
DICAM in the Extracellular Vesicles from Astrocytes Attenuates Microglia Activation and Neuroinflammation

Abstract
:1. Background
2. Materials and Methods
2.1. Animal and Mouse Models of CRPS
2.2. Hind Paw Volume Measurement
2.3. Threshold Punctate Mechanical Stimulation (von Frey Test)
2.4. Spontaneous Weight-Bearing Test (Incapacitance Test)
2.5. Rotarod Test
2.6. Immunofluorescence Staining
2.7. Primary Cell Isolation and BV2 Microglia Cell Line Culture
2.8. Nitric Oxide (NO) Quantification
2.9. Real-Time Quantitative PCR and Reverse Transcription PCR
2.10. Enzyme-Linked Immunosorbent Assay (ELISA)
2.11. Western Blot Analysis
2.12. Blood Sampling from CRPS Patients and Controls
2.13. Isolation of Extracellular Vesicles
2.14. Astrocyte-Derived Conditioned Media Treatment
2.15. Statistical Analysis
3. Results
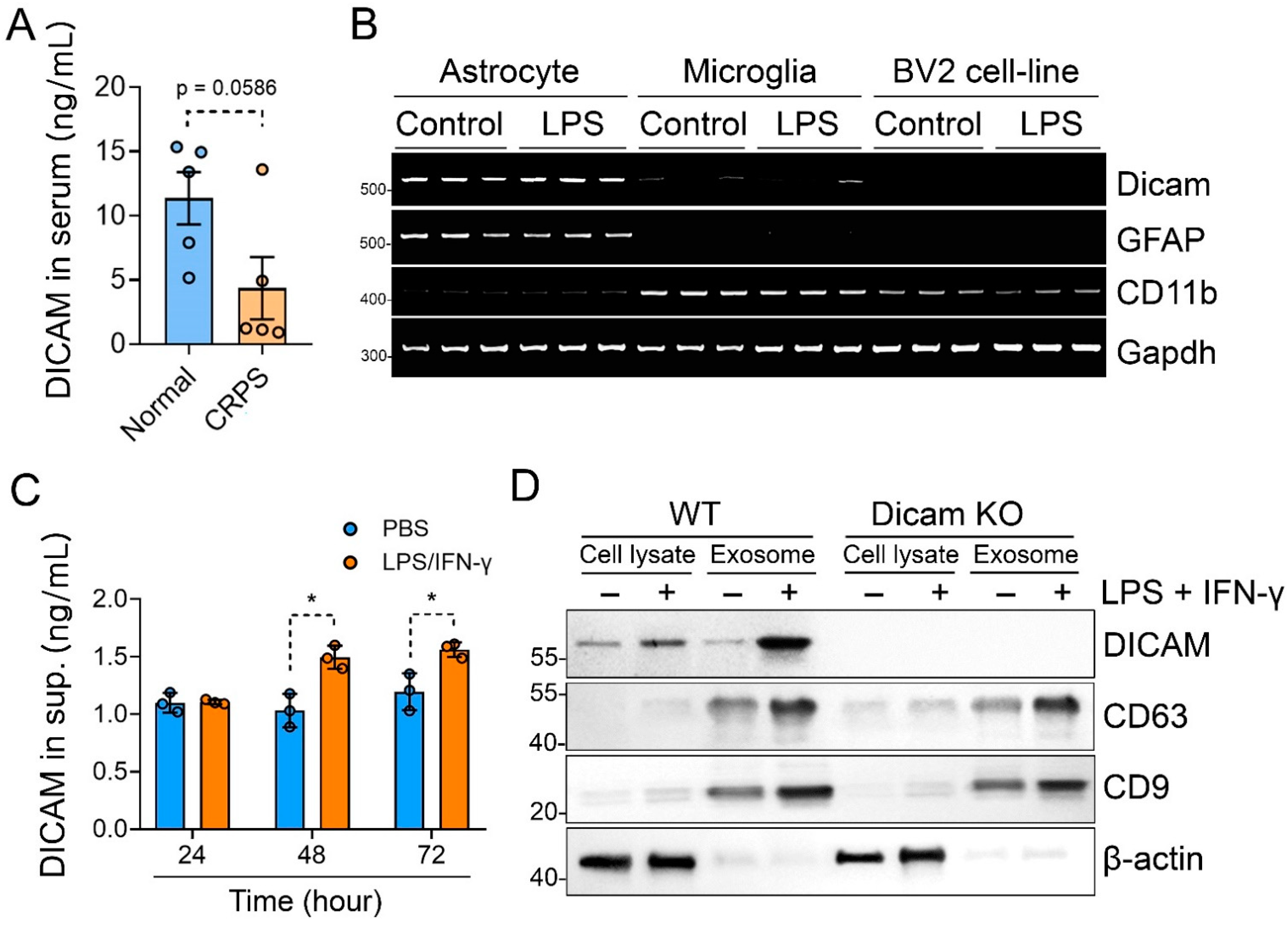
3.1. DICAM KO Mice Showed Enhanced Nociceptive Behavior and Increased Number of Astrocytes and Microglia in the Dorsal Horn of the CRPS Model
3.2. DICAM Deficiency Aggravates Proinflammatory Glial Cell Responses to Inflammatory Stimuli
3.3. DICAM Is Involved in the Activation of p38 MAPK in Mixed Glial Cells but Not in Astrocytes
3.4. Activated Astrocytes Secrete DICAM through Extracellular Vesicles
3.5. DICAM from Activated Astrocytes Acts as Coupling Factor That Regulates Mixed Glial Cell Activation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harte, S.E.; Harris, R.E.; Clauw, D.J. The neurobiology of central sensitization. J. Appl. Biobehav. Res. 2018, 23, e12137. [Google Scholar] [CrossRef]
- Nijs, J.; George, S.Z.; Clauw, D.J.; Fernández-de-las-Peñas, C.; Kosek, E.; Ickmans, K.; Fernández-Carnero, J.; Polli, A.; Kapreli, E.; Huysmans, E.; et al. Central sensitisation in chronic pain conditions: Latest discoveries and their potential for precision medicine. Lancet Rheumatol. 2021, 3, e383–e392. [Google Scholar] [CrossRef]
- Alban, L.; Clifford, J.W. Central Sensitization: A Generator of Pain Hypersensitivity by Central Neural Plasticity. J. Pain 2009, 10, 895–926. [Google Scholar]
- Ji, R.R.; Nackley, A.; Huh, Y.; Terrando, N.; Maixner, W. Neuroinflammation and Central Sensitization in Chronic and Widespread Pain. Anesthesiology 2018, 129, 343–366. [Google Scholar] [CrossRef]
