
Nogo-A Regulates the Fate of Human Dental Pulp Stem Cells toward Osteogenic, Adipogenic, and Neurogenic Differentiation


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Data and Code Availability
2.2. Collection of Human Tissues and Cells
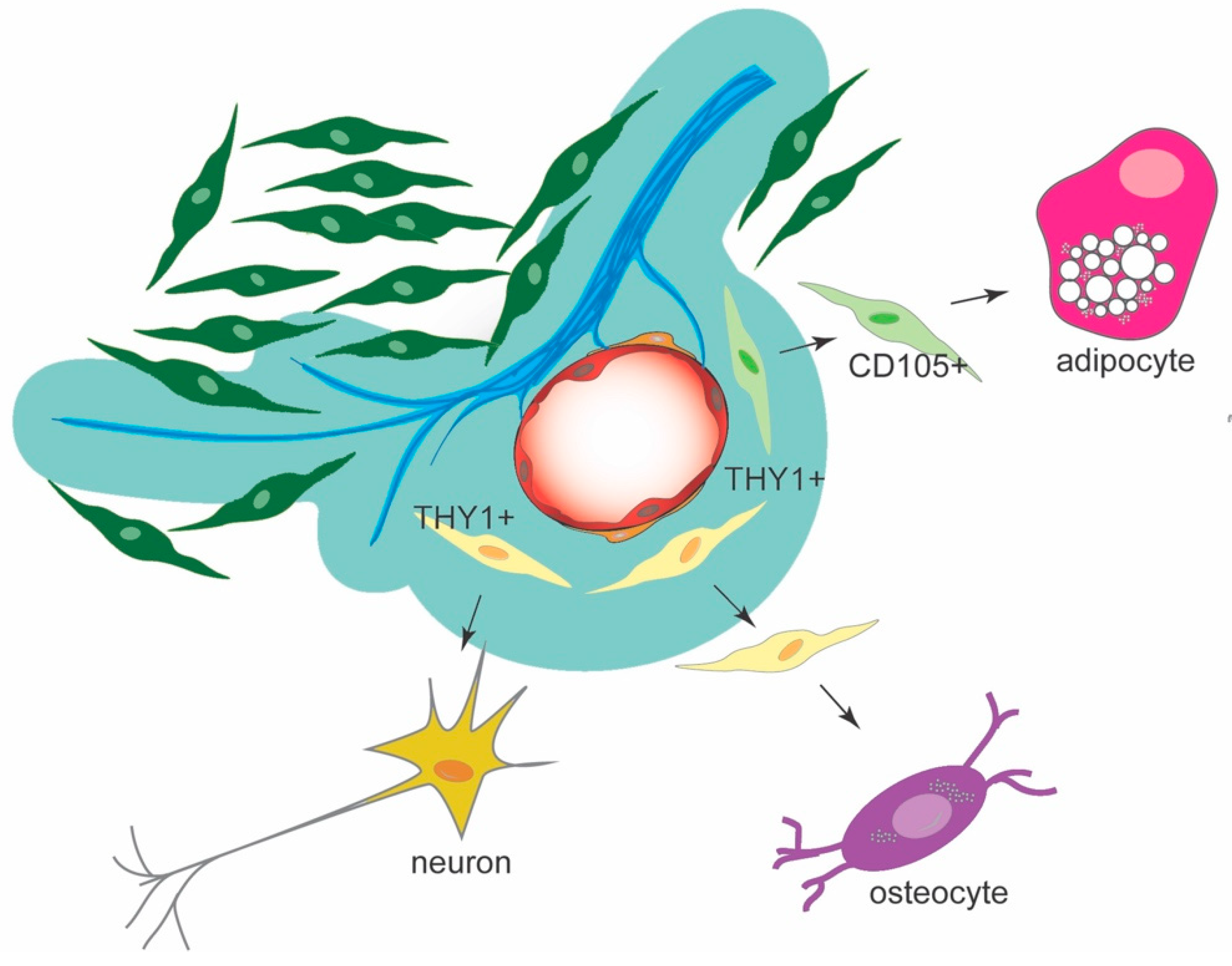
2.3. Cell Cultures and Differentiation Assays
2.4. Staining
2.5. RNA Extraction, Reverse Transcription and Quantitative Real-Time PCR (qRT-PCR)
2.6. Statistical Analysis
3. Results
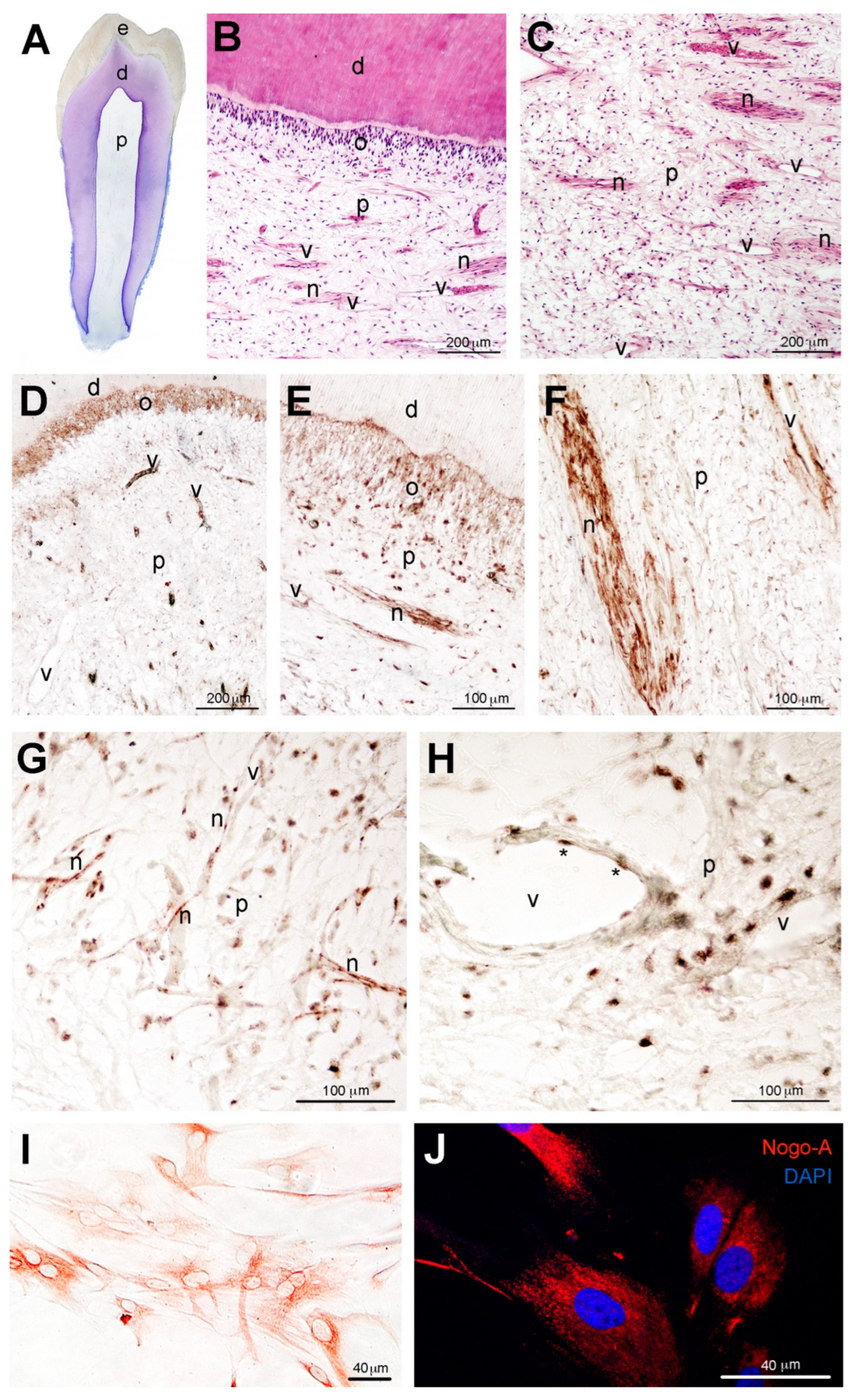
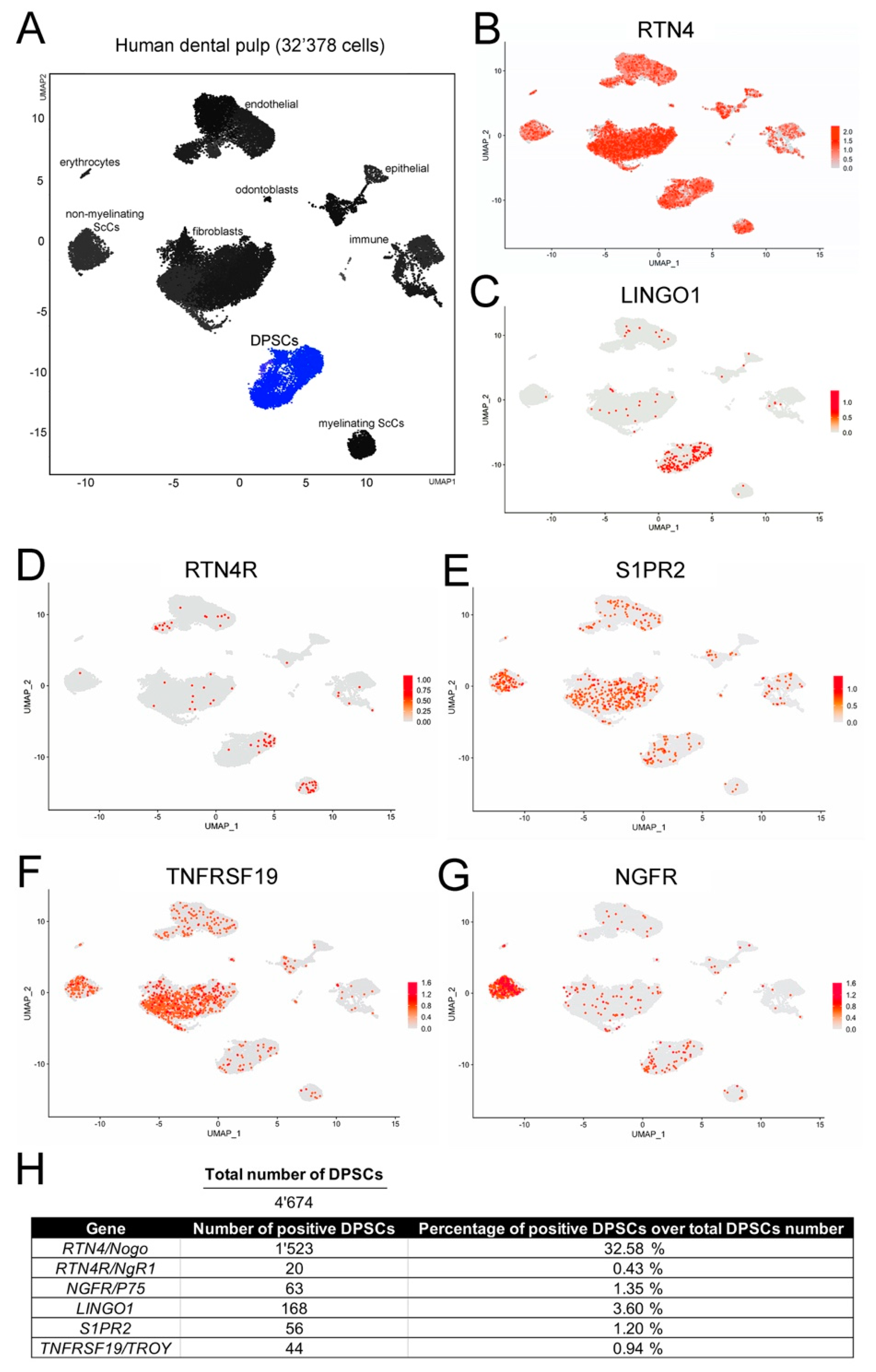
3.1. Localization of the Nogo-A Protein in Human Dental Pulp Cells
3.2. Nogo-A Effects on the In Vitro Osteogenic, Adipogenic, and Neurogenic Differentiation Potential of hDPSCs
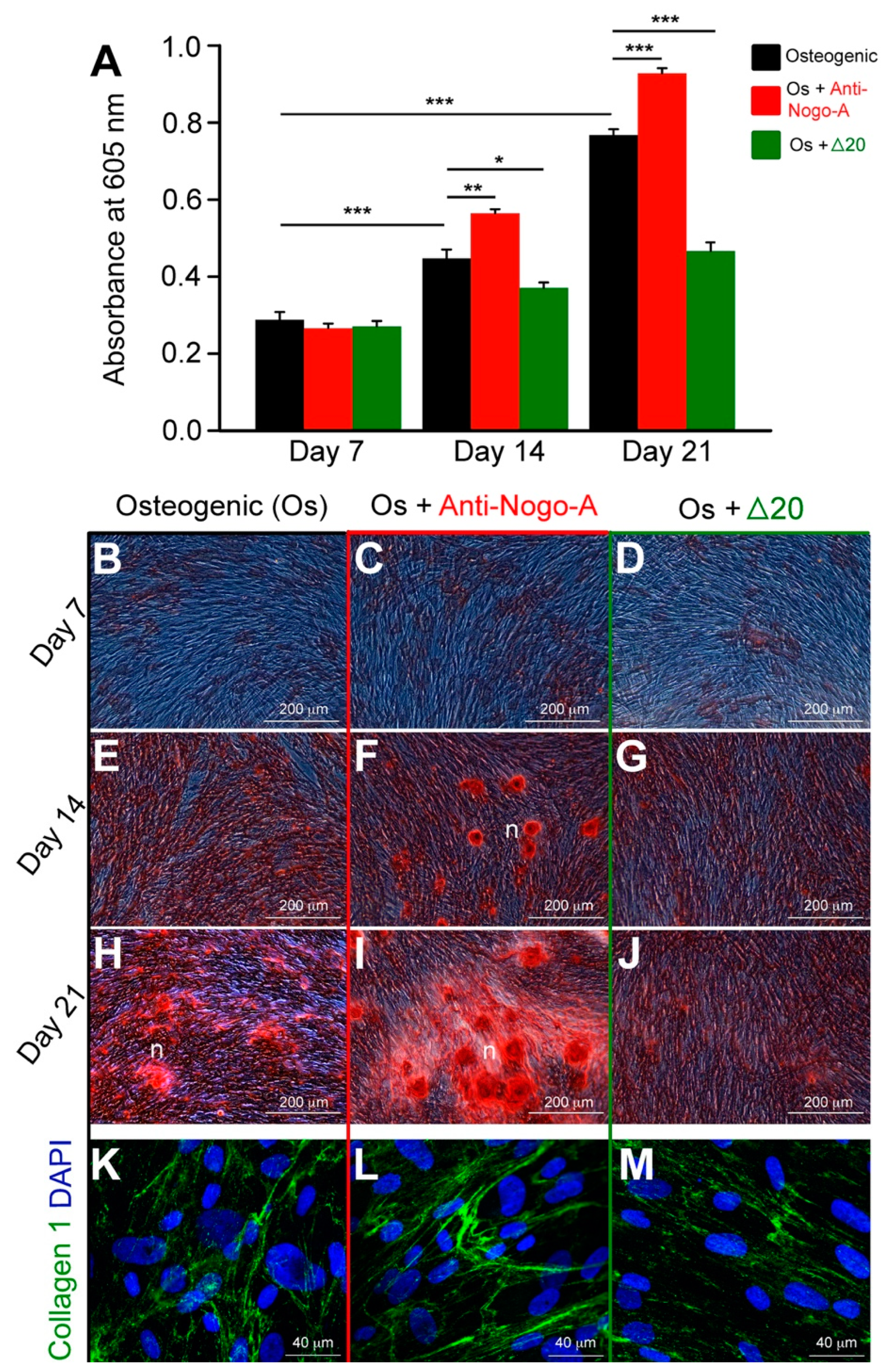
3.2.1. The Anti-Nogo-A Antibody Accelerates the Osteogenic Differentiation of hDPSCs
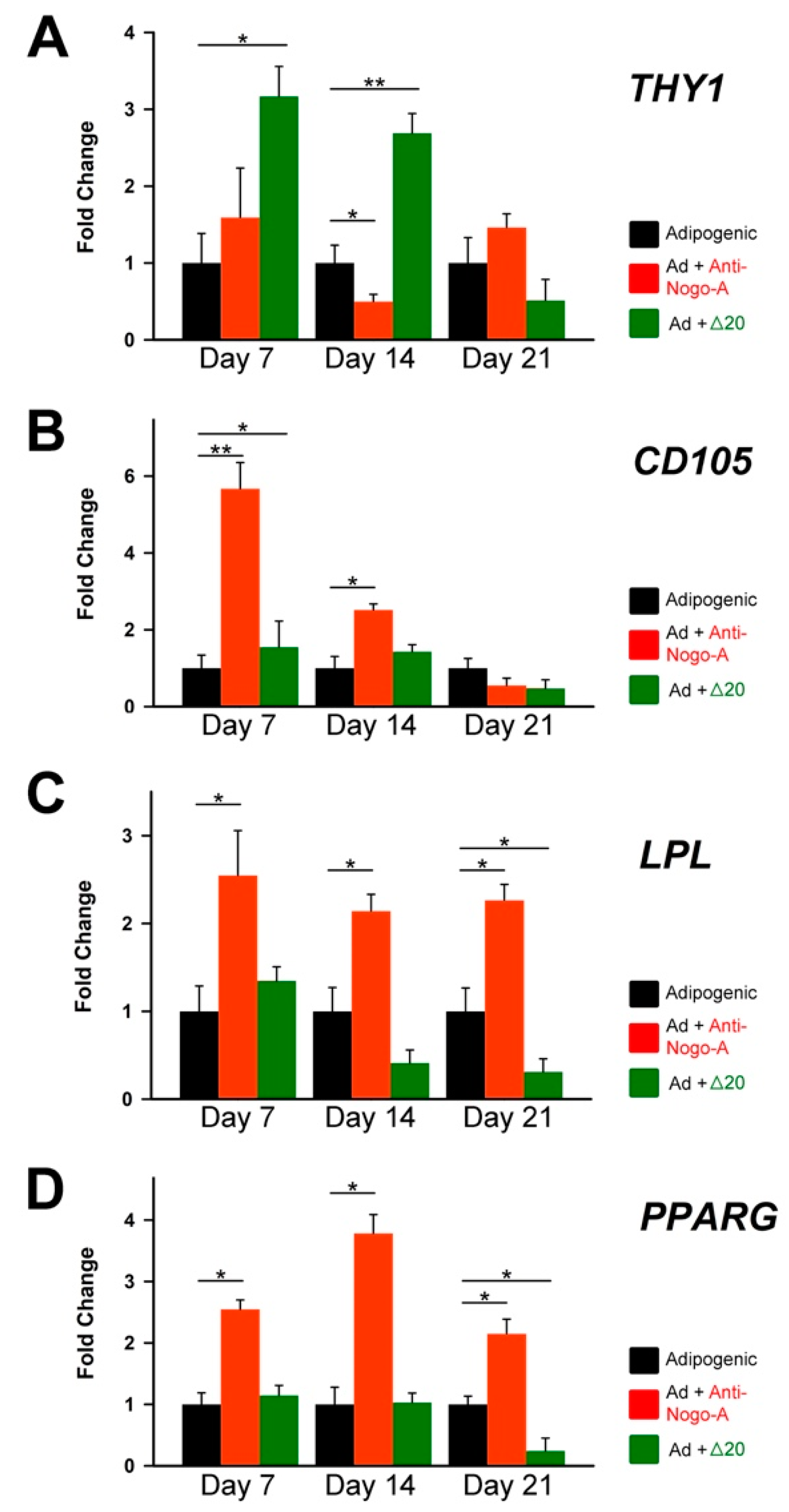
3.2.2. The Anti-Nogo-A Antibody Accelerates the Adipogenic Differentiation of hDPCSs
3.2.3. The Anti-Nogo-A Antibody Accelerates the Neurogenic Differentiation of hDPSCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pera, M.F.; Tam, P.P. Extrinsic regulation of pluripotent stem cells. Nature 2010, 465, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Nombela-Arrieta, C.; Ritz, J.; Silberstein, L.E. The elusive nature and function of mesenchymal stem cells. Nat. Rev. Mol. Cell Biol. 2011, 12, 126–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Signer, R.A.; Morrison, S.J. Mechanisms that regulate stem cell aging and life span. Cell Stem Cell. 2013, 12, 152–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, J.M.; Watt, F.M. Diverse mechanisms for endogenous regeneration and repair in mammalian organs. Nature 2018, 557, 322–328. [Google Scholar] [CrossRef] [Green Version]
- Mitsiadis, T.A.; Barrandon, O.; Rochat, A.; Barrandon, Y.; De Bari, C. Stem cell niches in mammals. Exp. Cell Res. 2007, 313, 3377–3385. [Google Scholar] [CrossRef]
- Morrison, S.J.; Scadden, D.T. The bone marrow niche for haematopoietic stem cells. Nature 2014, 505, 327–334. [Google Scholar] [CrossRef] [Green Version]
- Bautch, V.L. Stem cells and the vasculature. Nat. Med. 2011, 17, 1437–1443. [Google Scholar] [CrossRef]
- Pagella, P.; Neto, E.; Lamghari, M.; Mitsiadis, T.A. Investigation of orofacial stem cell niches and their innervation through microfluidic devices. Eur. Cell Mater. 2015, 29, 213–223. [Google Scholar] [CrossRef]
- Watt, F.M.; Hogan, B.L. Out of Eden: Stem cells and their niches. Science 2000, 287, 1427–1430. [Google Scholar] [CrossRef] [Green Version]
- Augello, A.; Kurth, T.B.; De Bari, C. Mesenchymal stem cells: A perspective from in vitro cultures to in vivo migration and niches. Eur. Cell Mater. 2010, 20, 121–133. [Google Scholar] [CrossRef]
- Pagella, P.; de Vargas Roditi, L.; Stadlinger, B.; Moor, A.E.; Mitsiadis, T.A. A single-cell atlas of human teeth. iScience 2021, 24, 102405. [Google Scholar] [CrossRef] [PubMed]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gronthos, S.; Brahim, J.; Li, W.; Fisher, L.W.; Cherman, N.; Boyde, A.; DenBesten, P.; Robey, P.G.; Shi, S. Stem cell properties of human dental pulp stem cells. J. Dent. Res. 2002, 81, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Morrison, S.J.; Wright, D.E.; Cheshier, S.H.; Weissman, I.L. Hematopoietic stem cells: Challenges to expectations. Curr. Opin. Immunol. 1997, 9, 216–221. [Google Scholar] [CrossRef]
- Jimenez-Rojo, L.; Pagella, P.; Harada, H.; Mitsiadis, T.A. Dental Epithelial Stem Cells as a Source for Mammary Gland Regeneration and Milk Producing Cells In Vivo. Cells 2019, 8, 1302. [Google Scholar] [CrossRef] [Green Version]
- Orsini, G.; Pagella, P.; Putignano, A.; Mitsiadis, T.A. Novel Biological and Technological Platforms for Dental Clinical Use. Front. Physiol. 2018, 9, 1102. [Google Scholar] [CrossRef]
