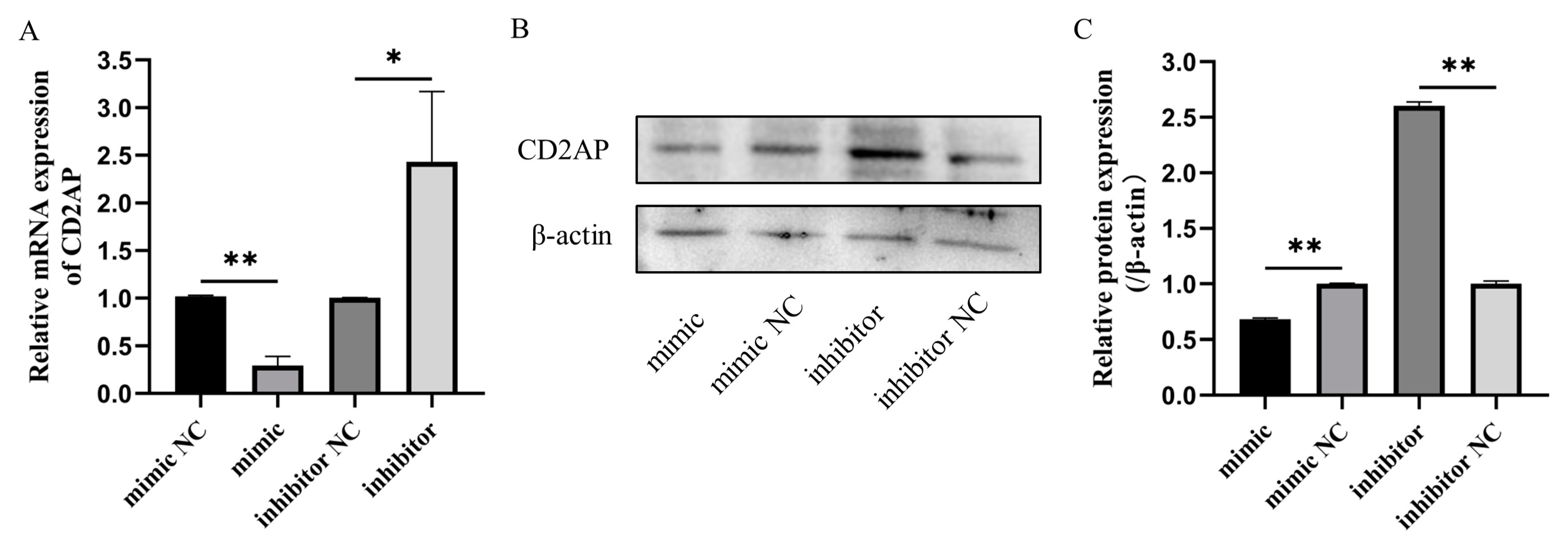
3.7. Bta-miR-199a-3p Negatively Regulates the Expression of CD2AP mRNA and Protein in MAC-T Cells
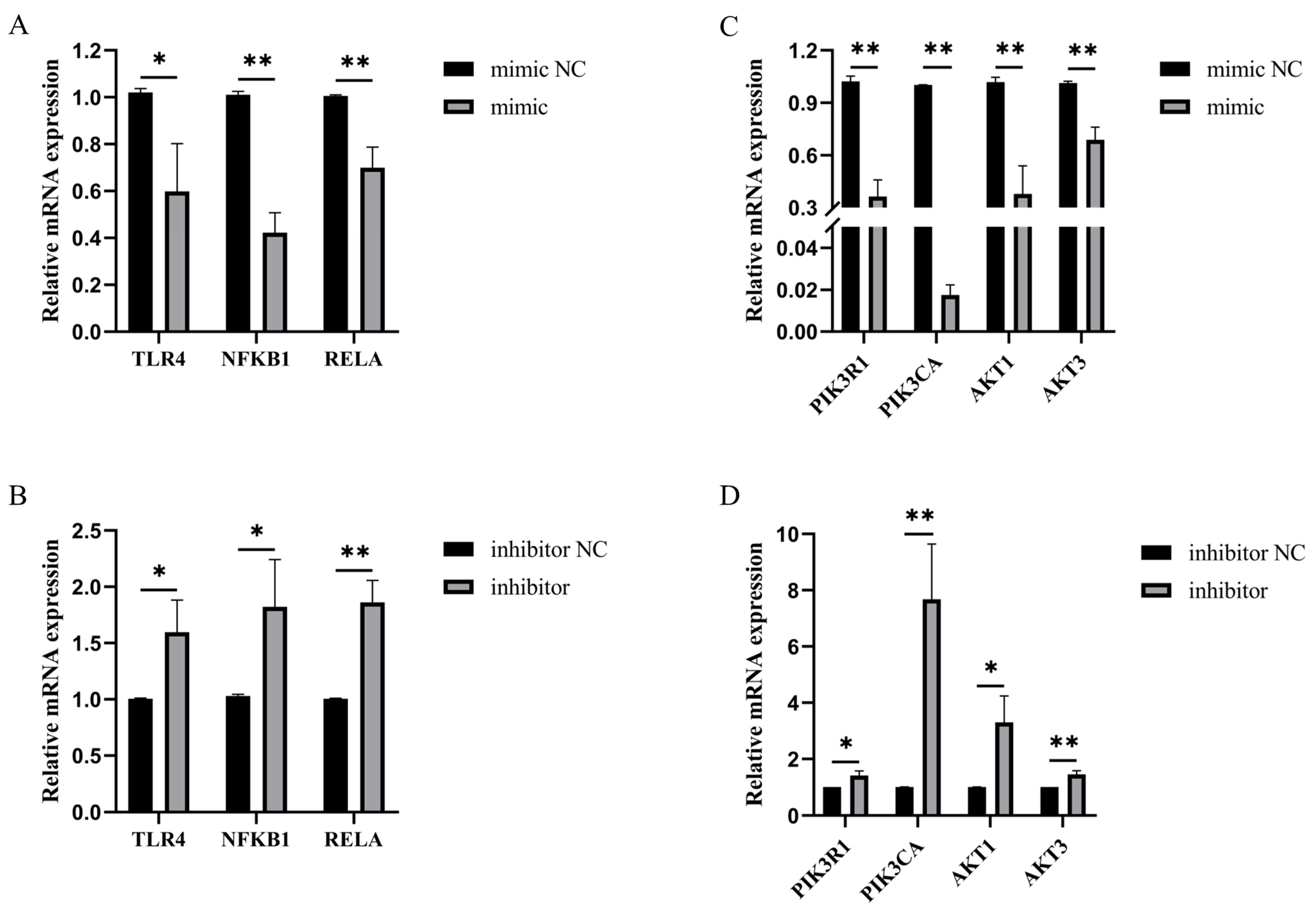
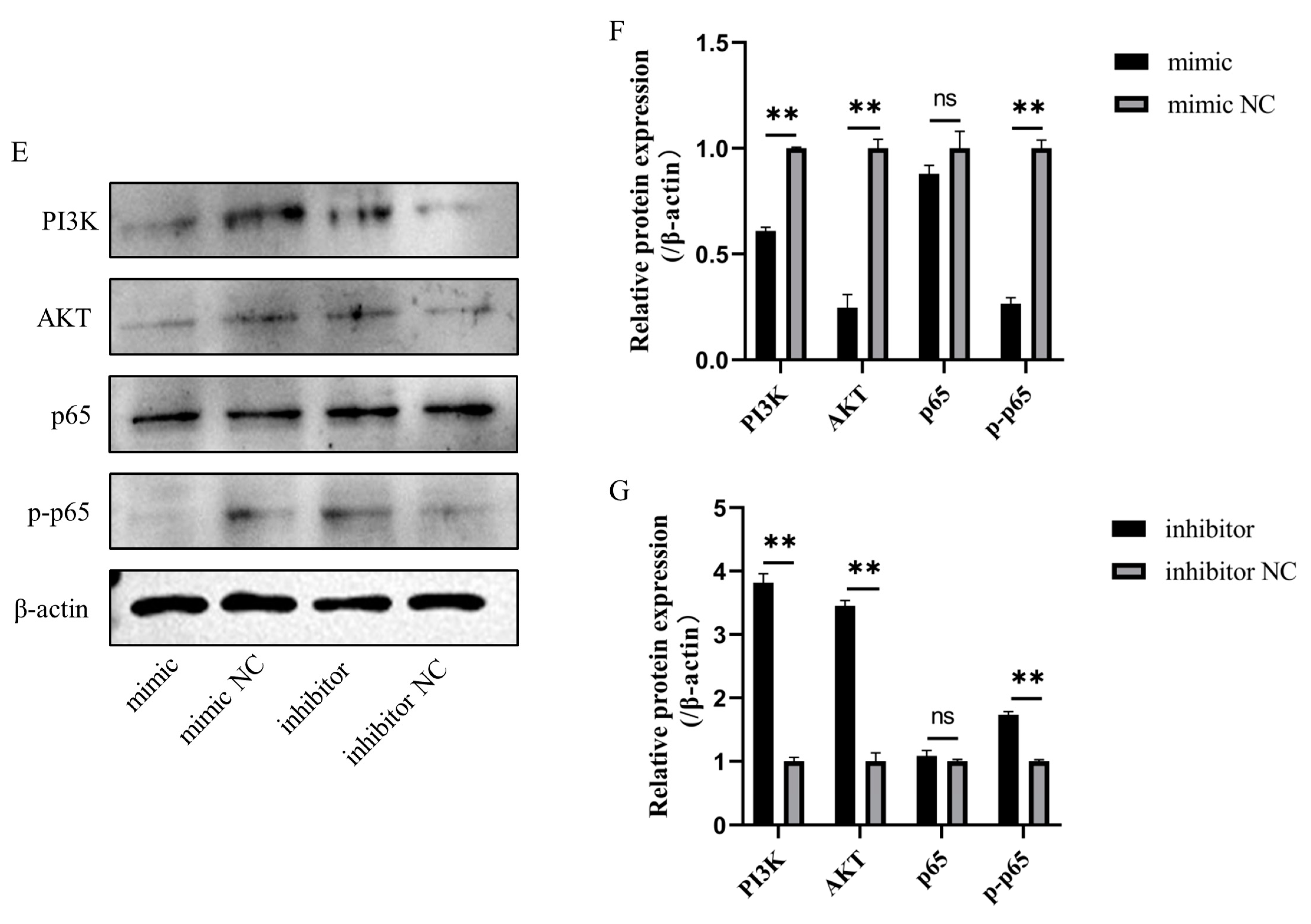
3.8. Bta-miR-199a-3p Inhibited LPS-Induced Activation of the PI3K/AKT/NF-κB Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, H.; Weersink, A.; Kelton, D.; von Massow, M. Estimating milk loss based on somatic cell count at the cow and herd level. J. Dairy Sci. 2021, 104, 7919–7931. [Google Scholar] [CrossRef] [PubMed]
- Dalanezi, F.M.; Joaquim, S.F.; Guimarães, F.F.; Guerra, S.T.; Lopes, B.C.; Schmidt, E.M.S.; Cerri, R.L.A.; Langoni, H. Influence of pathogens causing clinical mastitis on reproductive variables of dairy cows. J. Dairy Sci. 2020, 103, 3648–3655. [Google Scholar] [CrossRef] [PubMed]
- Zigo, F.; Vasil’, M.; Ondrašovičová, S.; Výrostková, J.; Bujok, J.; Pecka-Kielb, E. Maintaining optimal mammary gland health and prevention of mastitis. Front. Vet. Sci. 2021, 8, 607311. [Google Scholar] [CrossRef]
- Brand, B.; Hartmann, A.; Repsilber, D.; Griesbeck-Zilch, B.; Wellnitz, O.; Kuhn, C.; Ponsuksili, S.; Meyer, H.H.; Schwerin, M. Comparative expression profiling of E. coli and S. aureus inoculated primary mammary gland cells sampled from cows with different genetic predispositions for somatic cell score. Genet. Sel. Evol. 2011, 43, 24. [Google Scholar] [CrossRef] [Green Version]
- Bakhtiarizadeh, M.R.; Mirzaei, S.; Norouzi, M.; Sheybani, N.; Vafaei Sadi, M.S. Identification of gene modules and hub genes involved in mastitis development using a systems biology approach. Front. Genet. 2020, 11, 722. [Google Scholar] [CrossRef] [PubMed]
- Naserkheil, M.; Ghafouri, F.; Zakizadeh, S.; Pirany, N.; Manzari, Z.; Ghorbani, S.; Banabazi, M.H.; Bakhtiarizadeh, M.R.; Huq, M.A.; Park, M.N.; et al. Multi-omics integration and network analysis reveal potential hub genes and genetic mechanisms regulating bovine mastitis. Curr. Issues Mol. Biol. 2022, 44, 309–328. [Google Scholar] [CrossRef]