- Lee, B.J.; Kim, J.Y.; Cho, H.J.; Park, D. Sphingosine 1-phosphate receptor modulation attenuate mechanical allodynia in mouse model of chronic complex regional pain syndrome by suppressing pathogenic astrocyte activation. Reg. Anesth. Pain. Med. 2020, 45, 230–238. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, J.S.; Park, D. Anti-allodynic effect of interleukin 10 in a mouse model of complex regional pain syndrome through reduction of NK1 receptor expression of microglia in the spinal cord. J. Pain Res. 2018, 11, 1729–1741. [Google Scholar] [CrossRef]
- Tay, T.L.; Savage, J.C.; Hui, C.W.; Bisht, K.; Tremblay, M.E. Microglia across the lifespan: From origin to function in brain development, plasticity and cognition. J. Physiol. 2017, 595, 1929–1945. [Google Scholar] [CrossRef]
- Ji, R.R.; Chamessian, A.; Zhang, Y.Q. Pain regulation by non-neuronal cells and inflammation. Science 2016, 354, 572–577. [Google Scholar] [CrossRef]
- Kawasaki, Y.; Zhang, L.; Cheng, J.K.; Ji, R.R. Cytokine mechanisms of central sensitization: Distinct and overlapping role of interleukin-1beta, interleukin-6, and tumor necrosis factor-alpha in regulating synaptic and neuronal activity in the superficial spinal cord. J. Neurosci. 2008, 28, 5189–5194. [Google Scholar] [CrossRef]
- Coull, J.A.M.; Beggs, S.; Boudreau, D.; Boivin, D.; Tsuda, M.; Inoue, K.; Gravel, C.; Salter, M.W.; De Koninck, Y. BDNF from microglia causes the shift in neuronal anion gradient underlying neuropathic pain. Nature 2005, 438, 1017–1021. [Google Scholar] [CrossRef]
- Clark, A.K.; Gruber-Schoffnegger, D.; Drdla-Schutting, R.; Gerhold, K.J.; Malcangio, M.; Sandkuhler, J. Selective activation of microglia facilitates synaptic strength. J. Neurosci. 2015, 35, 4552–4570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linnerbauer, M.; Wheeler, M.A.; Quintana, F.J. Astrocyte Crosstalk in CNS Inflammation. Neuron 2020, 108, 608–622. [Google Scholar] [CrossRef] [PubMed]
- Pekny, M.; Wilhelmsson, U.; Pekna, M. The dual role of astrocyte activation and reactive gliosis. Neurosci. Lett. 2014, 565, 30–38. [Google Scholar] [CrossRef]
- Han, R.T.; Kim, R.D.; Molofsky, A.V.; Liddelow, S.A. Astrocyte-immune cell interactions in physiology and pathology. Immunity 2021, 54, 211–224. [Google Scholar] [CrossRef]
- Upadhya, R.; Zingg, W.; Shetty, S.; Shetty, A.K. Astrocyte-derived extracellular vesicles: Neuroreparative properties and role in the pathogenesis of neurodegenerative disorders. J. Control. Release 2020, 323, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Neckles, V.N.; Morton, M.C.; Holmberg, J.C.; Sokolov, A.M.; Nottoli, T.; Liu, D.; Feliciano, D.M. A transgenic inducible GFP extracellular-vesicle reporter (TIGER) mouse illuminates neonatal cortical astrocytes as a source of immunomodulatory extracellular vesicles. Sci. Rep. 2019, 9, 3094. [Google Scholar] [CrossRef]
