
The Immediate Early Response of Lens Epithelial Cells to Lens Injury
 , ,
, ,
Abstract
:1. Introduction
2. Methods
2.1. Animals
2.2. Morphological Analysis
2.3. Mouse Cataract Surgery Model
2.4. Next Generation RNA Sequencing and Bioinformatic Analysis
2.5. Pathway Analyses
2.6. Lens Dissections, Culture and RNA Extractions
2.7. cDNA Synthesis and Quantitative RT-PCR
2.8. Immunofluorescence and Confocal Imaging
2.9. ImageJ Quantification and Statistical Analysis
3. Results
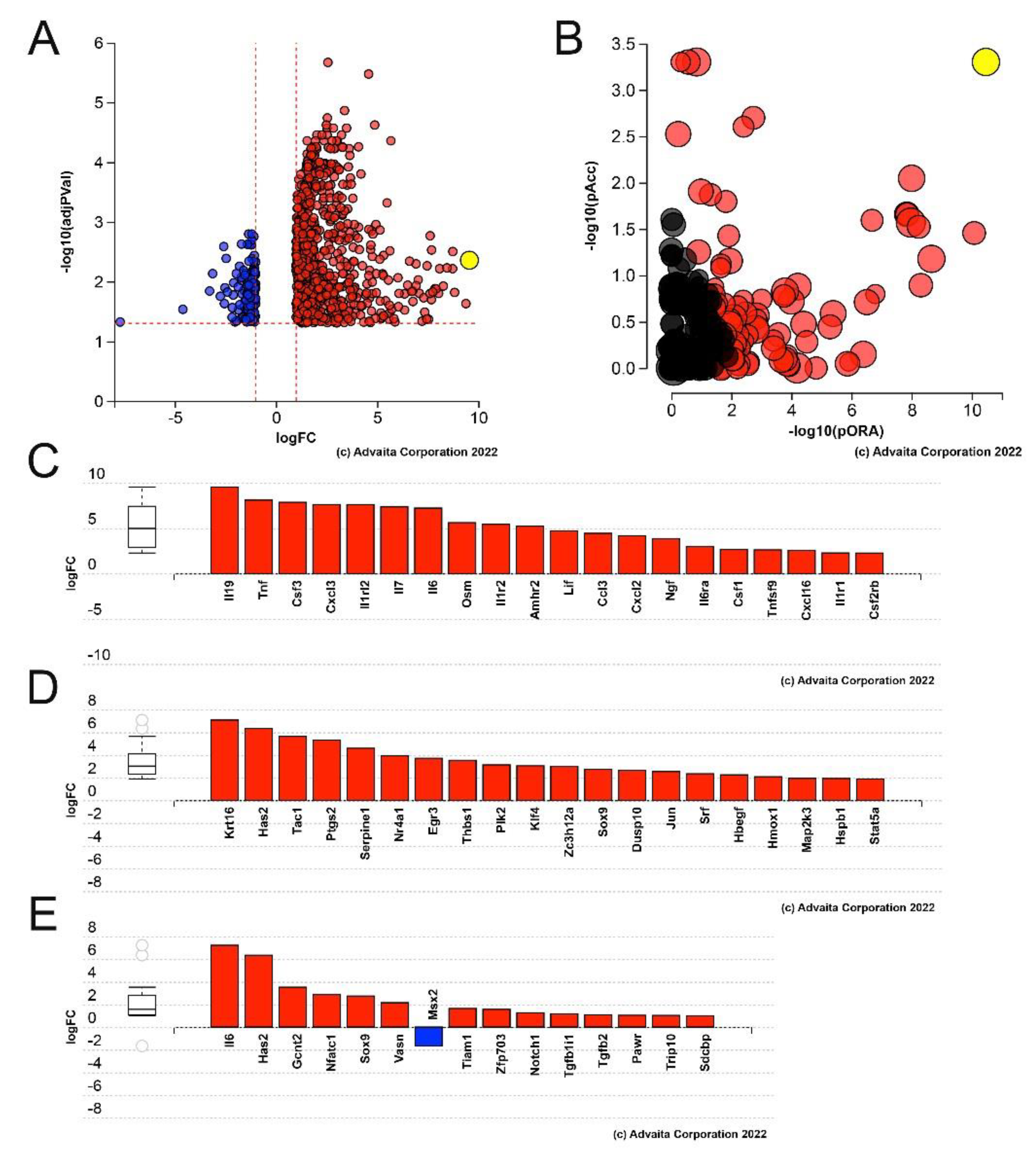
3.1. Lens Epithelial Cells Upregulate the Expression of Numerous Pro-Inflammatory Cytokines by 6 h PCS, but Do Not Yet Upregulate the Expression of Fibrotic Markers
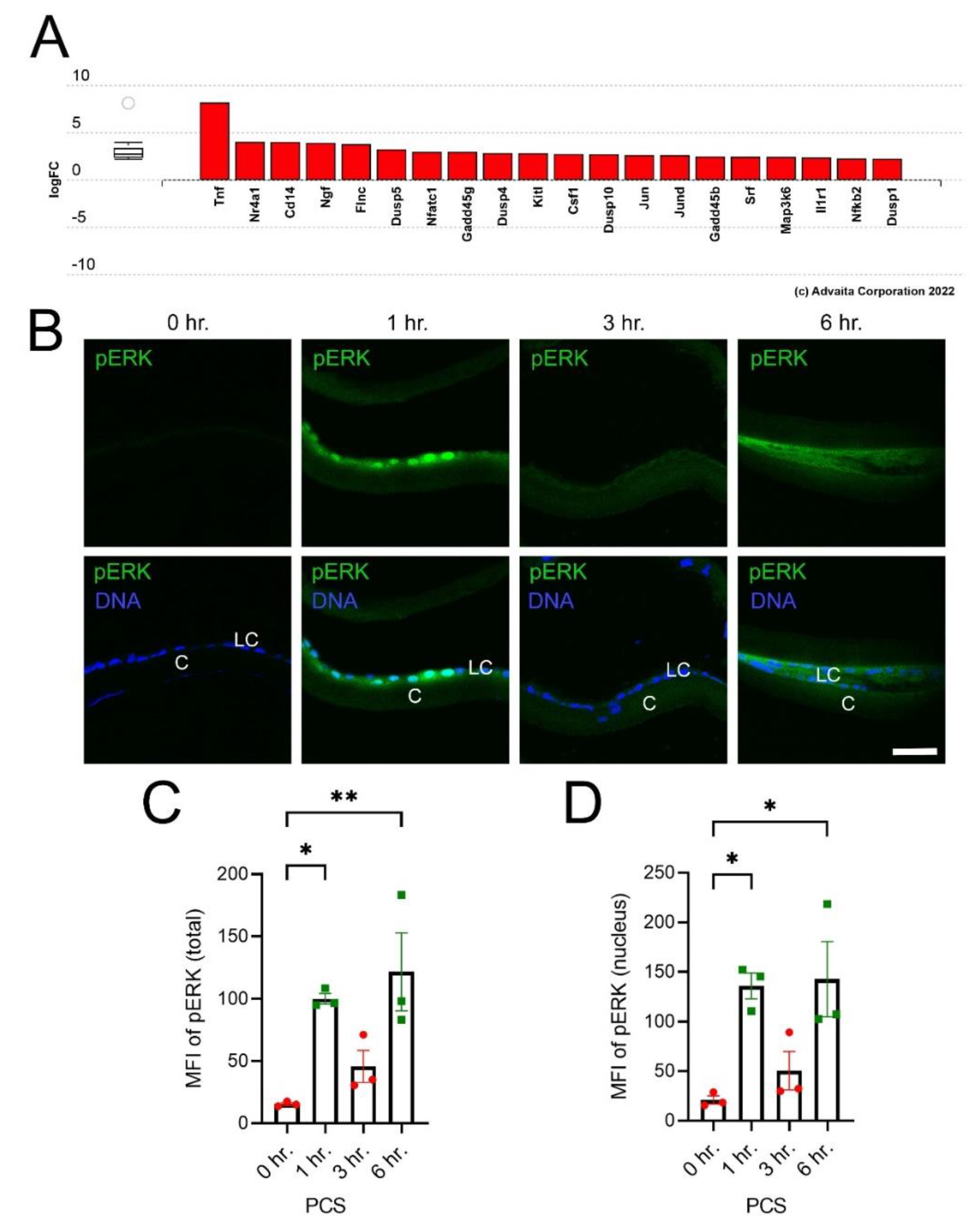