- Cobirka, M.; Tancin, V.; Slama, P. Epidemiology and classification of mastitis. Animals 2020, 10, 2212. [Google Scholar] [CrossRef]
- Wu, Y.; Sun, Y.; Dong, X.; Chen, J.; Wang, Z.; Chen, J.; Dong, G. The synergism of PGN, LTA and LPS in inducing transcriptome changes, inflammatory responses and a decrease in lactation as well as the associated epigenetic mechanisms in bovine mammary epithelial cells. Toxins 2020, 12, 387. [Google Scholar] [CrossRef]
- Fu, Y.; Zhou, E.; Liu, Z.; Li, F.; Liang, D.; Liu, B.; Song, X.; Zhao, F.; Fen, X.; Li, D.; et al. Staphylococcus aureus and Escherichia coli elicit different innate immune responses from bovine mammary epithelial cells. Vet. Immunol. Immunop. 2013, 155, 245–252. [Google Scholar] [CrossRef]
- Chen, L.; Liu, X.; Li, Z.; Wang, J.; Tian, R.; Zhang, H. Integrated analysis of transcriptome mRNA and miRNA profiles reveals self-protective mechanism of bovine MECs induced by LPS. Front. Vet. Sci. 2022, 9, 890043. [Google Scholar] [CrossRef]
- Huynh, H.T.; Robitaille, G.; Turner, J.D. Establishment of bovine mammary epithelial cells (MAC-T): An in vitro model for bovine lactation. Exp. Cell Res. 1991, 197, 191. [Google Scholar] [CrossRef]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy Clin. Immun. 2018, 141, 1202–1207. [Google Scholar] [CrossRef] [Green Version]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Luoreng, Z.; Wang, X.; Mei, C.; Zan, L. Expression profiling of peripheral blood miRNA using RNAseq technology in dairy cows with Escherichia coli-induced mastitis. Sci. Rep. 2018, 8, 12693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Mou, Y.; Wang, Y.; Wang, H.; Mou, X. MiR-199a-3p targets ETNK1 to promote invasion and migration in gastric cancer cells and is associated with poor prognosis. Pathol. Res. Pract. 2019, 215, 152511. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhou, Y.; Zhang, L.; Jia, K.; Wang, S.; Wang, M.; Li, N.; Yu, Y.; Cao, X.; Hou, J. MicroRNA-199a-3p inhibits hepatic apoptosis and hepatocarcinogenesis by targeting PDCD4. Oncogenesis 2020, 9, 95. [Google Scholar] [CrossRef]