- Dickens, A.M.; Tovar-Y-Romo, L.B.; Yoo, S.-W.; Trout, A.L.; Bae, M.; Kanmogne, M.; Megra, B.; Williams, D.W.; Witwer, K.W.; Gacias, M.; et al. Astrocyte-shed extracellular vesicles regulate the peripheral leukocyte response to inflammatory brain lesions. Sci. Signal. 2017, 10, eaai7696. [Google Scholar] [CrossRef]
- Hu, G.; Liao, K.; Niu, F.; Yang, L.; Dallon, B.W.; Callen, S.; Tian, C.; Shu, J.; Cui, J.; Sun, Z.; et al. Astrocyte EV-Induced lincRNA-Cox2 Regulates Microglial Phagocytosis: Implications for Morphine-Mediated Neurodegeneration. Mol. Ther. Nucleic Acids 2018, 13, 450–463. [Google Scholar] [CrossRef]
- Long, X.; Yao, X.; Jiang, Q.; Yang, Y.; He, X.; Tian, W.; Zhao, K.; Zhang, H. Astrocyte-derived exosomes enriched with miR-873a-5p inhibit neuroinflammation via microglia phenotype modulation after traumatic brain injury. J. Neuroinflammation 2020, 17, 89. [Google Scholar] [CrossRef]
- Liao, K.; Niu, F.; Hu, G.; Yang, L.; Dallon, B.; Villarreal, D.; Buch, S. Morphine-mediated release of miR-138 in astrocyte-derived extracellular vesicles promotes microglial activation. J. Extracell. Vesicles 2020, 10, e12027. [Google Scholar] [CrossRef]
- Yonezawa, T.; Ohtsuka, A.; Yoshitaka, T.; Hirano, S.; Nomoto, H.; Yamamoto, K.; Ninomiya, Y. Limitrin, a novel immunoglobulin superfamily protein localized to glia limitans formed by astrocyte endfeet. Glia 2003, 44, 190–204. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.K.; Jin, J.S.; Jeong, J.H.; Kim, H.N.; Park, N.R.; Choi, J.Y. DICAM, a novel dual immunoglobulin domain containing cell adhesion molecule interacts with alphavbeta3 integrin. J. Cell. Physiol. 2008, 216, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.K.; Han, S.W.; Kim, G.W.; Jeong, J.H.; Kim, H.J.; Choi, J.Y. DICAM inhibits osteoclast differentiation through attenuation of the integrin alphaVbeta3 pathway. J. Bone Miner. Res. 2012, 27, 2024–2034. [Google Scholar] [CrossRef] [PubMed]
- Charabati, M.; Grasmuck, C.; Ghannam, S.; Bourbonniere, L.; Fournier, A.P.; Lecuyer, M.A.; Tastet, O.; Kebir, H.; Rebillard, R.M.; Hoornaert, C.; et al. DICAM promotes TH17 lymphocyte trafficking across the blood-brain barrier during autoimmune neuroinflammation. Sci. Transl. Med. 2022, 14, eabj0473. [Google Scholar] [CrossRef]