3.2. Many Immediate Early Transcription Factors Are Upregulated in LECs by 6 h PCS
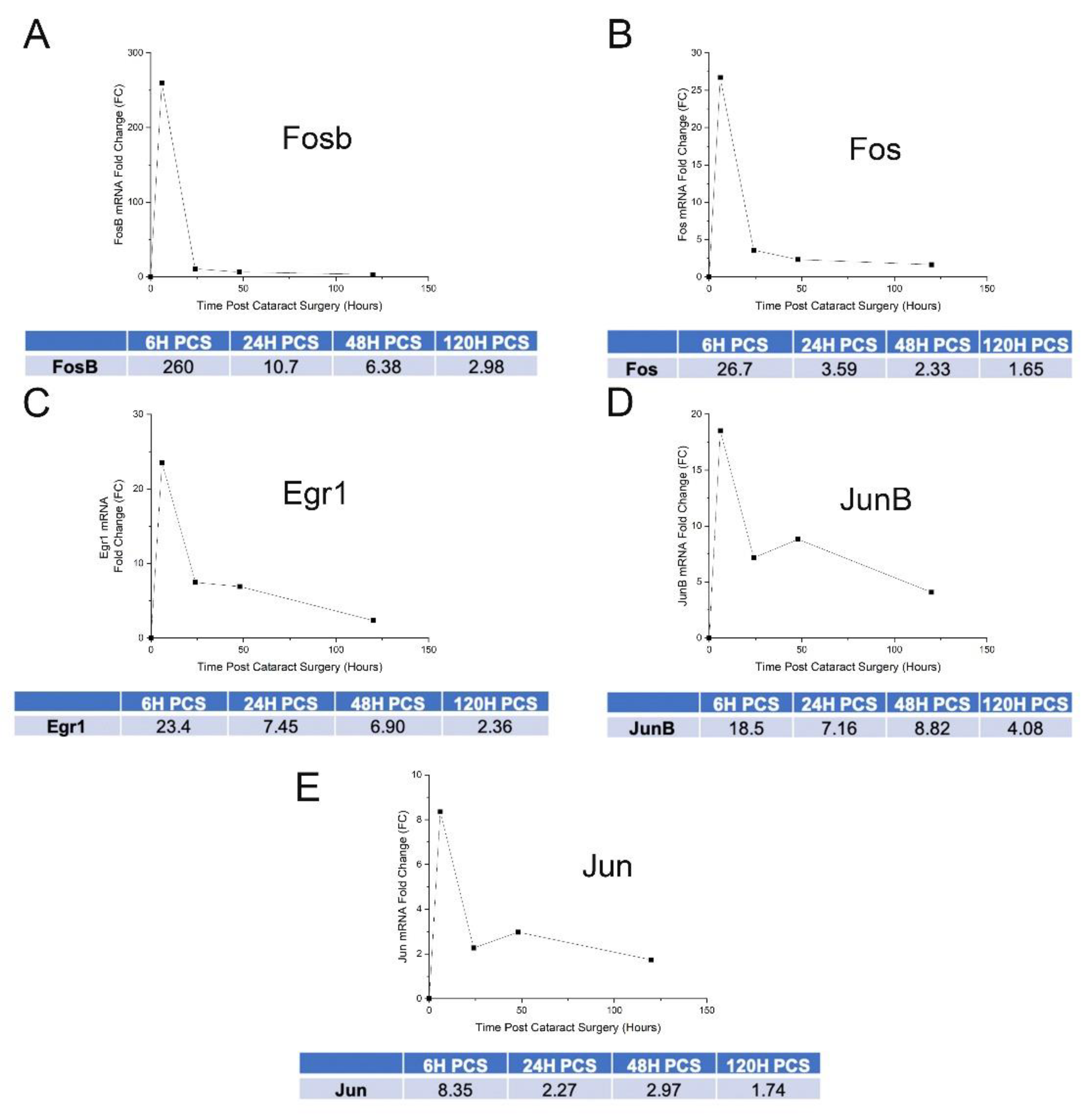
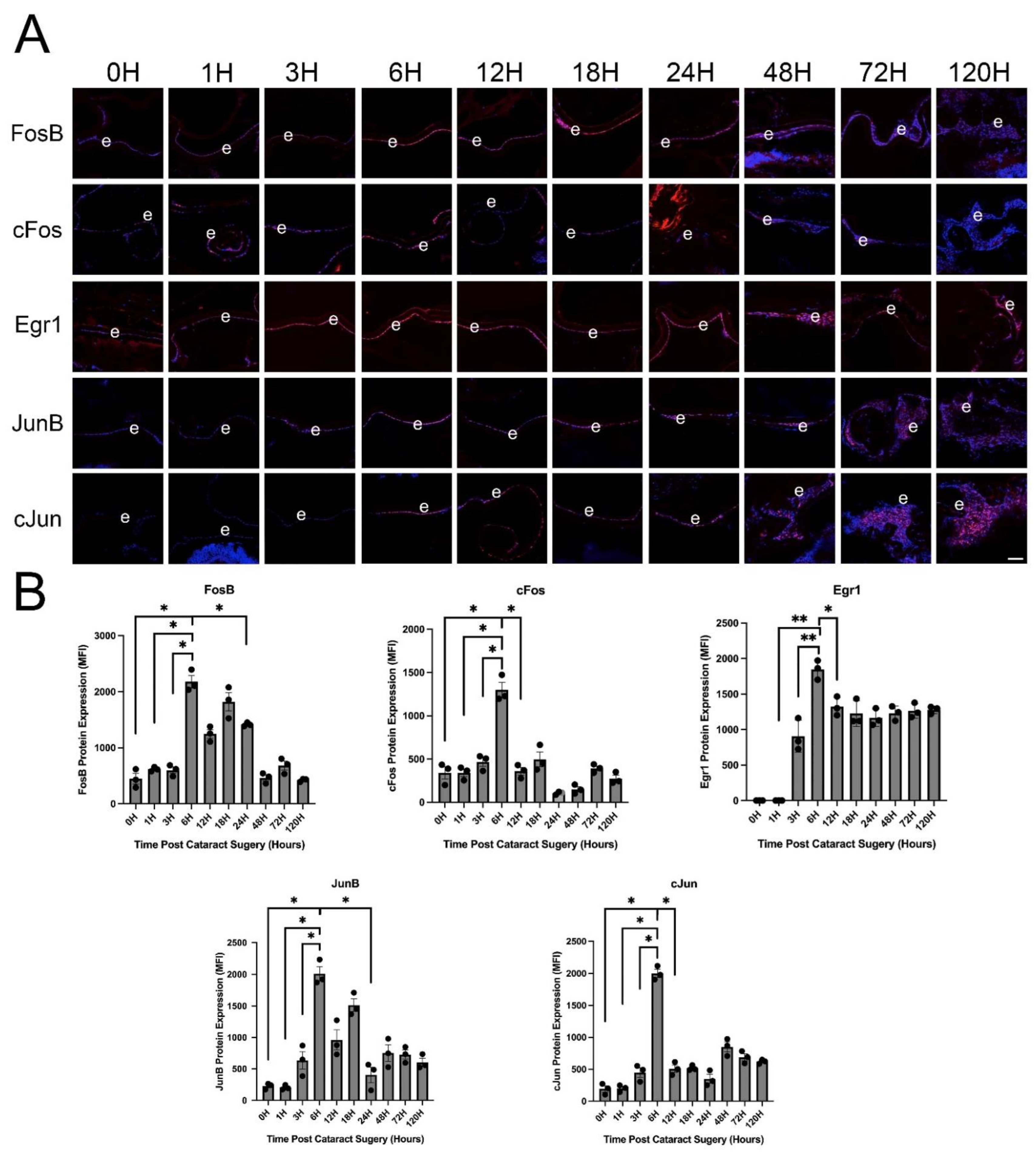
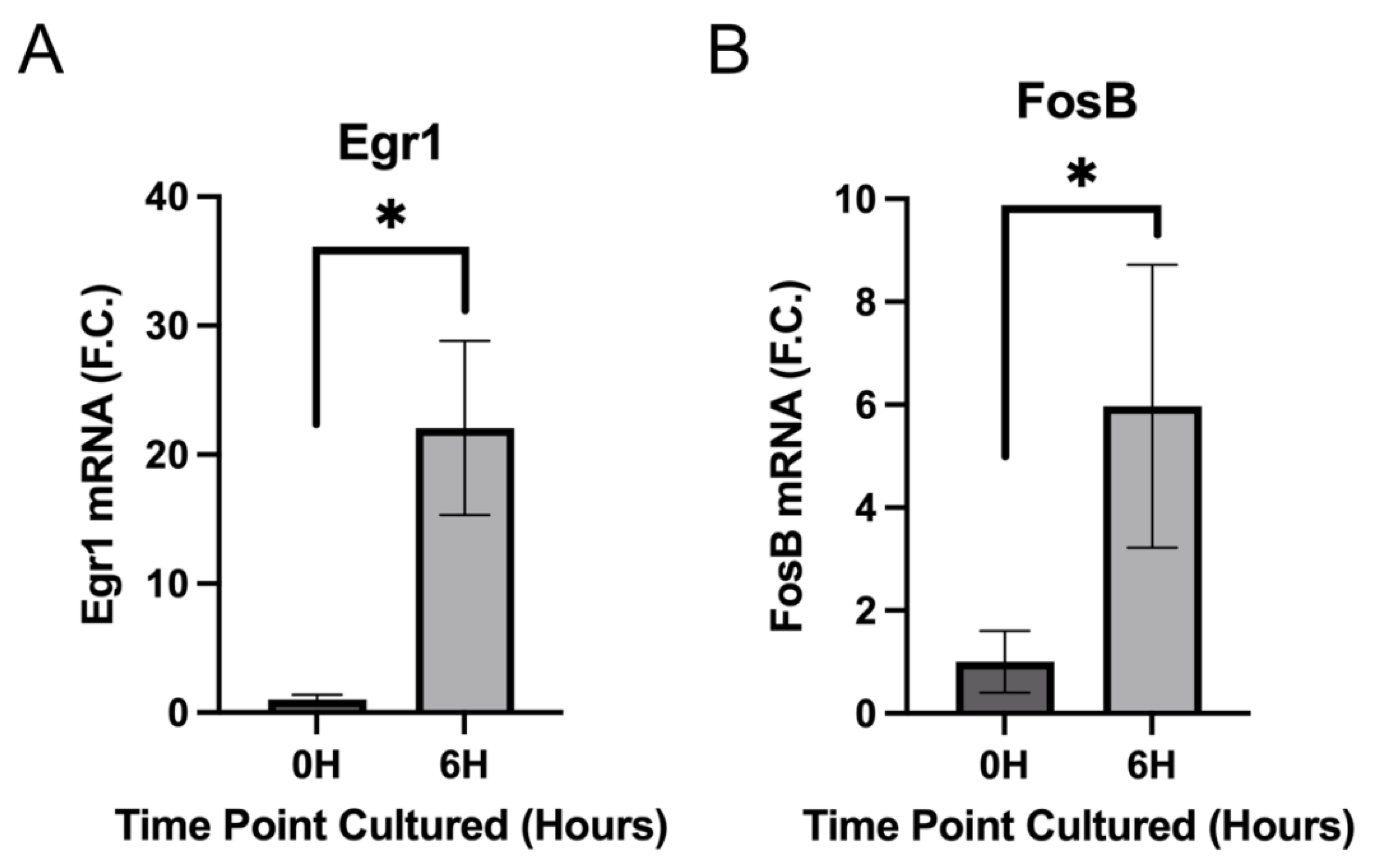
3.3. IETF MRNA and Protein Levels Acutely Upregulate in LECs at 6 h PCS then Rapidly Fall
3.4. Removal of the Gene Encoding the IETF Egr1 from the Lens Only Has a Minor Impact on Lens Phenotype
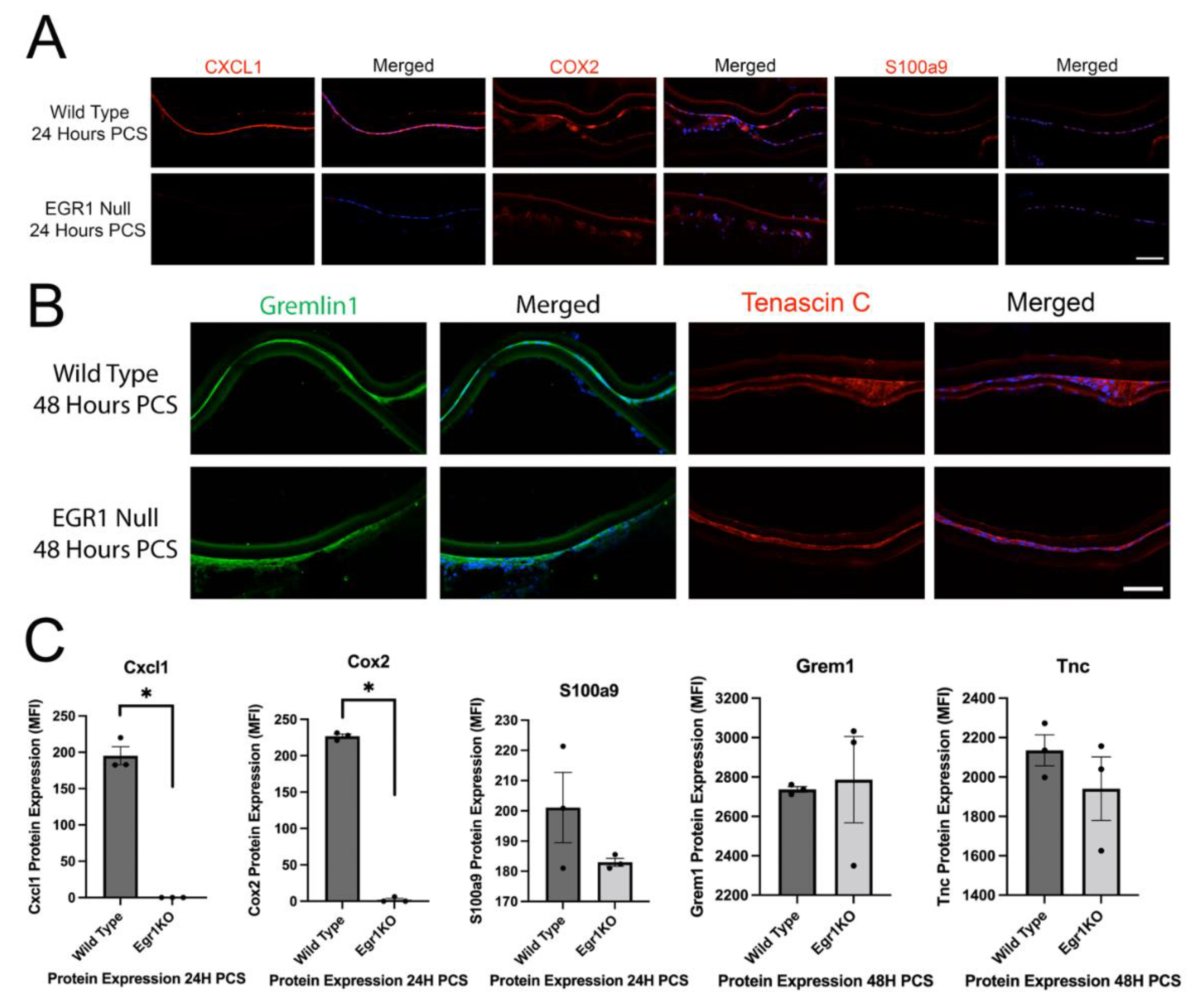
3.5. Egr1 May Mediate a Portion of the Injury Response of LECs PCS
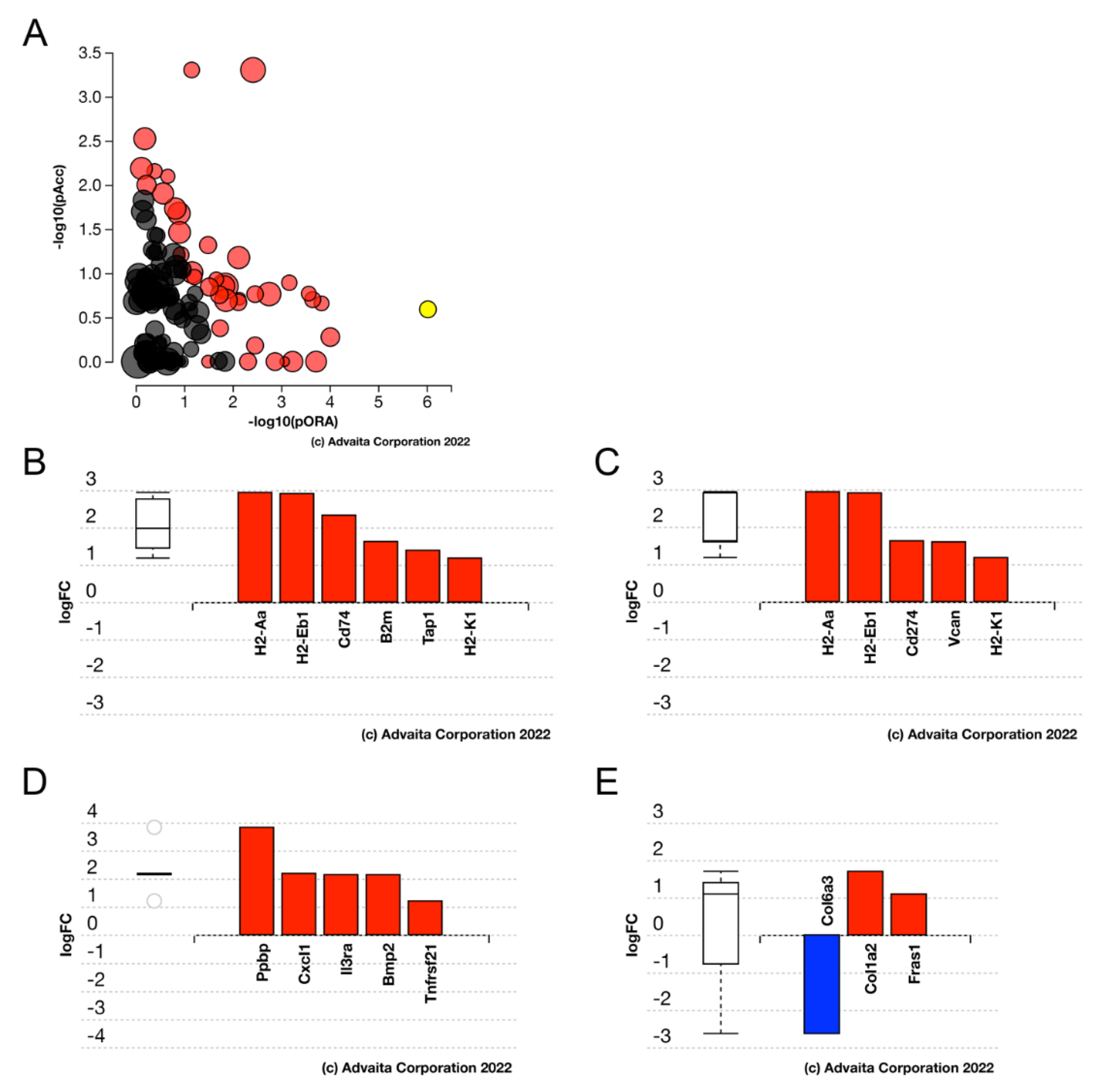
3.6. Deletion of the IETF Egr1 Only Slightly Affects the Transcriptome of Uninjured LECs but Altered a Portion of the Acute LEC Injury Response
3.7. Egr1KO LECs Upregulate Zonule Associated Genes upon Lens Injury