- Miura, M.; Gronthos, S.; Zhao, M.; Lu, B.; Fisher, L.W.; Robey, P.G.; Shi, S. SHED: Stem cells from human exfoliated deciduous teeth. Proc. Natl. Acad. Sci. USA 2003, 100, 5807–5812. [Google Scholar] [CrossRef] [Green Version]
- Sharpe, P.T. Dental mesenchymal stem cells. Development 2016, 143, 2273–2280. [Google Scholar] [CrossRef] [Green Version]
- Pagella, P.; de Vargas Roditi, L.; Stadlinger, B.; Moor, A.E.; Mitsiadis, T.A. Notch signaling in the dynamics of perivascular stem cells and their niches. Stem Cells Transl. Med. 2021, 10, 1433–1445. [Google Scholar] [CrossRef]
- Pagella, P.; Nombela-Arrieta, C.; Mitsiadis, T.A. Distinct Expression Patterns of Cxcl12 in Mesenchymal Stem Cell Niches of Intact and Injured Rodent Teeth. Int. J. Mol. Sci. 2021, 22, 3024. [Google Scholar] [CrossRef]
- Shi, S.; Gronthos, S. Perivascular niche of postnatal mesenchymal stem cells in human bone marrow and dental pulp. J. Bone Miner. Res. 2003, 18, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iohara, K.; Zayed, M.; Takei, Y.; Watanabe, H.; Nakashima, M. Treatment of Pulpectomized Teeth With Trypsin Prior to Transplantation of Mobilized Dental Pulp Stem Cells Enhances Pulp Regeneration in Aged Dogs. Front. Bioeng. Biotechnol. 2020, 8, 983. [Google Scholar] [CrossRef] [PubMed]
- Orsini, G.; Pagella, P.; Mitsiadis, T.A. Modern Trends in Dental Medicine: An Update for Internists. Am. J. Med. 2018, 131, 1425–1430. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Bartold, P.M.; Miura, M.; Seo, B.M.; Robey, P.G.; Gronthos, S. The efficacy of mesenchymal stem cells to regenerate and repair dental structures. Orthod. Craniofac. Res. 2005, 8, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Thesleff, I. From understanding tooth development to bioengineering of teeth. Eur. J. Oral. Sci. 2018, 126, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Chen, J.; Liu, S.; Jin, Y. Stem cell-based bone and dental regeneration: A view of microenvironmental modulation. Int. J. Oral. Sci. 2019, 11, 23. [Google Scholar] [CrossRef] [Green Version]
- Mattei, V.; Santacroce, C.; Tasciotti, V.; Martellucci, S.; Santilli, F.; Manganelli, V.; Piccoli, L.; Misasi, R.; Sorice, M.; Garofalo, T. Role of lipid rafts in neuronal differentiation of dental pulp-derived stem cells. Exp. Cell Res. 2015, 339, 231–240. [Google Scholar] [CrossRef]
- Delle Monache, S.; Martellucci, S.; Clementi, L.; Pulcini, F.; Santilli, F.; Mei, C.; Piccoli, L.; Angelucci, A.; Mattei, V. In vitro conditioning determines the capacity of dental pulp stem cells to function as pericyte-like cells. Stem Cells Dev. 2019, 28, 695–706. [Google Scholar] [CrossRef]
- Monterubbianesi, R.; Bencun, M.; Pagella, P.; Woloszyk, A.; Orsini, G.; Mitsiadis, T.A. A comparative in vitro study of the osteogenic and adipogenic potential of human dental pulp stem cells, gingival fibroblasts and foreskin fibroblasts. Sci. Rep. 2019, 9, 1761. [Google Scholar] [CrossRef] [Green Version]
- Bluteau, G.; Luder, H.U.; De Bari, C.; Mitsiadis, T.A. Stem cells for tooth engineering. Eur. Cell Mater. 2008, 16, 1–9. [Google Scholar] [CrossRef]
- Yasui, T.; Mabuchi, Y.; Toriumi, H.; Ebine, T.; Niibe, K.; Houlihan, D.D.; Morikawa, S.; Onizawa, K.; Kawana, H.; Akazawa, C.; et al. Purified Human Dental Pulp Stem Cells Promote Osteogenic Regeneration. J. Dent. Res. 2016, 95, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Rafiee, F.; Pourteymourfard-Tabrizi, Z.; Mahmoudian-Sani, M.R.; Mehri-Ghahfarrokhi, A.; Soltani, A.; Hashemza-deh-Chaleshtori, M.; and Jami, M.S. Differentiation of dental pulp stem cells into neuron-like cells. Int. J. Neurosci. 2020, 130, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Arimura, Y.; Shindo, Y.; Yamanaka, R.; Mochizuki, M.; Hotta, K.; Nakahara, T.; Ito, E.; Yoshioka, T.; Oka, K. Peripheral-neuron-like properties of differentiated human dental pulp stem cells (hDPSCs). PLoS ONE 2021, 16, e0251356. [Google Scholar] [CrossRef] [PubMed]