- Han, S.-W.; Jung, Y.-K.; Lee, E.-J.; Park, H.-R.; Kim, G.-W.; Jeong, J.-H.; Han, M.-S.; Choi, J.-Y. DICAM inhibits angiogenesis via suppression of AKT and p38 MAP kinase signalling. Cardiovasc. Res. 2013, 98, 73–82. [Google Scholar] [CrossRef]
- Han, S.-W.; Kim, J.M.; Lho, Y.; Cho, H.J.; Jung, Y.-K.; Kim, J.-A.; Lee, H.; Lee, Y.-J.; Kim, E.S. DICAM Attenuates Experimental Colitis via Stabilizing Junctional Complex in Mucosal Barrier. Inflamm. Bowel Dis. 2018, 25, 853–861. [Google Scholar] [CrossRef]
- Han, S.; Park, H.-R.; Lee, E.-J.; Jang, J.-A.; Han, M.-S.; Kim, G.-W.; Jeong, J.-H.; Choi, J.-Y.; Beier, F.; Jung, Y.-K. Dicam promotes proliferation and maturation of chondrocyte through Indian hedgehog signaling in primary cilia. Osteoarthr. Cartil. 2018, 26, 945–953. [Google Scholar] [CrossRef]
- Zhang, R.; Kim, A.S.; Fox, J.M.; Nair, S.; Basore, K.; Klimstra, W.B.; Diamond, M.S. Mxra8 is a receptor for multiple arthritogenic alphaviruses. Nature 2018, 557, 570–574. [Google Scholar] [CrossRef]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Shiotsuki, H.; Yoshimi, K.; Shimo, Y.; Funayama, M.; Takamatsu, Y.; Ikeda, K.; Takahashi, R.; Kitazawa, S.; Hattori, N. A rotarod test for evaluation of motor skill learning. J. Neurosci. Methods 2010, 189, 180–185. [Google Scholar] [CrossRef]
- Matejuk, A.; Ransohoff, R.M. Crosstalk Between Astrocytes and Microglia: An Overview. Front. Immunol. 2020, 11, 1416. [Google Scholar] [CrossRef]
- Chang, R.C.C.; Chen, W.; Hudson, P.; Wilson, B.; Han, D.S.K.; Hong, J.-S. Neurons reduce glial responses to lipopolysaccharide (LPS) and prevent injury of microglial cells from over-activation by LPS. J. Neurochem. 2001, 76, 1042–1049. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Sheng, S.; Wang, Y.; Ding, L.; Xu, X.; Xia, X.; Zheng, J.C. Astrocyte-derived extracellular vesicles: A double-edged sword in central nervous system disorders. Neurosci. Biobehav. Rev. 2021, 125, 148–159. [Google Scholar] [CrossRef]
- You, Y.; Borgmann, K.; Edara, V.V.; Stacy, S.; Ghorpade, A.; Ikezu, T. Activated human astrocyte-derived extracellular vesicles modulate neuronal uptake, differentiation and firing. J. Extracell. Vesicles 2020, 9, 1706801. [Google Scholar] [CrossRef]
- Altei, W.F.; Pachane, B.C.; Dos Santos, P.K.; Ribeiro, L.N.; Sung, B.H.; Weaver, A.M.; Selistre-de-Araújo, H.S. Inhibition of alphavbeta3 integrin impairs adhesion and uptake of tumor-derived small extracellular vesicles. Cell Commun. Signal. 2020, 18, 158. [Google Scholar] [CrossRef]