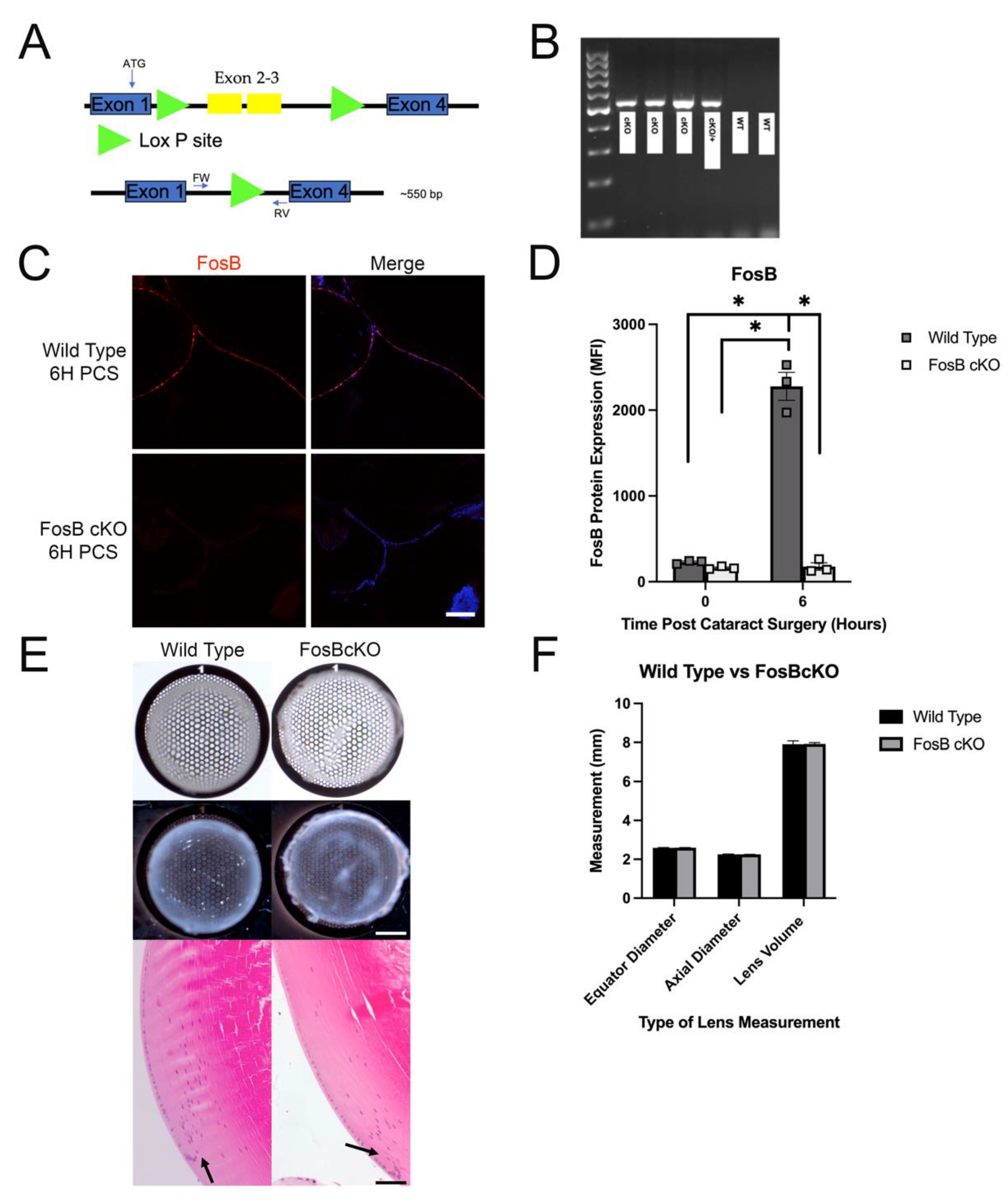
3.8. Generation, Validation and Morphological Analysis of FosB Conditional Knock Out Mice
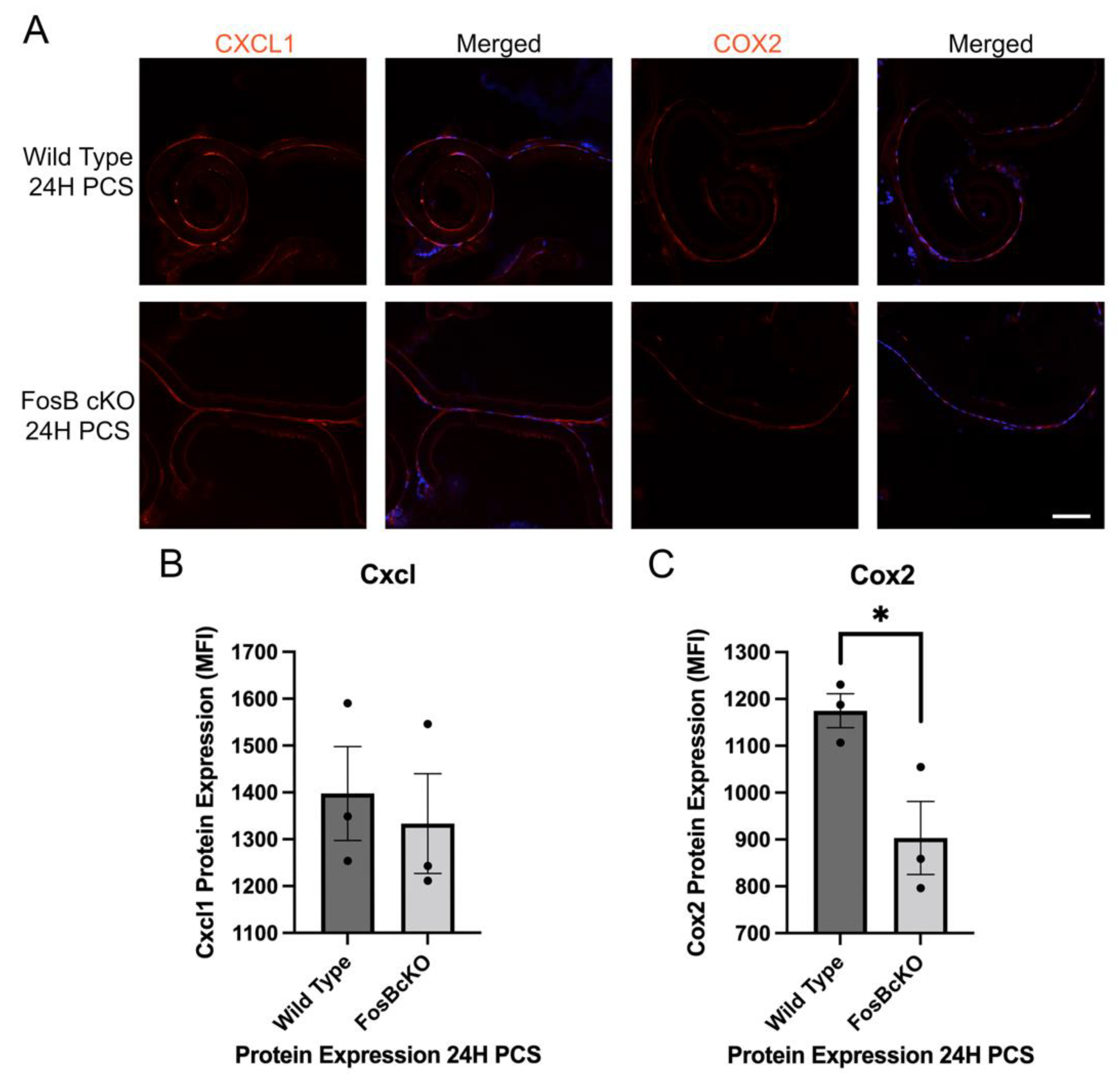
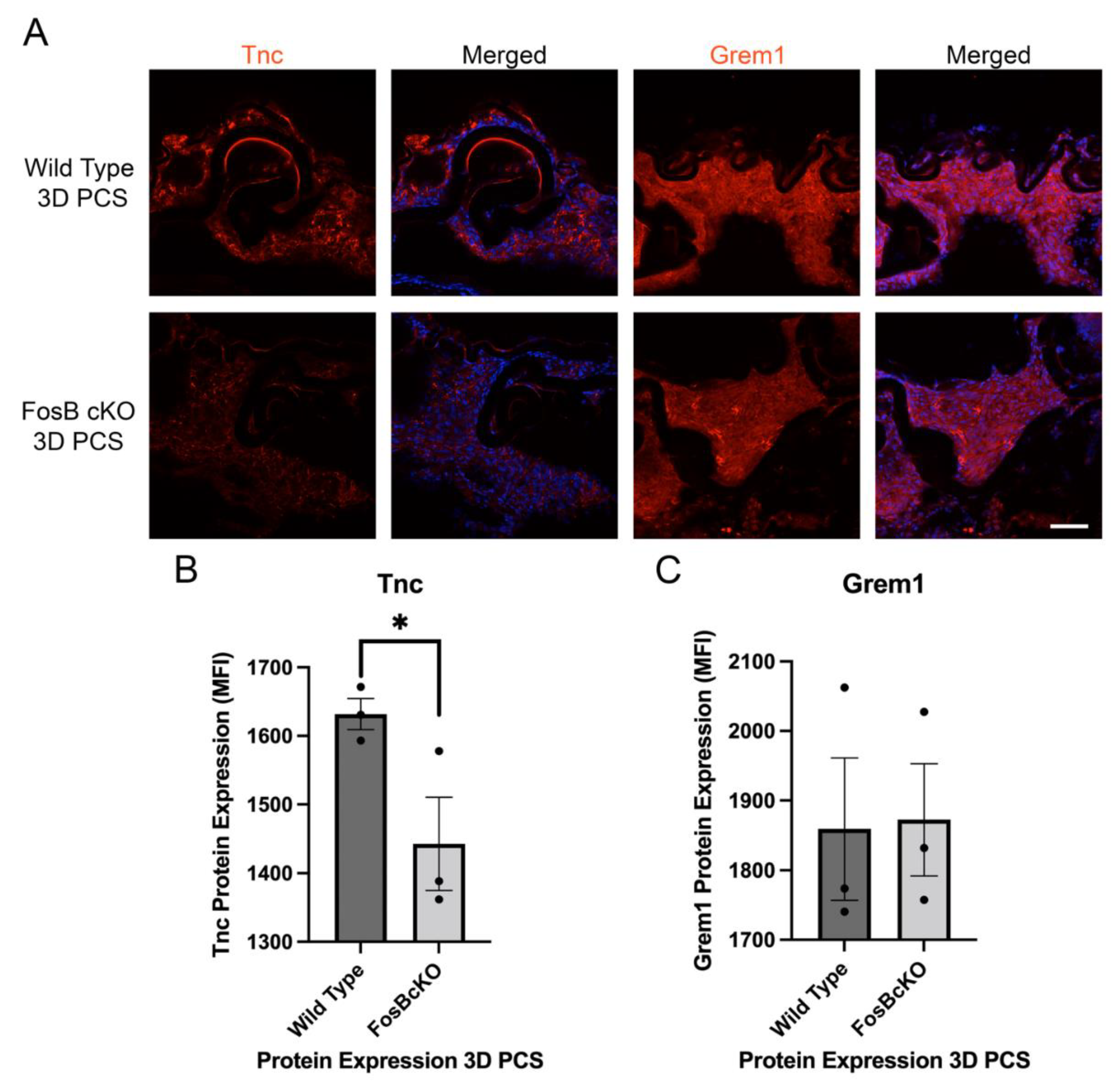
3.9. Acute FosB Elevation PCS May Regulate a Portion of Both the Inflammatory and Fibrotic Responses Post Cataract Surgery
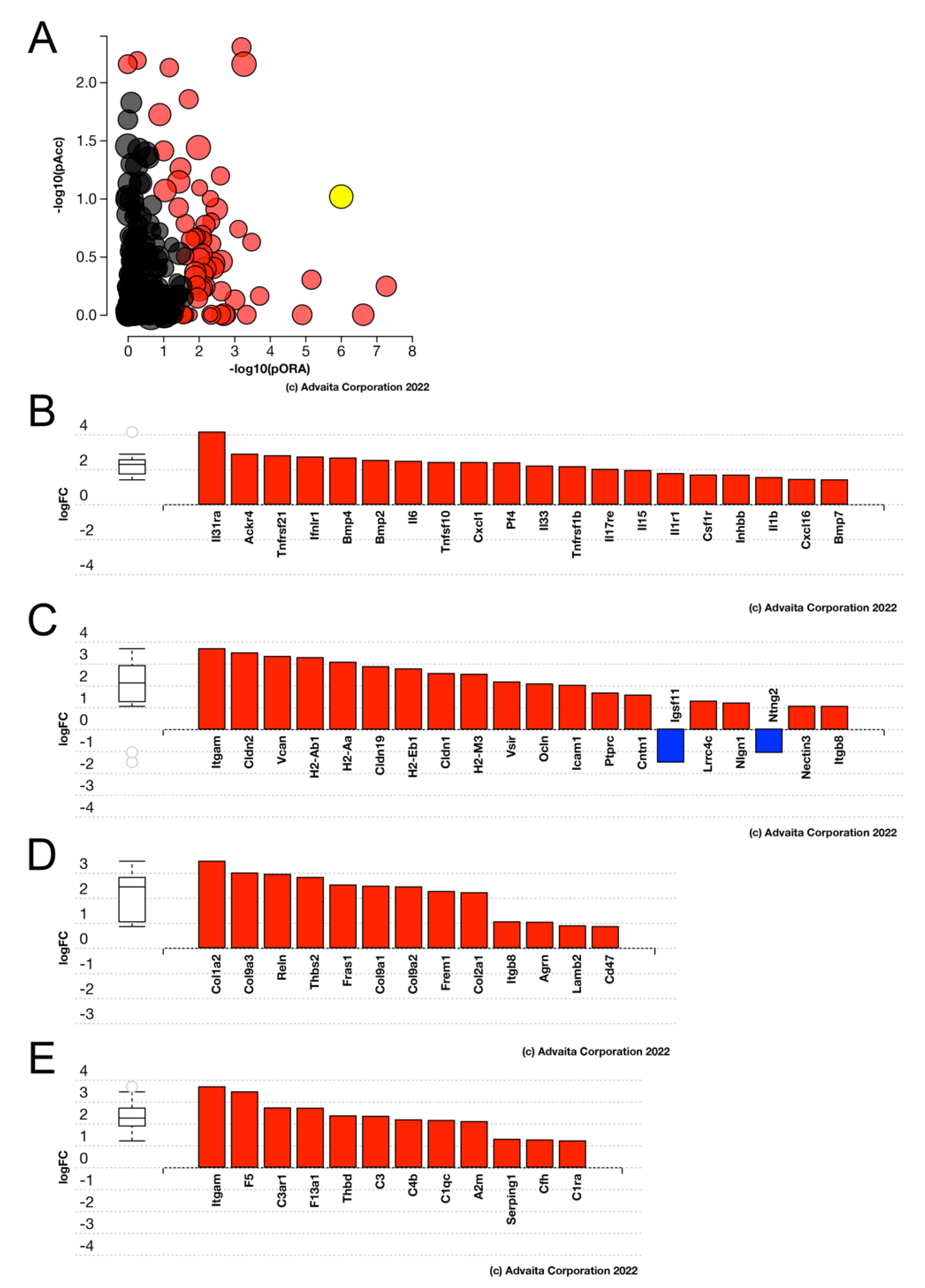
3.10. Acute FosB Upregulation PCS Regulates Many Genes in Injured LECs
4. Discussion
4.1. LECs Rapidly Induce the Expression of Pro-Inflammatory Cytokines after Lens Fiber Cell Removal but Fibrotic Responses Are More Delayed
4.2. Mitogen Activated Protein Kinase Signaling and Immediate Early Transcription Factor Expression Are Rapidly Induced in LECs |Following| Lens Fiber Cell Removal
4.3. The IETFs Egr1 and FosB Regulate Some Aspects of the Lens Wound Healing Response
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wormstone, M.; Wride, M.A. The ocular lens: A classic model for development, physiology and disease. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 1190–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.C.; Wilkins, M.; Kim, T.; Malyugin, B.; Mehta, J.S. Cataracts. Lancet 2017, 390, 600–612. [Google Scholar] [CrossRef]
- Ang, B. Cataract Surgery Steps: From Start to Finish. Victoria, Australia: Vision-and-Eye-Health.com. 2022. Available online: https://www.vision-and-eye-health.com/cataractsurgery-steps.html (accessed on 24 October 2022).