- Peng, J.; Jiang, J.; Wang, H.; Feng, X.; Dong, X. MiR-199a-3p suppresses cervical epithelial cell inflammation by inhibiting the HMGB1/TLR4/NF-κB pathway in preterm birth. Mol. Med. Rep. 2020, 22, 926–938. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, F.; Li, W. Function of intestinal barrier protected by regulating the miR-199a-3p in ulcerative colitis: Modulation of IL-23/IL-17A axis. Fund. Clin. Pharmacol. 2021, 35, 852–860. [Google Scholar] [CrossRef]
- Bardin, P.; Marchal-Duval, E.; Sonneville, F.; Blouquit-Laye, S.; Rousselet, N.; Le Rouzic, P.; Corvol, H.; Tabary, O. Small RNA and transcriptome sequencing reveal the role of miR-199a-3p in inflammatory processes in cystic fibrosis airways. J. Pathol. 2018, 245, 410–420. [Google Scholar] [CrossRef]
- Yang, J.; Hu, Q.; Wang, J.; Ren, Q.; Wang, X.; Luoreng, Z.; Wei, D.; Ma, Y. RNA-Seq reveals the role of miR-29c in regulating inflammation and oxidative stress of bovine mammary epithelial cells. Front. Vet. Sci. 2022, 9, 865415. [Google Scholar] [CrossRef]
- Strandberg, Y.; Gray, C.; Vuocolo, T.; Donaldson, L.; Broadway, M.; Tellam, R. Lipopolysaccharide and lipoteichoic acid induce different innate immune responses in bovine mammary epithelial cells. Cytokine 2005, 31, 72–86. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bougarn, S.; Cunha, P.; Gilbert, F.B.; Meurens, F.; Rainard, P. Technical note: Validation of candidate reference genes for normalization of quantitative PCR in bovine mammary epithelial cells responding to inflammatory stimuli. J. Dairy Sci. 2011, 94, 2425–2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Fornari, F.; Milazzo, M.; Chieco, P.; Negrini, M.; Calin, G.A.; Grazi, G.L.; Pollutri, D.; Croce, C.M.; Bolondi, L.; Gramantieri, L. MiR-199a-3p regulates mTOR and c-Met to influence the doxorubicin sensitivity of human hepatocarcinoma cells. Cancer Res. 2010, 70, 5184–5193. