
Emerging Roles of miRNA, lncRNA, circRNA, and Their Cross-Talk in Pituitary Adenoma

Abstract
:1. Introduction
2. miRNA and PA

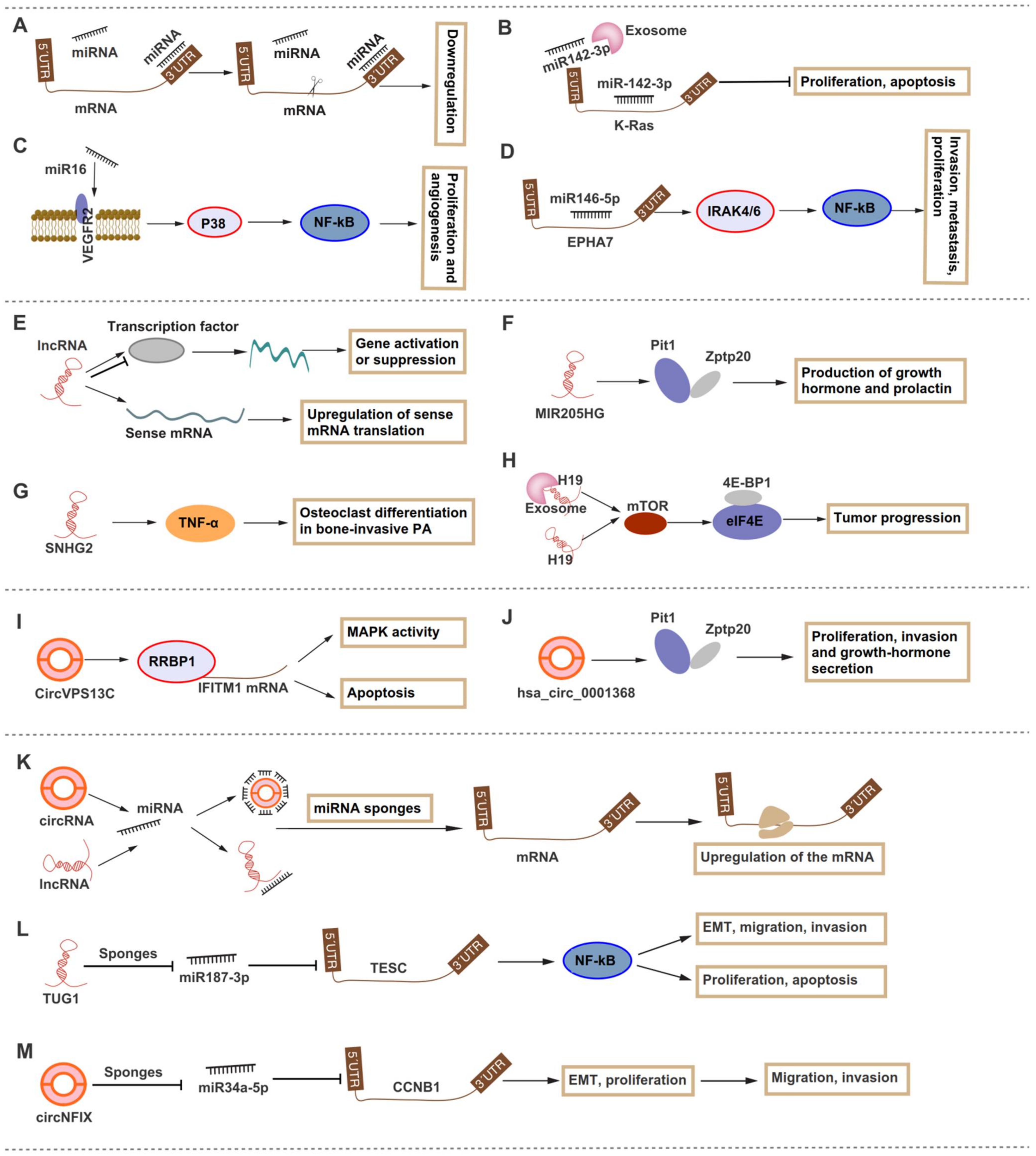{kind=link}
| Author (Ref.) | Type of PA | Sample | miRNA | Expression | Target Gene | Biological Function |
|---|---|---|---|---|---|---|
| Mossakowska [17] | CD | Tissues, AtT-20/D16v-F2 cells | hsa-miR-124-3p and hsa-miR-135-5p | Down | NR3C1, NR3C2 | Regulation of glucocorticoid receptor |
| Zhang [18] | NFPA | Serum | miRNA-26b, miRNA-138, miRNA-206, and let-7e | Down | NA | Tumor-suppressor gene |
| Chen [19] | PA | Tissues, GH3 cells | miR-424-3p | Down | JAG1 | Cell proliferation and EMT |
| Zhao [20] | IPA | Exosomes, tissues, and GH4C1, MMP, and GH3 cells | miR-149-5p and miR-99a-3p | Down | NOVA1, DTL, RAB27B | The expression of EMT- and ECM-related markers and tumor-related genes |
| Wei [21] | PA | Tissues, GH3 cells | miR-137 | Down | EGFR | Tumor suppressor, cell proliferation |
| Wu [22] | IPA | Tissues | miR-193a-3p | Down | Higher risk of postoperative residual and recurrence | |
| Wang [23] | PA | GT1.1 and AtT-20 cells | miR-219a-2-3p | Down | MDM2 | Cell proliferation and apoptosis |
| Németh [24] | GHPA, FSH/LH+ PA, hormone-immunonegative PA | Plasma | miR-143-3p | Down | NA | Distinguishing preoperative plasma samples from normal controls |
| He [25] | NFPA, GHPA, PRLPA | Tissues | miR-34c-3p, miR-34b-5p, miR-378, miR-338-5p, miR-493-5p, miR-181b-5p, miR-184, and miR-124-3p | PRLPA: miR-34c-3p, miR-34b-5p, miR-378, and miR-338-5p were downregulated; NFPA: miR-493-5p and miR-124-3p were downregulated and miR-181b-5p was upregulated; GHPA: miR-184 was upregulated and miR-124-3p was downregulated | NA | Novel biomarkers |
| Cai [26] | NFPA | Tissues | miR-370 | Down | HMGA2 | miR-370 was decreased by CXCL12 treatment and miR-370 inhibited tumor growth and invasiveness |
| Song [27] | NFPA | Tissues, GH3 cells | miR-137 | Down | WIF1 | Cell proliferation and invasion |
| Leone [28] | GHPA, NFPA | miR-23b and miR-130b | Down | HMGA2, CCNA2 | Cell proliferation arresting the cells in the G1 and G2 phase of the cell cycle | |