- Khairallah, M.; Kahloun, R.; Bourne, R.; Limburg, H.; Flaxman, S.R.; Jonas, J.B.; Keeffe, J.; Leasher, J.; Naidoo, K.; Pesudovs, K.; et al. Number of People Blind or Visually Impaired by Cataract Worldwide and in World Regions, 1990 to 2010. Investig. Opthalmol. Vis. Sci. 2015, 56, 6762–6769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diagourtas, A.; Petrou, P.; Georgalas, I.; Oikonomakis, K.; Giannakouras, P.; Vergados, A.; Papaconstantinou, D. Bleb failure and intraocular pressure rise following Nd: Yag laser capsulotomy. BMC Ophthalmol. 2017, 17, 18. [Google Scholar] [CrossRef] [Green Version]
- Ascaso, F.J.; Huerva, V.; Grzybowski, A. The Role of Inflammation in the Pathogenesis of Macular Edema Secondary to Retinal Vascular Diseases. Mediat. Inflamm. 2014, 2014, 432685. [Google Scholar] [CrossRef] [PubMed]
- Teh, B.L.; Megaw, R.; Borooah, S.; Dhillon, B. Optimizing cataract surgery in patients with age-related macular degeneration. Surv. Ophthalmol. 2017, 62, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Palestine, A.G.; Pecen, P.E.; Siringo, F.; Colo, A. Retinal Detachments in Patients with Uveitis. Newtown Square, PA, USA: Review of Ophthalmology. 2017. Available online: https://www.reviewofophthalmology.com/article/retinal-detachments-in-patients-with-uveitis (accessed on 10 July 2019).
- Bhutto, I.; Lutty, G. Understanding age-related macular degeneration (AMD): Relationships between the photoreceptor/retinal pigment epithelium/Bruch’s membrane/choriocapillaris complex. Mol. Asp. Med. 2012, 33, 295–317. [Google Scholar] [CrossRef] [Green Version]
- Abbouda, A.; Tortorella, P.; Restivo, L.; Santoro, E.; De Marco, F.; La Cava, M. Follow-Up Study of Over Three Years of Patients with Uveitis after Cataract Phacoemulsification: Outcomes and Complications. Semin. Ophthalmol. 2015, 31, 532–541. [Google Scholar] [CrossRef]
- Shihan, M.H.; Novo, S.G.; Wang, Y.; Sheppard, D.; Atakilit, A.; Arnold, T.D.; Rossi, N.M.; Faranda, A.P.; Duncan, M.K. αVβ8 integrin targeting to prevent posterior capsular opacification. JCI Insight 2021, 6, e145715. [Google Scholar] [CrossRef]
- Wormstone, I.M. Posterior Capsule Opacification: A Cell Biological Perspective. Exp. Eye Res. 2002, 74, 337–347. [Google Scholar] [CrossRef]
- Juthani, V.V.; Clearfield, E.; Chuck, R.S. Non-steroidal anti-inflammatory drugs versus corticosteroids for controlling inflammation after uncomplicated cataract surgery. Cochrane Database Syst. Rev. 2017, 2017, CD010516. [Google Scholar] [CrossRef]
- Colin, J. The Role of NSAIDs in the Management of Postoperative Ophthalmic Inflammation. Drugs 2007, 67, 1291–1308. [Google Scholar] [CrossRef] [PubMed]
- Liegner, J.; Grzybowski, A.; Galloway, M.; Lindstrom, R. Dropless Cataract Surgery: An Overview. Curr. Pharm. Des. 2017, 23, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Sawa, M. Laser flare-cell photometer: Principle and significance in clinical and basic ophthalmology. Jpn. J. Ophthalmol. 2016, 61, 21–42. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Li, P.; Geng, W.; Qin, M.; Su, S.; Zhou, T.; Yuan, Y.; Zhang, G.; Wu, J.; Ji, M.; et al. Cytokines possibly involved in idiopathic epiretinal membrane progression after uncomplicated cataract surgery. Exp. Eye Res. 2022, 217, 108957. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Shihan, M.H.; Wang, Y.; Duncan, M.K. Lens Epithelial Cells Initiate an Inflammatory Response Following Cataract Surgery. Investig. Opthalmology Vis. Sci. 2018, 59, 4986–4997. [Google Scholar] [CrossRef] [Green Version]
- Eldred, J.A.; Zheng, J.; Chen, S.; Wormstone, I.M. An In Vitro Human Lens Capsular Bag Model Adopting a Graded Culture Regime to Assess Putative Impact of IOLs on PCO Formation. Investig. Opthalmology Vis. Sci. 2019, 60, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Raj, S.M.; Vasavada, A.R.; Johar, S.R.K.; Vasavada, V.A.; Vasavada, V.A. Post-operative capsular opacification: A review. Int. J. Biomed. Sci. 2007, 3, 237–250. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3614664/ (accessed on 12 October 2021).
- Findl, O.; Neumayer, T.; Hirnschall, N.; Buehl, W. Natural Course of Elschnig Pearl Formation and Disappearance. Investig. Opthalmology Vis. Sci. 2010, 51, 1547–1553. [Google Scholar] [CrossRef] [Green Version]
- Bisevac, J.; Anisimova, N.S.; Nagymihály, R.; Kristianslund, O.; Katta, K.; Noer, A.; Sharafetdinov, I.H.; Drolsum, L.; Moe, M.C.; Malyugin, B.E.; et al. Long-term myofibroblast persistence in the capsular bag contributes to the late spontaneous in-the-bag intraocular lens dislocation. Sci. Rep. 2020, 10, 20532. [Google Scholar] [CrossRef]
- Donachie, P.H.J.; Barnes, B.L.; Olaitan, M.; Sparrow, J.M.; Buchan, J.C. The Royal College of Ophthalmologists’ National Ophthalmology Database study of cataract surgery: Report 9, Risk factors for posterior capsule opacification. Eye 2022, 29, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Shihan, M.H.; Novo, S.G.; Duncan, M.K. Cataract surgeon viewpoints on the need for novel preventative anti-inflammatory and anti-posterior capsular opacification therapies. Curr. Med Res. Opin. 2019, 35, 1971–1981. [Google Scholar] [CrossRef] [PubMed]
- Wormstone, I.M.; Eldred, J.A. Experimental models for posterior capsule opacification research. Exp. Eye Res. 2016, 142, 2–12. [Google Scholar] [CrossRef]
- Pande, M.V.; Spalton, D.J.; Kerr-Muir, M.G.; Marshall, J. Postoperative inflammatory response to phacoemulsification and extracapsular cataract surgery: Aqueous flare and cells. J. Cataract Refract. Surg. 1996, 22, 770–774. [Google Scholar] [CrossRef]
- Faranda, A.P.; Shihan, M.H.; Wang, Y.; Duncan, M.K. The effect of sex on the mouse lens transcriptome. Exp. Eye Res. 2021, 209, 108676. [Google Scholar] [CrossRef]
- Faranda, A.P.; Shihan, M.H.; Wang, Y.; Duncan, M.K. The aging mouse lens transcriptome. Exp. Eye Res. 2021, 209, 108663. [Google Scholar] [CrossRef]
- Lee, S.L.; Tourtellotte, L.C.; Wesselschmidt, R.L.; Milbrandt, J. Growth and Differentiation Proceeds Normally in Cells Deficient in the Immediate Early Gene NGFI-A. J. Biol. Chem. 1995, 270, 9971–9977. [Google Scholar] [CrossRef] [Green Version]
- Ohnishi, Y.N.; Eagle, A.L.; Ohnishi, Y.H.; Cahill, M.E.; Wirtz, A.J.; Robinson, A.J.; Nestler, E.J. Generation and Validation of a Floxed Fosb Mouse Line. 2017. Available online: https://www.biorxiv.org/content/10.1101/179309v1 (accessed on 24 October 2022).