- Bachstetter, A.D.; Rowe, R.K.; Kaneko, M.; Goulding, D.; Lifshitz, J.; Van Eldik, L.J. The p38alpha MAPK regulates microglial responsiveness to diffuse traumatic brain injury. J. Neurosci. 2013, 33, 6143–6153. [Google Scholar] [CrossRef]
- Schafers, M.; Svensson, C.I.; Sommer, C.; Sorkin, L.S. Tumor necrosis factor-alpha induces mechanical allodynia after spinal nerve ligation by activation of p38 MAPK in primary sensory neurons. J. Neurosci. 2003, 23, 2517–2521. [Google Scholar] [CrossRef]
- Perea, J.R.; García, E.; Vallés-Saiz, L.; Cuadros, R.; Hernández, F.; Bolós, M.; Avila, J. p38 activation occurs mainly in microglia in the P301S Tauopathy mouse model. Sci. Rep. 2022, 12, 2130. [Google Scholar] [CrossRef]
- Bachstetter, A.D.; Xing, B.; de Almeida, L.; Dimayuga, E.R.; Watterson, D.M.; Van Eldik, L.J. Microglial p38alpha MAPK is a key regulator of proinflammatory cytokine up-regulation induced by toll-like receptor (TLR) ligands or beta-amyloid (Abeta). J Neuroinflamm. 2011, 8, 79. [Google Scholar] [CrossRef]
- Jin, M.-M.; Wang, F.; Qi, D.; Liu, W.-W.; Gu, C.; Mao, C.-J.; Yang, Y.-P.; Zhao, Z.; Hu, L.-F.; Liu, C.-F. A Critical Role of Autophagy in Regulating Microglia Polarization in Neurodegeneration. Front. Aging Neurosci. 2018, 10, 378. [Google Scholar] [CrossRef]
- He, Y.; She, H.; Zhang, T.; Xu, H.; Cheng, L.; Yepes, M.; Zhao, Y.; Mao, Z. p38 MAPK inhibits autophagy and promotes microglial inflammatory responses by phosphorylating ULK1. J. Cell. Biol. 2018, 217, 315–328. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Chen, J.; Cheng, Y.; Fu, Y.; Zhao, H.; Tang, M.; Tan, J. Mesenchymal stem cell-derived exosomes protect beta cells against hypoxia-induced apoptosis via miR-21 by alleviating ER stress and inhibiting p38 MAPK phosphorylation. Stem. Cell. Res. Ther. 2020, 11, 97. [Google Scholar] [CrossRef]
- Zhang, Z.; Luo, X.; Ding, S.; Chen, J.; Chen, T.; Chen, X.; Zha, H.; Yao, L.; He, X.; Peng, H. MicroRNA-451 regulates p38 MAPK signaling by targeting of Ywhaz and suppresses the mesangial hypertrophy in early diabetic nephropathy. FEBS Lett. 2012, 586, 20–26. [Google Scholar] [CrossRef]
- Ji, R.R.; Berta, T.; Nedergaard, M. Glia and pain: Is chronic pain a gliopathy? Pain 2013, 154 (Suppl. 1), S10–S28. [Google Scholar] [CrossRef]