- Luzuriaga, J.; Irurzun, J.; Irastorza, I.; Unda, F.; Ibarretxe, G.; Pineda, J.R. Vasculogenesis from human dental pulp stem cells grown in matrigel with fully defined serum-free culture media. Biomedicines 2020, 8, 483. [Google Scholar] [CrossRef] [PubMed]
- Noda, S.; Kawashima, N.; Yamamoto, M.; Hashimoto, K.; Nara, K.; Sekiya, I.; Okiji, T. Effect of cell culture density on dental pulp-derived mesenchymal stem cells with reference to osteogenic. Differ. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Delle Monache, S.; Pulcini, F.; Santilli, F.; Martellucci, S.; Santacroce, C.; Fabrizi, J.; Angelucci, A.; Sorice, M.; Mattei, V. Hypoxia Induces DPSC Differentiation versus a Neurogenic Phenotype by the Paracrine Mechanism. Biomedicines 2022, 10, 1056. [Google Scholar] [CrossRef]
- Labedz-Maslowska, A.; Bryniarska, N.; Kubiak, A.; Kaczmarzyk, T.; Sekula-Stryjewska, M.; Noga, S.; Boruczkowski, D.; Madeja, Z.; Zuba-Surma, E. Multilineage Differentiation Potential of Human Dental Pulp Stem Cells-Impact of 3D and Hypoxic Environment on Osteogenesis in Vitro. Int. J. Mol. Sci. 2020, 21, 6172. [Google Scholar] [CrossRef]
- Mitsiadis, T.A.; Orsini, G.; Jimenez-Rojo, L. Stem cell-based approaches in dentistry. Eur. Cell Mater. 2015, 30, 248–257. [Google Scholar] [CrossRef]
- Madl, C.M.; Heilshorn, S.C.; Blau, H.M. Bioengineering strategies to accelerate stem cell therapeutics. Nature 2018, 557, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Nery, A.A.; Nascimento, I.C.; Glaser, T.; Bassaneze, V.; Krieger, J.E.; Ulrich, H. Human mesenchymal stem cells: From immunophenotyping by flow cytometry to clinical applications. Cytometry A 2013, 83, 48–61. [Google Scholar] [CrossRef]
- Zayed, M.; Iohara, K.; Watanabe, H.; Ishikawa, M.; Tominaga, M.; Nakashima, M. Characterization of stable hypoxia-preconditioned dental pulp stem cells compared with mobilized dental pulp stem cells for application for pulp regenerative therapy. Stem Cell Res. Ther. 2021, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.; Yuan, K.; Lin, W.; Niu, C.; Ma, R.; Huang, Z. Comparative characterization of mesenchymal stem cells from human dental pulp and adipose tissue for bone regeneration potential. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1577–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iohara, K.; Utsunomiya, S.; Kohara, S.; Nakashima, M. Allogeneic transplantation of mobilized dental pulp stem cells with the mismatched dog leukocyte antigen type is safe and efficacious for total pulp regeneration. Stem Cell Res. Ther. 2018, 9, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, M.; Iohara, K.; Murakami, M.; Nakamura, H.; Sato, Y.; Ariji, Y.; Matsushita, K. Pulp regeneration by transplantation of dental pulp stem cells in pulpitis: A pilot clinical study. Stem Cell Res. Ther. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- d’Aquino, R.; De Rosa, A.; Lanza, V.; Tirino, V.; Laino, L.; Graziano, A.; Desiderio, V.; Laino, G.; Papaccio, G. Human mandible bone defect repair by the grafting of dental pulp stem/progenitor cells and collagen sponge biocomplexes. Eur. Cell Mater. 2009, 18, 75–83. [Google Scholar] [CrossRef]
- Mason, C.; Dunnill, P. A brief definition of regenerative medicine. Regen Med. 2008, 3, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Angelova Volponi, A.; Zaugg, L.K.; Neves, V.; Liu, Y.; Sharpe, P.T. Tooth Repair and Regeneration. Curr. Oral Health Rep. 2018, 5, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.S.; Huber, A.B.; van der Haar, M.E.; Frank, M.; Schnell, L.; Spillmann, A.A.; Christ, F.; Schwab, M.E. Nogo-A is a myelin-associated neurite outgrowth inhibitor and an antigen for monoclonal antibody IN-1. Nature 2000, 403, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Delekate, A.; Zagrebelsky, M.; Kramer, S.; Schwab, M.E.; Korte, M. NogoA restricts synaptic plasticity in the adult hippocampus on a fast time scale. Proc. Natl. Acad. Sci. USA 2011, 108, 2569–2574. [Google Scholar] [CrossRef] [Green Version]