- Zhao, H.; Yang, Y.; Rizo, C.M.; Overbeek, P.A.; Robinson, M.L. Insertion of a Pax6 Consensus Binding Site into the αA-Crystallin Promoter Acts as a Lens Epithelial Cell Enhancer in Transgenic Mice. Investig. Opthalmology Vis. Sci. 2004, 45, 1930–1939. [Google Scholar] [CrossRef] [Green Version]
- Scheiblin, D.A.; Gao, J.; Caplan, J.L.; Simirskii, V.N.; Czymmek, K.J.; Mathias, R.T.; Duncan, M.K. Beta-1 integrin is important for the structural maintenance and homeostasis of differentiating fiber cells. Int. J. Biochem. Cell Biol. 2014, 50, 132–145. [Google Scholar] [CrossRef] [Green Version]
- Shiels, A.; King, J.M.; Mackay, D.S.; Bassnett, S. Refractive Defects and Cataracts in Mice Lacking Lens Intrinsic Membrane Protein-2. Investig. Opthalmology Vis. Sci. 2007, 48, 500–508. [Google Scholar] [CrossRef] [Green Version]
- Gokhin, D.S.; Nowak, R.B.; Kim, N.E.; Arnett, E.E.; Chen, A.C.; Sah, R.L.; Clark, J.I.; Fowler, V.M. Tmod1 and CP49 Synergize to Control the Fiber Cell Geometry, Transparency, and Mechanical Stiffness of the Mouse Lens. PLoS ONE 2012, 7, e48734. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mamuya, F.A.; Wang, Y.; Roop, V.H.; Scheiblin, D.A.; Zajac, J.C.; Duncan, M.K. The roles of αVintegrins in lensEMTand posterior capsular opacification. J. Cell. Mol. Med. 2014, 18, 656–670. [Google Scholar] [CrossRef] [PubMed]
- Desai, V.D.; Wang, Y.; Simirskii, V.; Duncan, M.K. CD44 expression is developmentally regulated in the mouse lens and increases in the lens epithelium after injury. Differentiation 2010, 79, 111–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yates, A.D.; Achuthan, P.; Akanni, W.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Azov, A.G.; Bennett, R.; Bhai, J.; et al. Ensembl 2020. Nucleic Acids Res. 2020, 48, D682–D688. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef] [Green Version]
- Phipson, B.; Lee, S.; Majewski, I.J.; Alexander, W.S.; Smyth, G.K. Robust hyperparameter estimation protects against hypervariable genes and improves power to detect differential expression. Ann. Appl. Stat. 2016, 10, 946–963. [Google Scholar] [CrossRef]
- Chen, Y.; Lun, A.T.L.; Smyth, G.K. From reads to genes to pathways: Differential expression analysis of RNA-Seq experiments using Rsubread and the edgeR quasi-likelihood pipeline. F1000Research 2016, 5, 1438. [Google Scholar] [CrossRef] [Green Version]
- Manthey, A.L.; Terrell, A.M.; Lachke, S.A.; Polson, S.W.; Duncan, M.K. Development of novel filtering criteria to analyze RNA-sequencing data obtained from the murine ocular lens during embryogenesis. Genom. Data 2014, 2, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarca, A.L.; Draghici, S.; Khatri, P.; Hassan, S.S.; Mittal, P.; Kim, J.-S.; Kim, C.J.; Kusanovic, J.P.; Romero, R. A novel signaling pathway impact analysis. Bioinformatics 2008, 25, 75–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Draghici, S.; Khatri, P.; Tarca, A.L.; Amin, K.; Done, A.; Voichita, C.; Georgescu, C.; Romero, R. A systems biology approach for pathway level analysis. Genome Res. 2007, 17, 1537–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahsan, S.; Drăghici, S. Identifying Significantly Impacted Pathways and Putative Mechanisms with iPathwayGuide. Curr. Protoc. Bioinform. 2017, 57, 7.15.1–7.15.30. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- A Reed, N.; Oh, D.-J.; Czymmek, K.J.; Duncan, M.K. An immunohistochemical method for the detection of proteins in the vertebrate lens. J. Immunol. Methods 2001, 253, 243–252. [Google Scholar] [CrossRef]
- Shihan, M.H.; Novo, S.G.; Le Marchand, S.J.; Wang, Y.; Duncan, M.K. A simple method for quantitating confocal fluorescent images. Biochem. Biophys. Rep. 2021, 25, 100916. [Google Scholar] [CrossRef]
- Novo, S.G. Injury Drives Inflammation, then Fibrosis: The Response of the Lens Epithelium to Cataract Surgery Dissertation in Partial Fulfillment of the Ph.D. In Biological Sciences; University of Delaware: Newark, DE, USA, 2022; in press; Available online: https://udspace.udel.edu/ (accessed on 24 October 2022).
- Fujimoto, Y.; Kuramoto, N.; Yoneyama, M.; Azuma, Y.-T. Interleukin-19 as an Immunoregulatory Cytokine. Curr. Mol. Pharmacol. 2020, 14, 191–199. [Google Scholar] [CrossRef]
- Kuroda, K.O.; Meaney, M.J.; Uetani, N.; Fortin, Y.; Ponton, A.; Kato, T. ERK-FosB signaling in dorsal MPOA neurons plays a major role in the initiation of parental behavior in mice. Mol. Cell. Neurosci. 2007, 36, 121–131. [Google Scholar] [CrossRef]
- Granet, C.; Vico, A.G.L.; Alexandre, C.; Lafage-Proust, M.-H. MAP and src kinases control the induction of AP-1 members in response to changes in mechanical environment in osteoblastic cells. Cell. Signal. 2002, 14, 679–688. [Google Scholar] [CrossRef]
- Tyssowski, K.; DeStefino, N.R.; Cho, J.-H.; Dunn, C.J.; Poston, R.G.; Carty, C.E.; Jones, R.D.; Chang, S.M.; Romeo, P.; Wurzelmann, M.K.; et al. Different Neuronal Activity Patterns Induce Different Gene Expression Programs. Neuron 2018, 98, 530–546.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tullai, J.W.; Schaffer, M.E.; Mullenbrock, S.; Sholder, G.; Kasif, S.; Cooper, G.M. Immediate-Early and Delayed Primary Response Genes Are Distinct in Function and Genomic Architecture. J. Biol. Chem. 2007, 282, 23981–23995. [Google Scholar] [CrossRef] [Green Version]
- Schippert, R.; Burkhardt, E.; Feldkaemper, M.; Schaeffel, F. Relative Axial Myopia inEgr-1(ZENK) Knockout Mice. Investig. Opthalmology Vis. Sci. 2007, 48, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, T.; Belusko, P.; Walkup, R.D.; Azuma, M.; Shearer, T.R. Involvement of Egr-1 in lens epithelial cell death induced by selenite. Exp. Eye Res. 2006, 82, 874–878. [Google Scholar] [CrossRef] [PubMed]
- Tian, R.; Xu, Y.; Dou, W.-W.; Zhang, H. Bioinformatics analysis of microarray data to explore the key genes involved in HSF4 mutation-induced cataract. Int. J. Ophthalmol. 2018, 11, 910–917. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Terrell, A.M.; Riggio, B.A.; Anand, D.; Lachke, S.; Duncan, M.K. β1-Integrin Deletion From the Lens Activates Cellular Stress Responses Leading to Apoptosis and Fibrosis. Investig. Opthalmology Vis. Sci. 2017, 58, 3896–3922. [Google Scholar] [CrossRef] [Green Version]
- Shihan, M.H.; Kanwar, M.; Wang, Y.; Jackson, E.E.; Faranda, A.P.; Duncan, M.K. Fibronectin has multifunctional roles in posterior capsular opacification (PCO). Matrix Biol. 2020, 90, 79–108. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Wu, M.; Fang, F.; Tourtellotte, W.; Feghali-Bostwick, C.; Varga, J. Early growth response transcription factors: Key mediators of fibrosis and novel targets for anti-fibrotic therapy. Matrix Biol. 2011, 30, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Jones, W.; Rodriguez, J.; Bassnett, S. Targeted deletion of fibrillin-1 in the mouse eye results in ectopia lentis and other ocular phenotypes associated with Marfan syndrome. Dis. Model. Mech. 2019, 12, dmm037283. [Google Scholar] [CrossRef] [Green Version]
- De Maria, A.; Wilmarth, P.A.; David, L.L.; Bassnett, S. Proteomic Analysis of the Bovine and Human Ciliary Zonule. Investig. Opthalmology Vis. Sci. 2017, 58, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.R.; Ye, H.; Bronson, R.T.; Dikkes, P.; Greenberg, M.E. A Defect in Nurturing in Mice Lacking the Immediate Early Gene fosB. Cell 1996, 86, 297–309. [Google Scholar] [CrossRef] [Green Version]
- de Iongh, R.; Wederell, E.; Lovicu, F.; McAvoy, J. Transforming Growth Factor-β-Induced Epithelial-Mesenchymal Transition in the Lens: A Model for Cataract Formation. Cells Tissues Organs 2005, 179, 43–55. [Google Scholar] [CrossRef]
- Leigh, T.; Scalia, R.G.; Autieri, M.V. Resolution of inflammation in immune and nonimmune cells by interleukin-19. Am. J. Physiol. Physiol. 2020, 319, C457–C464. [Google Scholar] [CrossRef]
- Presicce, P.; Park, C.-W.; Senthamaraikannan, P.; Bhattacharyya, S.; Jackson, C.; Kong, F.; Rueda, C.M.; DeFranco, E.; Miller, L.A.; Hildeman, D.A.; et al. IL-1 signaling mediates intrauterine inflammation and chorio-decidua neutrophil recruitment and activation. JCI Insight 2018, 3, e98306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.; Wu, P.W.; Seo, J.J.; Fernando, T.; Jin, M.; Contreras, J.; Montano, E.N.; Gabhann, J.N.; Cunningham, K.; Widaa, A.; et al. IL-16/miR-125a axis controls neutrophil recruitment in pristane-induced lung inflammation. JCI Insight 2018, 3, e120798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, L.S. Role and regulation of cyclooxygenase-2 during inflammation. Am. J. Med. 1999, 106, 37S–42S. [Google Scholar] [CrossRef]