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Huang, H.; He, C.; Wang, K. MicroRNA-199a-3p regulates endometrial cancer cell proliferation by targeting mammalian target of rapamycin (mTOR). Int. J. Gynecol. Cancer 2013, 23, 1191–1197. [Google Scholar] [CrossRef]
- Zhao, X.; Lacasse, P. Mammary tissue damage during bovine mastitis: Causes and control. J. Anim. Sci. 2008, 86, 57–65. [Google Scholar] [CrossRef]
- Sharun, K.; Dhama, K.; Tiwari, R.; Gugjoo, M.B.; Iqbal, Y.M.; Patel, S.K.; Pathak, M.; Karthik, K.; Khurana, S.K.; Singh, R.; et al. Advances in therapeutic and managemental approaches of bovine mastitis: A comprehensive review. Vet. Q. 2021, 41, 107–136. [Google Scholar] [CrossRef]
- Singh, R.P.; Massachi, I.; Manickavel, S.; Singh, S.; Rao, N.P.; Hasan, S.; Mc Curdy, D.K.; Sharma, S.; Wong, D.; Hahn, B.H.; et al. The role of miRNA in inflammation and autoimmunity. Autoimmun Rev. 2013, 12, 1160–1165. [Google Scholar] [CrossRef]
- Luoreng, Z.; Wang, X.; Mei, C.; Zan, L. Comparison of microRNA profiles between bovine mammary glands infected with Staphylococcus aureus and Escherichia coli. Int. J. Biol. Sci. 2018, 14, 87–99. [Google Scholar] [CrossRef]
- Vidya, M.K.; Kumar, V.G.; Sejian, V.; Bagath, M.; Krishnan, G.; Bhatta, R. Toll-like receptors: Significance, ligands, signaling pathways, and functions in mammals. Int. Rev. Immunol. 2018, 37, 20–36. [Google Scholar] [CrossRef] [PubMed]
- Dolasia, K.; Bisht, M.K.; Pradhan, G.; Udgata, A.; Mukhopadhyay, S. TLRs/NLRs: Shaping the landscape of host immunity. Int. Rev. Immunol. 2018, 37, 3–19. [Google Scholar] [CrossRef]
- Ciesielska, A.; Matyjek, M.; Kwiatkowska, K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell Mol. Life Sci. 2021, 78, 1233–1261. [Google Scholar] [CrossRef]