| Gentilin [29] | ACTH-secreting PA | Tissues, AtT-20 cells | miR-26a | Up | PRKCD | Delayed the cell cycle in G1 phase |
| Trivellin [30] | GHPA, NFPA | Tissues, GH3 cells | miR-107 | Up | AIP | Cell proliferation |
| Butz [31] | NFPA | Tissues | miR-135a, miR-140-5p, miR-582-3p, miR-582-5p, and miR-938 | Up | Smad3 | Tumor size |
| Butz [32] | NFPA, GHPA | Tissues | miR-128a, miR-155, and miR-516a-3p | Up | Wee1 | Cell proliferation |
| Qian [33] | PA | Tissues | let-7 | Down | HMGA2 | Tumor invasion |
| Bottoni [34] | GHPA, PRLPA | Tissues | miR-15a and miR-16-1 | Down | Tumor diameter | |
| Lyu [35] | NFPA | Exosome | hsa-miR-486-5p | Down | NA | Tumor progression |
| He [36] | PA | Tissues, MMQ and HP75 cells | miR-448 | Down | BCL2 | Cell proliferation and migration |
| Xiong [37] | GHPA | Exosome, tissues | hsa-miR-21-5p | Up | PDCD4 | Bone formation and trabecula number |
| Yang [38] | PA | GH4C1 cells | miR-34a | Down | Sox7 | Cell proliferation and apoptosis |
| Fan [39] | GHPA | Tissues, GH3 cells | miR-185 | Up | SSTR2 | Cell proliferation and apoptosis |
| Liao [40] | PA | GH3 and MMQ cells | miR-200c | Up | PTEN | Pituitary tumor formation |
2.1. Circulating miRNA and PA
2.2. miRNA in PA Tissue
2.3. miRNA in PA-Associated Cell Lines
2.4. Exosome-Derived miRNA and PA
3. lncRNA and PA
| Author (Ref.) | Type of PA | Sample | lncRNA | Expression | Target Gene | Biological Function |
|---|---|---|---|---|---|---|
| Peng [85] | IPA | Tissues | FAM182B, LOC105371531, LOC105375785 | Down | NA | FAM182B and LOC105375785 can distinguish IPA from NIPA. |
| Cheng [94] | NFPA | Tissues | LOC101927765, RP11-23N2.4, RP4-533D7.4 | NA | NA | High prediction accuracy for NFPA recurrence. |
| Lu [95] | PA | Tissues, HP75 cell lines | IFNG-AS1 | Up | ESRP2 | An oncogene that promoted tumor progression. |
| Lu [90] | GHPA | Tissues | H19, MALAT-1 | Up | NA | NA |
| Fu [96] | PA | Tissues, HP75 cells | CCTA2 | Up | PTTG1 | Poor prognosis, cell proliferation, migration, and invasion. |
| Li [97] | NFPA | Tissues | MEG, HOTAIR | MEG3: Down; HOTAIR: Up | PCNA | Tumor development and invasion. |
| Cheng [98] | NFPA | Tissues | COA6-AS1, RP11-23N2.4 | Up | NA | Tumor regrowth with a high predictive accuracy. |