- Garrison, C.J.; Dougherty, P.M.; Kajander, K.C.; Carlton, S.M. Staining of glial fibrillary acidic protein (GFAP) in lumbar spinal cord increases following a sciatic nerve constriction injury. Brain. Res. 1991, 565, 1–7. [Google Scholar] [CrossRef]
- Ikeda, H.; Kiritoshi, T.; Murase, K. Contribution of microglia and astrocytes to the central sensitization, inflammatory and neuropathic pain in the juvenile rat. Mol. Pain 2012, 8, 43. [Google Scholar] [CrossRef]
- Schinkel, C.; Gaertner, A.; Zaspel, J.; Zedler, S.; Faist, E.; Schuermann, M. Inflammatory mediators are altered in the acute phase of posttraumatic complex regional pain syndrome. Clin. J. Pain 2006, 22, 235–239. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Primer Sequences |
|---|---|---|
| Il-1β | Forward | TGA AAT GCC ACC TTT TGA CAG TG |
| Reverse | ATG TGC TGC TGC GAG ATT TG | |
| Cxcl10 | Forward | CCA AGT GCT GCC GTC ATT TTC |
| Reverse | TCC CTA AGG CCC TCA TTC TCA | |
| Il-10 | Forward | CTT ACT GAC TGG CAT GAG GAT CA |
| Reverse | GCA GCT CTA GGA GCA TGT GG | |
| Cd86 | Forward | TCA ATG GGA CTG CAT ATC TGC C |
| Reverse | GCC AAA ATA CTA CCA GCT CAC T | |
| Gfap | Forward | AGG CAG AAG CTC CAA GAT GA |
| Reverse | TGT GAG GTC TGC AAA CTT GG | |
| Cd68 | Forward | TGT CTG ATC TTG CTA GGA CCG |
| Reverse | GAG AGT AAC GGC CTT TTT GTG A | |
| Arg | Forward | ACA TCA ACA CTC CCC TGA CAA |
| Reverse | TAC GTC TCG CAA GCC AAT GTA | |
| Cd11b | Forward | CTG GTG CTC TTG GCT CTC AT |
| Reverse | GGC AGC TTC ATT CAT CAT GT | |
| Gapdh | Forward | AGC CCA AGA TGC CCT TCA GT |
| Reverse | CCG TGT TCC TAC CCC CAA TG |
| Age | Sex | Onset Duration | NRS | Pain Site | |
|---|---|---|---|---|---|
| 1 | 81 | Male | 32 | 8 | Lt arm |
| 2 | 55 | Male | 40 | 7 | Rt arm |
| 3 | 65 | Male | 39 | 9 | Lt arm |
| 4 | 54 | Male | 32 | 6 | Lt arm |
| 5 | 48 | Female | 32 | 7 | Lt leg |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, J.; Cho, H.-J.; Park, D.; Han, S. DICAM in the Extracellular Vesicles from Astrocytes Attenuates Microglia Activation and Neuroinflammation. Cells 2022, 11, 2977. https://doi.org/10.3390/cells11192977
Han J, Cho H-J, Park D, Han S. DICAM in the Extracellular Vesicles from Astrocytes Attenuates Microglia Activation and Neuroinflammation. Cells. 2022; 11(19):2977. https://doi.org/10.3390/cells11192977
Chicago/Turabian StyleHan, Jin, Hyun-Jung Cho, Donghwi Park, and Seungwoo Han. 2022. "DICAM in the Extracellular Vesicles from Astrocytes Attenuates Microglia Activation and Neuroinflammation" Cells 11, no. 19: 2977. https://doi.org/10.3390/cells11192977