- Kempf, A.; Tews, B.; Arzt, M.E.; Weinmann, O.; Obermair, F.J.; Pernet, V.; Zagrebelsky, M.; Delekate, A.; Iobbi, C.; Zemmar, A.; et al. The sphingolipid receptor S1PR2 is a receptor for Nogo-a repressing synaptic plasticity. PLoS Biol. 2014, 12, e1001763. [Google Scholar] [CrossRef] [Green Version]
- Grandpre, T.; Strittmatter, S.M. Nogo: A molecular determinant of axonal growth and regeneration. Neuroscientist 2001, 7, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Fournier, A.E.; GrandPre, T.; Strittmatter, S.M. Identification of a receptor mediating Nogo-66 inhibition of axonal regeneration. Nature 2001, 409, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Park, J.B.; Yiu, G.; Kaneko, S.; Wang, J.; Chang, J.; He, Z. A TNF receptor family member, TROY, is a coreceptor with Nogo receptor in mediating the inhibitory activity of myelin inhibitors. Neuron 2005, 45, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Shao, Z.; Browning, J.L.; Lee, X.; Scott, M.L.; Shulga-Morskaya, S.; Allaire, N.; Thill, G.; Levesque, M.; Sah, D.; McCoy, J.M.; et al. TAJ/TROY, an orphan TNF receptor family member, binds Nogo-66 receptor 1 and regulates axonal regeneration. Neuron 2005, 45, 353–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, S.; Lee, X.; Shao, Z.; Thill, G.; Ji, B.; Relton, J.; Levesque, M.; Allaire, N.l.; Perrin, S.; Sands, B.; et al. LINGO-1 is a component of the Nogo-66 receptor/p75 signaling complex. Nat. Neurosci. 2004, 7, 221–228. [Google Scholar] [CrossRef]
- Bronfman, F.C.; Fainzilber, M. Multi-tasking by the p75 neurotrophin receptor: Sortilin things out? EMBO Rep. 2004, 5, 867–871. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, T.; Tohyama, M. The p75 receptor acts as a displacement factor that releases Rho from Rho-GDI. Nat. Neurosci. 2003, 6, 461–467. [Google Scholar] [CrossRef]
- Wang, K.C.; Kim, J.A.; Sivasankaran, R.; Segal, R.; He, Z. P75 interacts with the Nogo receptor as a co-receptor for Nogo, MAG and OMgp. Nature 2002, 420, 74–78. [Google Scholar] [CrossRef]
- Seiler, S.; Di Santo, S.; Widmer, H.R. Non-canonical actions of Nogo-A and its receptors. Biochem. Pharmacol. 2016, 100, 28–39. [Google Scholar] [CrossRef]
- Wu, Q.; Zhang, H.; Nie, H.; Zeng, Z. Anti NogoA antibody promotes brain function recovery after cardiopulmonary resuscitation in rats by reducing apoptosis. Mol. Med. Rep. 2020, 21, 77–88. [Google Scholar] [CrossRef]
- Guo, M.F.; Zhang, H.Y.; Zhang, P.J.; Liu, X.Q.; Song, L.J.; Wei, W.Y.; Wang, Y.Y.; Mu, B.T.; Chai, Z.; Yu, J.Z.; et al. Fasudil reduces beta-amyloid levels and neuronal apoptosis in APP/PS1 transgenic mice via inhibition of the Nogo-A/NgR/RhoA signaling axis. J. Integr. Neurosci. 2020, 19, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Shu, H.; Yang, T.; Huang, H.; Li, S.; Zhao, Z.; Kuang, Y. Upregulated expression of Nogo-A and NgR in an experimental model of focal microgyria regulates the migration, proliferation and self-renewal of subventricular zone neural progenitors. Biochem. Biophys. Res. Commun. 2016, 473, 482–489. [Google Scholar] [CrossRef] [PubMed]
- Ullah, H.A.; Elfadl, A.K.; Park, S.; Kim, Y.D.; Chung, M.J.; Son, J.Y.; Yun, H.H.; Park, J.M.; Yim, J.H.; Jung, S.J.; et al. Nogo-A Is Critical for Pro-Inflammatory Gene Regulation in Myocytes and Macrophages. Cells 2021, 10, 282. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Cui, Y.; Li, Y.; Mi, Y. A Novel Role of Nogo Proteins: Regulating Macrophages in Inflammatory Disease. Cell Mol. Neurobiol. 2021, 42, 2439–2448. [Google Scholar] [CrossRef]
- Wälchli, T.; Pernet, V.; Weinmann, O.; Shiu, J.Y.; Guzik-Kornacka, A.; Decrey, G.; Yüksel, D.; Schneider, H.; Vogel, J.; Ingber, D.E.; et al. Nogo-A is a negative regulator of CNS angiogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, E1943–E1952. [Google Scholar] [CrossRef] [Green Version]