- Lucchini, A.C.; Gachanja, N.N.; Rossi, A.G.; Dorward, D.A.; Lucas, C.D. Epithelial Cells and Inflammation in Pulmonary Wound Repair. Cells 2021, 10, 339. [Google Scholar] [CrossRef]
- Chi, Z.; Melendez, A.J. Role of Cell Adhesion Molecules and Immune-Cell Migration in the Initiation, Onset and Development of Atherosclerosis. Cell Adhes. Migr. 2007, 1, 171–175. [Google Scholar] [CrossRef] [Green Version]
- Camilleri, M. Leaky gut: Mechanisms, measurement and clinical implications in humans. Gut 2019, 68, 1516–1526. [Google Scholar] [CrossRef]
- Naik, S.; Larsen, S.; Gomez, N.C.; Alaverdyan, K.; Sendoel, A.; Yuan, S.; Polak, L.; Kulukian, A.; Chai, S.; Fuchs, E. Inflammatory memory sensitizes skin epithelial stem cells to tissue damage. Nature 2017, 550, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Eldred, J.A.; Dawes, L.J.; Wormstone, I.M. The lens as a model for fibrotic disease. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 1301–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregg, J.; Fraizer, G. Transcriptional Regulation of EGR1 by EGF and the ERK Signaling Pathway in Prostate Cancer Cells. Genes Cancer 2011, 2, 900–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujioka, S.; Niu, J.; Schmidt, C.; Sclabas, G.M.; Peng, B.; Uwagawa, T.; Li, Z.; Evans, D.B.; Abbruzzese, J.L.; Chiao, P.J. NF-κB and AP-1 Connection: Mechanism of NF-κB-Dependent Regulation of AP-1 Activity. Mol. Cell. Biol. 2004, 24, 7806–7819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peroval, M.Y.; Boyd, A.C.; Young, J.R.; Smith, A.L. A Critical Role for MAPK Signalling Pathways in the Transcriptional Regulation of Toll Like Receptors. PLoS ONE 2013, 8, e51243. [Google Scholar] [CrossRef] [Green Version]
- Dulle, J.E.; Fort, P.E. Crystallins and neuroinflammation: The glial side of the story. Biochim. et Biophys. Acta (BBA)-Gen. Subj. 2016, 1860, 278–286. [Google Scholar] [CrossRef] [Green Version]
- Shirai, K.; Saika, S.; Okada, Y.; Miyamoto, T.; Ueyama, T.; Ohnishi, Y. Transcriptional activation in lens epithelial cells following an ocular blunt trauma. J. Cataract Refract. Surg. 2005, 31, 1226–1230. [Google Scholar] [CrossRef]
- Shirai, K.; Okada, Y.; Saika, S.; Senba, E.; Ohnishi, Y. Expression of Transcription Factor AP-1 in Rat Lens Epithelial Cells during Wound Repair. Exp. Eye Res. 2001, 73, 461–468. [Google Scholar] [CrossRef]
- Tang, J.; Salzman, I.J.; Sable, M.D. Traumatic cataract formation after vigorous ocular massage. J. Cataract Refract. Surg. 2003, 29, 1641–1642. [Google Scholar] [CrossRef]
- Gremida, A.; Kassem, I.; Traish, A. Anterior capsular rupture following blunt ocular injury. Digit. J. Ophthalmol. 2011, 17, 66–68. [Google Scholar] [CrossRef]
- Lovicu, F.J.; Schulz, M.W.; Hales, A.M.; Vincent, L.N.; Overbeek, P.A.; Chamberlain, C.G.; McAvoy, J.W. Tgfβ induces morphological and molecular changes similar to human anterior subcapsular cataract. Br. J. Ophthalmol. 2002, 86, 220–226. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1771017/ (accessed on 12 October 2021). [CrossRef] [PubMed]
- Allen, A.; Maddala, R.; Eldawy, C.; Rao, P.V. Mechanical Load and Piezo1 Channel Regulated Myosin II Activity in Mouse Lenses. Int. J. Mol. Sci. 2022, 23, 4710. [Google Scholar] [CrossRef] [PubMed]
- Shishido, S.; Bã¶nig, H.; Kim, Y.-M. Role of Integrin Alpha4 in Drug Resistance of Leukemia. Front. Oncol. 2014, 4, 99. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Hu, J.; Li, G.; Li, Y.; Zhang, J.; Wang, F.; Hu, L.; Fan, Z.; Lü, S.; Ding, G.; et al. Biomechanical stress regulates mammalian tooth replacement via the integrin β1-RUNX2-Wnt pathway. EMBO J. 2019, 39, e102374. [Google Scholar] [CrossRef]
- Ramachandran, A.; Gong, E.M.; Pelton, K.; Ranpura, S.A.; Mulone, M.; Seth, A.; Gomez, P.; Adam, R.M. FosB Regulates Stretch-Induced Expression of Extracellular Matrix Proteins in Smooth Muscle. Am. J. Pathol. 2011, 179, 2977–2989. [Google Scholar] [CrossRef] [PubMed]
- Baron, V.T.; Pio, R.; Jia, Z.; Mercola, D. Early Growth Response 3 regulates genes of inflammation and directly activates IL6 and IL8 expression in prostate cancer. Br. J. Cancer 2015, 112, 755–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahrami, S.; Drabløs, F. Gene regulation in the immediate-early response process. Adv. Biol. Regul. 2016, 62, 37–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreucci, J.J.; Grant, D.; Cox, D.M.; Tomc, L.K.; Prywes, R.; Goldhamer, D.J.; Rodrigues, N.; Bédard, P.-A.; McDermott, J.C. Composition and Function of AP-1 Transcription Complexes during Muscle Cell Differentiation. J. Biol. Chem. 2002, 277, 16426–16432. [Google Scholar] [CrossRef] [Green Version]
- Cervantes-Madrid, D.L.; Nagi, S.; Gustafsson, A.A. FosB transcription factor regulates COX-2 expression in colorectal cancer cells without affecting PGE2 expression. Oncol. Lett. 2017, 13, 1411–1416. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.Y.; Lee, J.M.; Lim, Y.; Lee, Y.H. Transcriptional regulation of the growth-regulated oncogene α gene by early growth response protein-1 in response to tumor necrosis factor α stimulation. Biochim. et Biophys. Acta 2013, 1829, 1066–1074. [Google Scholar] [CrossRef]
- Veremeyko, T.; Yung, A.W.Y.; Anthony, D.C.; Strekalova, T.; Ponomarev, E.D. Early Growth Response Gene-2 Is Essential for M1 and M2 Macrophage Activation and Plasticity by Modulation of the Transcription Factor CEBPβ. Front. Immunol. 2018, 9, 2515. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Xue, C.; Chen, Z.; Jiang, W.; He, S.; Zhang, X. c-Fos is a mechanosensor that regulates inflammatory responses and lung barrier dysfunction during ventilator-induced acute lung injury. BMC Pulm. Med. 2022, 22, 9. [Google Scholar] [CrossRef] [PubMed]
- Zenz, R.; Eferl, R.; Scheinecker, C.; Redlich, K.; Smolen, J.; Schonthaler, H.B.; Kenner, L.; Tschachler, E.; Wagner, E.F. Activator protein 1 (Fos/Jun) functions in inflammatory bone and skin disease. Arthritis Res. Ther. 2007, 10, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelz, M.B.; Kuszak, J.R.; Yang, Y.; Ma, W.; Steffen, C.; Al-Ghoul, K.; Zhang, Y.J.; Chen, J.; Nestler, E.J.; Spector, A. Deltafosb-induced cataract. Invest. Ophthalmol. Vis. Sci. 2000, 41, 3523–3538. Available online: https://www.ncbi.nlm.nih.gov/pubmed/11006248 (accessed on 16 September 2022). [PubMed]
- McClung, C.A.; Nestler, E.J. Regulation of gene expression and cocaine reward by CREB and ΔFosB. Nat. Neurosci. 2003, 6, 1208–1215. [Google Scholar] [CrossRef]
- Nestler, E.J. Transcriptional mechanisms of addiction: Role of ΔFosB. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 3245–3255. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Name | Wild Type 0H FPKM | Wild Type 6H FPKM | Fold Change | FDR |
|---|---|---|---|---|---|
| Il19 | Interleukin 19 | 0.00 | 3.38 | 753.97 | 4.39 × 10−3 |
| Krtap4-16 | Keratin associated protein 4–16 | 0.00 | 2.14 | 427.68 | 3.14 × 10−3 |
| Csf3 | Colony Stimulating Factor 3 | 0.11 | 21.34 | 232.91 | 1.31 × 10−2 |
| Il6 | Interleukin 6 | 0.07 | 8.55 | 149.03 | 4.61 × 10−2 |
| Fosb | FBJ osteosarcoma oncogene B | 1.89 | 198.30 | 130.49 | 9.36 × 10−3 |
| Ptx3 | Pentraxin 3 | 1.08 | 105.47 | 120.93 | 5.05 × 10−3 |
| Esm1 | Endothelial cell-specific molecule 1 | 0.16 | 12.52 | 96.90 | 4.56 × 10−3 |
| Iqcn | IQ motif containing N | 0.16 | 8.66 | 67.98 | 1.58 × 10−2 |
| C2cd4b | C2 calcium-dependent domain containing 4B | 0.15 | 8.17 | 65.46 | 8.63 × 10−3 |
| Emp1 | Epithelial membrane protein 1 | 1.85 | 76.36 | 51.28 | 4.39 × 10−5 |
| Ifi202b | Interferon activated gene 202B | 0.09 | 3.64 | 47.32 | 9.48 × 10−3 |
| Arc | Activity regulated cytoskeletal associated protein | 5.64 | 183.82 | 39.94 | 3.64 × 10−3 |
| Ptgs2 | Prostaglandin-endoperoxide synthase 2 | 0.65 | 20.96 | 39.62 | 7.93 × 10−3 |
| Plaur | Plasminogen activator, urokinase receptor | 0.18 | 5.54 | 36.69 | 4.17 × 10−2 |
| Klf2 | Kruppel-like factor 2 | 1.14 | 28.17 | 30.60 | 1.11 × 10−3 |
| Htr1d | 5-hydroxytrptamine receptor 1D | 0.54 | 13.06 | 29.37 | 2.39 × 10−5 |