- Zhou, L.; Zhang, S.; Zhang, L.; Li, F.; Sun, H.; Feng, J. MiR-199a-3p inhibits the proliferation, migration, and invasion of endothelial cells and retinal pericytes of diabetic retinopathy rats through regulating FGF7 via EGFR/PI3K/AKT pathway. J. Recept Sig. Transd. 2021, 41, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, Z.; Tang, Q. Reduced expression of microRNA-199a-3p is associated with vascular endothelial cell injury induced by type 2 diabetes mellitus. Exp. Ther. Med. 2018, 16, 3639–3645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dustin, M.L.; Olszowy, M.W.; Holdorf, A.D.; Li, J.; Bromley, S.; Desai, N.; Widder, P.; Rosenberger, F.; van der Merwe, P.A.; Allen, P.M.; et al. A novel adaptor protein orchestrates receptor patterning and cytoskeletal polarity in T-cell contacts. Cell 1998, 94, 667–677. [Google Scholar] [CrossRef] [Green Version]
- Tao, Q.; Chen, Y.; Wu, Z. The role of CD2AP in the pathogenesis of Alzheimer’s disease. Aging Dis. 2019, 10, 901. [Google Scholar] [CrossRef] [Green Version]
- Wolf, G.; Stahl, R.A. CD2-associated protein and glomerular disease. Lancet 2003, 362, 1746–1748. [Google Scholar] [CrossRef]
- Huber, T.B.; Hartleben, B.; Kim, J.; Schmidts, M.; Schermer, B.; Keil, A.; Egger, L.; Lecha, R.L.; Borner, C.; Pavenstadt, H.; et al. Nephrin and CD2AP associate with phosphoinositide 3-OH kinase and stimulate AKT-dependent signaling. Mol. Cell Biol. 2003, 23, 4917–4928. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, C.; Tang, H.; Gao, S.; Sun, F.; Yang, Y.; Zhou, W.; Hu, Y.; Ke, C.; Wu, Y.; et al. CD2-associated protein contributes to hepatitis C, virus propagation and steatosis by disrupting insulin signaling. Hepatology 2018, 68, 1710–1725. [Google Scholar] [CrossRef] [Green Version]
- Liang, M.; Li, W.; Xiong, S.; Bian, J. MiR-133a-3p targeted regulating CD2AP gene reduces RSV-infected damage of human bronchial epithelial 16-HBE cell line. Basic Clin. Med. 2021, 41, 1463–1469. (In Chinese) [Google Scholar] [CrossRef]
- Wang, H.; Wang, S.; Chiufai, K.; Wang, Q.; Cheng, X. Umbelliferone ameliorates renal function in diabetic nephropathy rats through regulating inflammation and TLR/NF-κB pathway. Chin. J. Nat. Med. 2019, 17, 346–354. [Google Scholar] [CrossRef]