- Rust, R.; Grönnert, L.; Gantner, C.; Enzler, A.; Mulders, G.; Weber, R.Z.; Siewert, A.; Limasale, Y.D.; Meinhardt, A.; Maurer, M.A.; et al. Nogo-A targeted therapy promotes vascular repair and functional recovery following stroke. Proc. Natl. Acad. Sci. USA 2019, 116, 14270–14279. [Google Scholar] [CrossRef] [Green Version]
- Xiao, P.; Gu, J.; Xu, W.; Niu, X.; Zhang, J.; Li, J.; Chen, Y.; Pei, Z.; Zeng, J.; Xing, S. RTN4/Nogo-A-S1PR2 negatively regulates angiogenesis and secondary neural repair through enhancing vascular autophagy in the thalamus after cerebral cortical infarction. Autophagy 2022, 1–20. [Google Scholar] [CrossRef]
- Wang, J.L.; Chen, W.G.; Zhang, J.J.; Xu, C.J. Nogo-A-Delta20/EphA4 interaction antagonizes apoptosis of neural stem cells by integrating p38 and JNK MAPK signaling. J. Mol. Histol. 2021, 52, 521–537. [Google Scholar] [CrossRef]
- Ai, C.; Zhou, Y.; Pu, K.; Yang, Y.; Zhou, Y. Nogo-A/NgR signaling regulates stemness in cancer stem-like cells derived from U87MG glioblastoma cells. Oncol. Lett. 2022, 24, 230. [Google Scholar] [CrossRef]
- Boghdadi, A.G.; Spurrier, J.; Teo, L.; Li, M.; Skarica, M.; Cao, B.; Kwan, W.C.; Merson, T.D.; Nilsson, S.K.; Sestan, N.; et al. NogoA-expressing astrocytes limit peripheral macrophage infiltration after ischemic brain injury in primates. Nat. Commun. 2021, 12, 6906. [Google Scholar] [CrossRef]
- Rolando, C.; Parolisi, R.; Boda, E.; Schwab, M.E.; Rossi, F.; Buffo, A. Distinct roles of Nogo-a and Nogo receptor 1 in the homeostatic regulation of adult neural stem cell function and neuroblast migration. J. Neurosci. 2012, 32, 17788–17799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucher, K.; Johns, D.; Maier, D.; Abel, R.; Badke, A.; Baron, H.; Thietje, R.; Casha, S.; Meindl, R.; Gomez-Mancilla, B.; et al. First-in-Man Intrathecal Application of Neurite Growth-Promoting Anti-Nogo-A Antibodies in Acute Spinal Cord Injury. Neurorehabil. Neural Repair 2018, 32, 578–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Halabiah, H.; Delezoide, A.L.; Cardona, A.; Moalic, J.M.; Simonneau, M. Expression pattern of NOGO and NgR genes during human development. Gene Expr. Patterns 2005, 5, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Mitsiadis, T.A.; Dicou, E.; Joffre, A.; Magloire, H. Immunohistochemical localization of nerve growth factor (NGF) and NGF receptor (NGF-R) in the developing first molar tooth of the rat. Differentiation 1992, 49, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Mitsiadis, T.A.; Romeas, A.; Lendahl, U.; Sharpe, P.T.; Farges, J.C. Notch2 protein distribution in human teeth under normal and pathological conditions. Exp. Cell Res. 2003, 282, 101–109. [Google Scholar] [CrossRef]
- Oertle, T.; Van Der Haar, M.E.; Bandtlow, C.E.; Robeva, A.; Burfeind, P.; Buss, A.; Huber, A.B.; Simonen, M.; Schnell, L.; Brösamle, C.; et al. Nogo-A inhibits neurite outgrowth and cell spreading with three discrete regions. J. Neurosci. 2003, 23, 5393–5406. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Barry, F.P.; Murphy, J.M. Mesenchymal stem cells: Clinical applications and biological characterization. Int. J. Biochem. Cell Biol. 2004, 36, 568–584. [Google Scholar] [CrossRef]
- Cossu, G.; Birchall, M.; Brown, T.; De Coppi, P.; Culme-Seymour, E.; Gibbon, S.; Hitchcock, J.; Mason, C.; Montgomery, J.; Morris, S.; et al. Lancet Commission: Stem cells and regenerative medicine. Lancet Comm. Stem Cells Regen. Med. Lancet 2018, 391, 883–910. [Google Scholar] [CrossRef] [Green Version]
- Waters, S.L.; Schumacher, L.J.; El Haj, A.J. Regenerative medicine meets mathematical modelling: Developing symbiotic relationships. NPJ Regen. Med. 2021, 6, 24. [Google Scholar] [CrossRef]