| Rnf125 | Ring finger protein 125 | 0.18 | 4.17 | 27.65 | 1.60 × 10−3 |
| F3 | Coagulation factor III | 1.97 | 43.91 | 27.01 | 3.80 × 10−3 |
| Lif | Leukemia inhibitory factor | 1.50 | 33.10 | 26.40 | 1.41 × 10−3 |
| Fgl2 | Fibrinogen-like protein 2 | 0.56 | 11.78 | 26.39 | 3.12 × 10−2 |
| Gene Symbol | Gene Name | Wild Type 0 h FPKM | Wild Type 6 h FPKM | Fold Change | FDR |
|---|---|---|---|---|---|
| Col1a1 | Collagen, type I, alpha 1 | 1.53 | 7.24 | 5.80 | 2.14 × 10−1 |
| Tnc | Tenascin C | 0.65 | 1.10 | 2.10 | 3.43 × 10−1 |
| Acta2 | α Smooth Muscle Actin | 53.17 | 63.90 | 1.48 | 7.06 × 10−1 |
| Itgav | Integrin alpha V | 27.55 | 31.57 | 1.42 | 7.71 × 10−2 |
| Vtn | Vitronectin | 23.57 | 27.60 | 1.41 | 6.02 × 10−1 |
| Fn1 | Fibronectin 1 | 2.79 | 2.16 | −1.03 | 9.70 × 10−1 |
| Itgb1 | Integrin beta 1 | 56.79 | 56.59 | 1.24 | 4.17 × 10−1 |
| Gene Symbol | Gene Name | Wild Type 0 h FPKM | Wild Type 6 h FPKM | Fold Change | FDR |
|---|---|---|---|---|---|
| Fosb | FBJ osteosarcoma oncogene B | 1.89 | 198.30 | 130.49 | 9.36 × 10−3 |
| Klf2 | Kruppel-like factor 2 | 1.14 | 28.17 | 30.60 | 1.11 × 10−3 |
| Nr4a1 | Nuclear receptor subfamily 4, group A, member 1 | 8.67 | 105.28 | 15.14 | 3.35 × 10−2 |
| Zfp36 | Zinc finger protein 36 | 13.93 | 163.10 | 14.56 | 7.59 × 10−3 |
| Ier2 | Immediate early response 2 | 10.16 | 111.54 | 13.66 | 1.43 × 10−2 |
| Egr3 | Early growth response 3 | 1.26 | 13.40 | 13.09 | 2.40 × 10−2 |
| Ier5 | Immediate early response 5 | 5.55 | 55.82 | 12.30 | 5.95 × 10−5 |
| Egr1 | Early growth response 1 | 33.41 | 328.89 | 12.10 | 3.09 × 10−2 |
| Atf3 | Activating transcription factor 3 | 33.02 | 268.96 | 10.17 | 3.76 × 10−2 |
| Ier3 | Immediate early response 3 | 3.45 | 28.17 | 9.94 | 2.35 × 10−4 |
| Junb | Jun B proto-oncogene | 20.98 | 160.07 | 9.51 | 2.97 × 10−3 |
| Maff | V-maf musculoaponeurotic fibrosarcoma oncogene family, protein F | 12.55 | 89.64 | 8.82 | 1.31 × 10−4 |
| Klf4 | Kruppel-like factor 4 | 25.06 | 166.32 | 8.21 | 3.38 × 10−3 |
| Egr2 | Early growth response 2 | 0.88 | 5.49 | 7.61 | 4.84 × 10−2 |
| Gene Symbol | Gene Name | Wild Type 6 h FPKM | Egr1KO 6 h FPKM | Fold Change | FDR |
|---|---|---|---|---|---|
| Pebp4 | Phosphatidylethanolamine binding protein 4 | 0.14 | 2.18 | 14.58 | 3.33 × 10−3 |
| Dio3 | Deiodinase, iodothyronine type III | 0.38 | 3.29 | 8.56 | 9.90 × 10−4 |
| H2-Aa | Histocompatibility 2, class II antigen A, alpha | 0.37 | 2.88 | 7.67 | 4.98 × 10−3 |
| H2-Eb1 | Histocompatibility 2, class II antigen E beta | 0.87 | 6.63 | 7.50 | 1.23 × 10−2 |
| Cd74 | CD74 antigen | 4.77 | 24.04 | 5.02 | 2.73 × 10−2 |
| Cxcl1 | Chemokine (C-X-C motif) ligand 1 | 15.11 | 68.71 | 4.55 | 7.51 × 10−7 |
| Fbn2 | Fibrillin 2 | 0.60 | 2.65 | 4.42 | 2.60 × 10−3 |
| Il3ra | Interleukin 3 receptor, alpha chain | 0.71 | 3.17 | 4.41 | 2.79 × 10−2 |
| Bmp2 | Bone morphogenetic protein 2 | 5.37 | 23.64 | 4.40 | 5.02 × 10−4 |
| Slc6a13 | Solute carrier family 6, member 13 | 2.60 | 11.31 | 4.34 | 3.62 × 10−3 |
| Gene Code | Gene Name | Wild Type 6 h FPKM | Egr1KO 6 h FPKM | Fold Change | FDR |
|---|---|---|---|---|---|
| Sst | Somatostatin | 6.71 | 0.13 | −39.55 | 4.98 × 10−3 |
| Krt5 | Keratin 5 | 14.06 | 0.64 | −21.81 | 2.89 × 10−2 |
| C1ql2 | Complement component 1 | 4.94 | 0.68 | −7.08 | 9.20 × 10−3 |
| Rit2 | Ras-like without CAAX 2 | 3.51 | 0.52 | −6.66 | 4.24 × 10−2 |
| Zfp804a | Zinc finger protein 804A | 2.42 | 0.37 | −6.32 | 2.22 × 10−2 |
| Col6a3 | Collagen, type VI, alpha 3 | 36.99 | 5.97 | −6.19 | 3.91 × 10−3 |
| Spc25 | SPC25 | 14.50 | 2.88 | −5.02 | 2.14 × 10−3 |
| Map2k3os | Mitogen-activated protein kinase kinase 3, opposite strand | 4.04 | 0.77 | −5.01 | 2.89 × 10−2 |
| Rab7b | RAB7B, member Ras oncogene family | 4.88 | 1.01 | −4.79 | 2.22 × 10−2 |
| Galnt13 | Polypeptide N-acetylgalactosaminyltransferase13 | 2.76 | 0.64 | −4.25 | 2.22 × 10−2 |
| Gene Code. | Gene Name | Wild Type 6 h FPKM | Egr1KO 6 h FPKM | Fold Change | FDR |
|---|---|---|---|---|---|
| Fbn2 | Fibrillin 2 | 0.60 | 2.65 | 4.42 | 2.60 × 10−3 |
| Fbn1 | Fibrillin 1 | 8.27 | 30.34 | 3.67 | 1.55 × 10−2 |
| Col18a1 | Collagen, type XVIII, alpha 1 | 21.09 | 71.66 | 3.40 | 2.79 × 10−2 |
| Ltbp2 | Latent transforming growth factor beta binding protein 2 | 6.54 | 21.56 | 3.30 | 2.70 × 10−2 |
| Ltbp1 | Latent transforming growth factor beta binding protein 1 | 6.54 | 17.41 | 2.66 | 5.22 × 10−3 |
| Gene Code | Gene Name | Wild Type 6 h FPKM | FosBcKO 6 h FPKM | Fold Change | FDR |
|---|---|---|---|---|---|
| Dio3 | Deiodinase, iodothyronine type III | 0.39 | 10.75 | 27.00 | 4.19 × 10−24 |
| Pebp4 | Phosphatidylethanolamine binding protein 4 | 0.14 | 4.11 | 26.88 | 4.75 × 10−12 |
| Clec14a | C-type lectin domain family 14, member a | 0.09 | 2.48 | 24.75 | 2.60 × 10−8 |
| Edn3 | Endothelin 3 | 0.22 | 3.76 | 17.00 | 1.78 × 10−8 |
| Foxi3 | Forkhead box i3 | 0.83 | 12.46 | 14.80 | 1.39 × 10−10 |
| Kcnj13 | Potassium inwardly rectifying channel, subfamily J, member 13 | 0.74 | 11.13 | 14.74 | 6.61 × 10−8 |
| Defb9 | Defensin beta 9 | 0.80 | 12.02 | 14.46 | 2.14 × 10−7 |
| Fbn2 | Fibrillin 2 | 0.61 | 8.81 | 14.27 | 3.60 × 10−14 |
| Dsg2 | Desmoglein 2 | 0.36 | 5.07 | 13.78 | 6.46 × 10−24 |
| Gpx3 | Glutathione peroxidase 3 | 55.77 | 735.90 | 13.19 | 1.38 × 10−14 |
| Gene Code | Gene Name | Wild Type 6 h FPKM | FosBcKO 6 h FPKM | Fold Change | FDR |
|---|---|---|---|---|---|
| Slc29a4 | Solute carrier family 29 | 6.89 | 0.87 | −7.86 | 2.95 × 10−4 |
| Sox30 | SRY box 30 | 3.17 | 0.43 | −7.24 | 1.17 × 10−3 |
| Gjb3 | Gap junction protein, beta 3 | 4.37 | 0.62 | −7.06 | 2.73 × 10−2 |
| Cdcp3 | CUB domain containing protein 3 | 8.71 | 1.60 | −5.43 | 1.54 × 10−8 |
| Gjb4 | Gap junction protein, beta 4 | 8.02 | 1.52 | −5.29 | 3.36 × 10−3 |
| H1f8 | H1.8 linker histone | 35.69 | 6.96 | −5.13 | 1.18 × 10−3 |
| Cdsn | Corneodesmosin | 3.16 | 0.75 | −4.21 | 1.12 × 10−4 |
| Alpk2 | Alpha-kinase 2 | 3.24 | 0.78 | −4.16 | 5.48 × 10−3 |
| Asb5 | Ankyrin repeat and SOCs box-containing 5 | 6.02 | 1.45 | −4.16 | 5.48 × 10−3 |
| Cd109 | CD109 antigen | 4.25 | 1.06 | −4.01 | 1.71 × 10−2 |
| Gene Code | Gene Name | Wild Type 6 h FPKM | FosBcKO 6 h FPKM | Fold Change | FDR |
|---|---|---|---|---|---|
| Fbn2 | Fibrillin 2 | 0.61 | 8.81 | 14.27 | 3.60 × 10−14 |
| Fbn1 | Fibrillin 1 | 8.52 | 105.46 | 12.38 | 6.38 × 10−11 |
| Col18a1 | Collagen, type XVIII, alpha 1 | 21.68 | 233.53 | 10.77 | 1.43 × 10−9 |
| Ltbp2 | Latent transforming growth factor beta binding protein 2 | 6.72 | 72.04 | 10.71 | 2.55 × 10−10 |
| Nid2 | Nidogen 2 | 1.59 | 16.90 | 10.58 | 4.08 × 10−10 |
| Efemp1 | Epidermal growth factor-containing fibulin-like extracellular matrix protein 1 | 18.34 | 135.34 | 7.37 | 4.46 × 10−12 |
| Prelp | Proline arginine-rich end leucine-rich repeat | 14.44 | 83.96 | 5.81 | 3.11 × 10−11 |
| Col9a2 | Collagen, type IX, alpha 2 | 6.41 | 34.69 | 5.41 | 1.78 × 10−8 |
| Hmcn1 | Hemicentin 1 | 0.95 | 4.20 | 4.41 | 1.09 × 10−10 |
| Ltbp1 | Latent transforming growth factor beta binding protein 1 | 6.75 | 24.79 | 3.67 | 1.32 × 10−10 |
| Megf | Multiple EGF-like domains 6 | 1.27 | 4.39 | 3.45 | 4.86 × 10−5 |
| Loxl1 | Lysyl oxidase-like 1 | 26.12 | 64.47 | 2.47 | 6.21 × 10−4 |
| Ltbp3 | Latent transforming growth factor beta binding protein 3 | 23.90 | 52.78 | 2.21 | 1.58 × 10−4 |
| Ctsd | Cathepsin D | 31.84 | 70.16 | 2.20 | 4.60 × 10−5 |
| Timp3 | Tissue inhibitor of metalloproteinase 3 | 94.83 | 198.21 | 2.09 | 1.40 × 10−2 |
| Tgfb3 | Transforming growth factor, beta 3 | 2.83 | 5.74 | 2.03 | 1.21 × 10−2 |
| Agrn | Agrin | 8.37 | 16.96 | 2.03 | 1.42 × 10−3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Novo, S.G.; Faranda, A.P.; Shihan, M.H.; Wang, Y.; Garg, A.; Duncan, M.K. The Immediate Early Response of Lens Epithelial Cells to Lens Injury. Cells 2022, 11, 3456. https://doi.org/10.3390/cells11213456
Novo SG, Faranda AP, Shihan MH, Wang Y, Garg A, Duncan MK. The Immediate Early Response of Lens Epithelial Cells to Lens Injury. Cells. 2022; 11(21):3456. https://doi.org/10.3390/cells11213456
Chicago/Turabian StyleNovo, Samuel G., Adam P. Faranda, Mahbubul H. Shihan, Yan Wang, Ananya Garg, and Melinda K. Duncan. 2022. "The Immediate Early Response of Lens Epithelial Cells to Lens Injury" Cells 11, no. 21: 3456. https://doi.org/10.3390/cells11213456