- Wang, D.; Chen, J.; Ding, Y.; Kong, H.; You, H.; Zhao, Y.; Wei, H.; Liu, Y. MiR-188-5p promotes tumor growth by targeting CD2AP through PI3K/AKT/mTOR signaling in children with acute promyelocytic leukemia. Onco. Targets Ther. 2020, 13, 6681–6697. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, L.; Wu, C.; Sun, S.; Pan, J. E2F2 directly regulates the STAT1 and PI3K/AKT/NF-κB pathways to exacerbate the inflammatory phenotype in rheumatoid arthritis synovial fibroblasts and mouse embryonic fibroblasts. Arthritis Res. Ther. 2018, 20, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahmani, F.; Asgharzadeh, F.; Avan, A.; Barneh, F.; Parizadeh, M.R.; Ferns, G.A.; Ryzhikov, M.; Ahmadian, M.R.; Giovannetti, E.; Jafari, M.; et al. Rigosertib potently protects against colitis-associated intestinal fibrosis and inflammation by regulating PI3K/AKT and NF-kappaB signaling pathways. Life Sci. 2020, 249, 117470. [Google Scholar] [CrossRef]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-κB pathway for the therapy of diseases: Mechanism and clinical study. Signal. Transduct. Target. Ther. 2020, 5, 209. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Ren, Q.; Wang, X.; Luoreng, Z.; Wei, D. Bta-miR-199a-3p Inhibits LPS-Induced Inflammation in Bovine Mammary Epithelial Cells via the PI3K/AKT/NF-κB Signaling Pathway. Cells 2022, 11, 3518. https://doi.org/10.3390/cells11213518
Li Y, Ren Q, Wang X, Luoreng Z, Wei D. Bta-miR-199a-3p Inhibits LPS-Induced Inflammation in Bovine Mammary Epithelial Cells via the PI3K/AKT/NF-κB Signaling Pathway. Cells. 2022; 11(21):3518. https://doi.org/10.3390/cells11213518
Chicago/Turabian StyleLi, Yuhang, Qianqian Ren, Xingping Wang, Zhuoma Luoreng, and Dawei Wei. 2022. "Bta-miR-199a-3p Inhibits LPS-Induced Inflammation in Bovine Mammary Epithelial Cells via the PI3K/AKT/NF-κB Signaling Pathway" Cells 11, no. 21: 3518. https://doi.org/10.3390/cells11213518