- El Haj, A.J. Regenerative Medicine: “Are We There Yet?”. Tissue Eng. Part A 2019, 25, 1067–1071. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal stem cell perspective: Cell biology to clinical progress. NPJ Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toure, S.B.; Kleiderman, E.; Knoppers, B.M. Bridging stem cell research and medicine: A learning health system. Regen. Med. 2018, 13, 741–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gothard, D.; Smith, E.L.; Kanczler, J.M.; Rashidi, H.; Qutachi, O.; Henstock, J.R.; Rotherham, M.; El Haj, A.J.; Shakesheff, K.M.; Oreffo, R.O. Tissue engineered bone using select growth factors: A comprehensive review of animal studies and clinical translation studies in man. Eur. Cell Mater. 2014, 28, 166–207. [Google Scholar] [CrossRef] [PubMed]
- Iohara, K.; Murakami, M.; Takeuchi, N.; Osako, Y.; Ito, M.; Ishizaka, R.; Utunomiya, S.; Nakamura, H.; Matsushita, K.; Nakashima, M. A novel combinatorial therapy with pulp stem cells and granulocyte colony-stimulating factor for total pulp regeneration. Stem Cells Transl. Med. 2013, 2, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Pagella, P.; Cordiale, A.; Marconi, G.D.; Trubiani, O.; Rasponi, M.; Mitsiadis, T.A. Bioengineered tooth emulation systems for regenerative and pharmacological purposes. Eur. Cell Mater. 2021, 41, 502–516. [Google Scholar] [CrossRef] [PubMed]
- Jussila, M.; Thesleff, I. Signaling networks regulating tooth organogenesis and regeneration, and the specification of dental mesenchymal and epithelial cell lineages. Cold Spring Harb. Perspect Biol. 2012, 4, a008425. [Google Scholar] [CrossRef]
- Mitsiadis, T.A.; Rahiotis, C. Parallels between tooth development and repair: Conserved molecular mechanisms following carious and dental injury. J. Dent. Res. 2004, 83, 896–902. [Google Scholar] [CrossRef]
- Mitsiadis, T.A.; Feki, A.; Papaccio, G.; Caton, J. Dental pulp stem cells, niches, and notch signaling in tooth injury. Adv. Dent. Res. 2011, 23, 275–279. [Google Scholar] [CrossRef]
- Mitsiadis, T.A.; Magloire, H.; Pagella, P. Nerve growth factor signalling in pathology and regeneration of human teeth. Sci. Rep. 2017, 7, 1327. [Google Scholar] [CrossRef]
- Schwab, M.E.; Strittmatter, S.M. Nogo limits neural plasticity and recovery from injury. Curr. Opin. Neurobiol. 2014, 27, 53–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahl, A.S.; Omlor, W.; Rubio, J.C.; Chen, J.L.; Zheng, H.; Schröter, A.; Gullo, M.; Weinmann, O.; Kobayashi, K.; Helmchen, F.; et al. Neuronal repair. Asynchronous therapy restores motor control by rewiring of the rat corticospinal tract after stroke. Science 2014, 344, 1250–1255. [Google Scholar] [CrossRef] [PubMed]
- Freund, P.; Schmidlin, E.; Wannier, T.; Bloch, J.; Mir, A.; Schwab, M.E.; Rouiller, E.M. Nogo-A-specific antibody treatment enhances sprouting and functional recovery after cervical lesion in adult primates. Nat. Med. 2006, 12, 790–792. [Google Scholar] [CrossRef]
- Nagaraj, V.; Theis, T.; Johal, A.S.; Seth, A.; Gore, J.; Arsha, N.; Patel, M.; Hao, H.B.; Kurian, N.; Schachner, M. Application of Antibodies to Neuronally Expressed Nogo-A Increases Neuronal Survival and Neurite Outgrowth. Int. J. Mol. Sci. 2020, 21, 5417. [Google Scholar] [CrossRef]
- Tang, H.; Xu, Y.; Liu, L.; He, L.; Huang, J.; Pan, J.; He, W.; Wang, Y.; Yang, X.; Hou, X.; et al. Nogo-A/S1PR2 Signaling Pathway Inactivation Decreases Microvascular Damage and Enhances Microvascular Regeneration in PDMCI Mice. Neuroscience 2020, 449, 21–34. [Google Scholar] [CrossRef]
- Li, B.; Carey, M.; Workman, J.L. The role of chromatin during transcription. Cell 2007, 128, 707–719. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Rajewsky, N. The evolution of gene regulation by transcription factors and microRNAs. Nat. Rev. Genet. 2007, 8, 93–103. [Google Scholar] [CrossRef]
- Basu, A.; Tiwari, V.K. Epigenetic reprogramming of cell identity: Lessons from development for regenerative medicine. Clin. Epigenetics 2021, 13, 144. [Google Scholar] [CrossRef]
- Yang, J.; Zhan, X.Z.; Malola, J.; Li, Z.Y.; Pawar, J.S.; Zhang, H.T.; Zha, Z.G. The multiple roles of Thy-1 in cell differentiation and regeneration. Differentiation 2020, 113, 38–48. [Google Scholar] [CrossRef]
- Saalbach, A.; Anderegg, U. Thy-1: More than a marker for mesenchymal stromal cells. FASEB J. 2019, 33, 6689–6696. [Google Scholar] [CrossRef]
- Moraes, D.A.; Sibov, T.T.; Pavon, L.F.; Alvim, P.Q.; Bonadio, R.S.; Da Silva, J.R.; Pic-Taylor, A.; Toledo, O.A.; Marti, L.C.; Azevedo, R.B.; et al. A reduction in CD90 (THY-1) expression results in increased differentiation of mesenchymal stromal cells. Stem Cell Res. Ther. 2016, 7, 97. [Google Scholar] [CrossRef] [Green Version]
- Chamberlain, G.; Fox, J.; Ashton, B.; Middleton, J. Concise review: Mesenchymal stem cells: Their phenotype, differentiation capacity, immunological features, and potential for homing. Stem Cells 2007, 25, 2739–2749. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.S.; Xin, Z.C.; Dai, J.; Lue, T.F. Commonly used mesenchymal stem cell markers and tracking labels: Limitations and challenges. Histol. Histopathol. 2013, 28, 1109–1116. [Google Scholar] [CrossRef]
- Komori, T. Regulation of osteoblast differentiation by Runx2. Adv. Exp. Med. Biol. 2010, 658, 43–49. [Google Scholar] [CrossRef]
- Kawane, T.; Qin, X.; Jiang, Q.; Miyazaki, T.; Komori, H.; Yoshida, C.A.; Matsuura-Kawata, V.K.D.S.; Sakane, C.; Matsuo, Y.; Nagai, K.; et al. Runx2 is required for the proliferation of osteoblast progenitors and induces proliferation by regulating Fgfr2 and Fgfr3. Sci. Rep. 2018, 8, 13551. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, A.; Srinaath, N.; Rohini, M.; Selvamurugan, N. Regulation of Runx2 by MicroRNAs in osteoblast differentiation. Life Sci. 2019, 232, 116676. [Google Scholar] [CrossRef]
- Zhang, C. Transcriptional regulation of bone formation by the osteoblast-specific transcription factor Osx. J. Orthop Surg Res. 2010, 5, 37. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, K.; Zhou, X.; Kunkel, G.; Zhang, Z.; Deng, J.M.; Behringer, R.R.; De Crombrugghe, B. The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation. Cell 2002, 108, 17–29. [Google Scholar] [CrossRef] [Green Version]
- Stefkova, K.; Prochazkova, J.; Pachernik, J. Alkaline phosphatase in stem cells. Stem Cells Int. 2015, 2015, 628368. [Google Scholar] [CrossRef] [Green Version]
- Beck, G.R., Jr.; Zerler, B.; Moran, E. Phosphate is a specific signal for induction of osteopontin gene expression. Proc. Natl. Acad. Sci. USA 2000, 97, 8352–8357. [Google Scholar] [CrossRef]
- Sadie-Van Gijsen, H.; Hough, F.S.; Ferris, W.F. Determinants of bone marrow adiposity: The modulation of peroxisome proliferator-activated receptor-gamma2 activity as a central mechanism. Bone 2013, 56, 255–265. [Google Scholar] [CrossRef]
- Chalise, J.P.; Hashimoto, S.; Parajuli, G.; Kang, S.; Singh, S.K.; Gemechu, Y.; Metwally, H.; Nyati, K.K.; Dubey, P.K.; Zaman, M.M.U.; et al. Feedback regulation of Arid5a and Ppar-gamma2 maintains adipose tissue homeostasis. Proc. Natl. Acad. Sci. USA 2019, 116, 15128–15133. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; He, P.P.; Zhang, D.W.; Zheng, X.L.; Cayabyab, F.S.; Yin, W.D.; Tang, C.K. Lipoprotein lipase: From gene to atherosclerosis. Atherosclerosis 2014, 237, 597–608. [Google Scholar] [CrossRef]
- Voshol, P.J.; Rensen, P.C.; van Dijk, K.W.; Romijn, J.A.; Havekes, L.M. Effect of plasma triglyceride metabolism on lipid storage in adipose tissue: Studies using genetically engineered mouse models. Biochim. Biophys. Acta 2009, 1791, 479–485. [Google Scholar] [CrossRef]
- Gregoire, F.M. Adipocyte differentiation: From fibroblast to endocrine cell. Exp. Biol. Med. 2001, 226, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Filipovic, R.; Kumar, S.S.; Bahr, B.A.; Loturco, J. Slice Culture Method for Studying Migration of Neuronal Progenitor Cells Derived from Human Embryonic Stem Cells (hESC). Curr. Protoc. Stem Cell Biol 2014, 29, 1H.7.1–1H.7.14. [Google Scholar] [CrossRef]
- Ayanlaja, A.A.; Xiong, Y.; Gao, Y.; Ji, G.; Tang, C.; Abdikani Abdullah, Z.; Gao, D. Distinct Features of Doublecortin as a Marker of Neuronal Migration and Its Implications in Cancer Cell Mobility. Front. Mol. Neurosci. 2017, 10, 199. [Google Scholar] [CrossRef] [Green Version]
- Sainio, M.T.; Rasila, T.; Molchanova, S.M.; Järvilehto, J.; Torregrosa-Muñumer, R.; Harjuhaahto, S.; Pennonen, J.; Huber, N.; Herukka, S.K.; Haapasalo, A.; et al. Neurofilament Light Regulates Axon Caliber, Synaptic Activity, and Organelle Trafficking in Cultured Human Motor Neurons. Front. Cell Dev. Biol. 2021, 9, 820105. [Google Scholar] [CrossRef]
- Foudah, D.; Monfrini, M.; Donzelli, E.; Niada, S.; Brini, A.T.; Orciani, M.; Tredici, G.; Miloso, M. Expression of neural markers by undifferentiated mesenchymal-like stem cells from different sources. J. Immunol. Res. 2014, 2014, 987678. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, J.; Dhayal, M. Electrochemically differentiated human MSCs biosensing platform for quantification of nestin and beta-III tubulin as whole-cell system. Biosens. Bioelectron. 2022, 206, 114134. [Google Scholar] [CrossRef]
- Montzka, K.; Lassonczyk, N.; Tschöke, B.; Neuss, S.; Führmann, T.; Franzen, R.; Smeets, R.; Brook, G.A.; Wöltje, M. Neural differentiation potential of human bone marrow-derived mesenchymal stromal cells: Misleading marker gene expression. BMC Neurosci. 2009, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Liu, Y.; Wei, P.; Peng, H.; Winger, R.; Hussain, R.Z.; Ben, L.H.; Cravens, P.D.; Gocke, A.R.; Puttaparthi, K.; et al. Silencing Nogo-A promotes functional recovery in demyelinating disease. Ann. Neurol. 2010, 67, 498–507. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Genes | Sequence (5′ → 3′) |
|---|---|
| hGAPDH–F | AGC-CAC-ATC-GCT-CAG-ACA-C |
| hGAPDH–R | GCC-CAA-TAC-GAC-CAA-ATC-C |
| hCD90/THY1–F | AAT-ACC-AGC-AGT-TCA-CCC-AT |
| hCD90/THY1–R | GCT-AGT-GAA-GGC-GGA-TAA-GT |
| hCD105–F | AGT-CTT-GCA-GAA-ACA-GTC-CA |
| hCD105–R | TGG-ACT-TCA-AGG-ATG-GCA-TT |
| hRUNX2–F | CCG-CTT-CAG-TGA-TTT-AGG-GC |
| hRUNX2–R | GGG-TCT-GTA-ATC-TGA-CTC-TGT-CC |
| hALP–F | ATG-AAG-GAA-AAG-CCA-AGC-AG |
| hALP–R | ATG-GAG-ACA-TTC-TCT-CGT-TC |
| hOSX–F | ACG-GGT-CAG-GTA-GAG-TGA-GC |
| hOSX–R | GGG-ATC-CCC-CTA-ATC-AAG-AG |
| hLPL–F | ACG-GCA-TGT-GAA-TTC-TGT-GA |
| hLPL–R | GGA-TGT-GCT-ATT-TGG-CCA-CT |
| hPPARγ2–F | GAA-CGA-CCA-AGT-AAC-TCT-CC |
| hPPARγ2–R | CGC-AGG-CTC-TTT-AGA-AAC-TCC |
| hβIII-Tubulin–F | GGC-CAA-GGG-TCA-CTA-CAC-G |
| hβIII-Tubulin–R | GCA-GTC-GCA-GTT-TTC-ACA-CTC |
| hNeurofilament-L–F | ATG-AGT-TCC-TTC-AGC-TAC-GAG-C |
| hNeurofilament-L–R | CTG-GGC-ATC-AAC-GAT-CCA-GA |
| hDCX–F | TCC-CGG-ATC-AAT-GGG-TTG-C |
| hDCX–R | GCG-TAC-ACA-ATC-CCC-TTG-AAG-TA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, C.F.; Shen, J.; Balic, A.; Pagella, P.; Schwab, M.E.; Mitsiadis, T.A. Nogo-A Regulates the Fate of Human Dental Pulp Stem Cells toward Osteogenic, Adipogenic, and Neurogenic Differentiation. Cells 2022, 11, 3415. https://doi.org/10.3390/cells11213415
Lai CF, Shen J, Balic A, Pagella P, Schwab ME, Mitsiadis TA. Nogo-A Regulates the Fate of Human Dental Pulp Stem Cells toward Osteogenic, Adipogenic, and Neurogenic Differentiation. Cells. 2022; 11(21):3415. https://doi.org/10.3390/cells11213415
Chicago/Turabian StyleLai, Chai Foong, Juliet Shen, Anamaria Balic, Pierfrancesco Pagella, Martin E. Schwab, and Thimios A. Mitsiadis. 2022. "Nogo-A Regulates the Fate of Human Dental Pulp Stem Cells toward Osteogenic, Adipogenic, and Neurogenic Differentiation" Cells 11, no. 21: 3415. https://doi.org/10.3390/cells11213415