| Zhang [99] | PA | Tissues, GH3 and HP75 cells | PVT1 | Up | NA | Cell migration, proliferation, and EMT. |
| Zhang [93] | GHPA, NFPA, PRLPA | Exosome, GH3 cells | H19 | Down | 4E-BP1 | The prognosis or drug response. |
| Xing [84] | NFPA | Tissues | n334366, n335657, n409198, MEG3, n337303, n340496, n334406, n332607, n333074, n332409 | Down: n334366, n335657, n409198, MEG3. Up: n337303, n340496, n334406, n332607, n333074, n332409 | NA | Tumorigenesis. |
4. circRNA and PA
| Author (Ref.) | Type of PA | Sample | circRNA | Expression | Target Gene | Biological Function |
|---|---|---|---|---|---|---|
| Hu [111] | NFPA | Tissues | hsa_circRNA_102597 | Down | NA | Associated with tumor diameter and Knosp grade, differentiated invasive from noninvasive NFPAs, and predicted tumor progression and recurrence. |
| Zhang [101] | NFPA | Tissues, PDFS cells | CircVPS13C | Up | RRBP1 | Promoted proliferation and development. |
| Du [110] | GHPA | Tissues | hsa_circ_0001368 | Up | Pit-1 | Associated with the invasiveness and serum GH level; promoted cell proliferation and invasion. |
| Guo [109] | NFPA | Tissues | hsa_circ_0000066 and hsa_circ_0069707 | Up | NA | Associated with the PFS, and had a high prediction accuracy for tumor recurrence. |
| Wang [112] | NFPA | Tissues | hsa_circ_0054722, hsa_circ_0007362, hsa_circ_0012346, hsa_circ_0062222, hsa_circ_0016403, hsa_circ_0033349 | Up: hsa_circ_0054722, hsa_circ_0007362, hsa_circ_0012346, Down: hsa_circ_0062222, hsa_circ_0016403, hsa_circ_0033349 | NA | Contributed to diagnosis, prognosis, and clinical treatment. |
5. ceRNA and PA
| Author (Ref.) | Type of PA | Sample | LncRNA/ circRNA | Expression | Target miRNA | miRNA Target | Biological Function |
|---|---|---|---|---|---|---|---|
| Du [123] | NFPA | Tissues, pituitary tumor-derived folliculostellate (PDFS) cell line | circOMA1 | Down | miR-145-5p | TPT1 | Promoted cell proliferation and invasiveness. |
| Cheng [124] | IPA | Tissues; GT1-1 and GH3 cells | circNFIX | Up | miR-34a-5p | CCNB1 | Promoted cell invasion, migration, and proliferation. |
| Zhu [79] | BIPA | Tissues | MEG8 | Up | miR-454-3p | TNF-α | Promoted bone destruction and associated with poor PFS. |
| Zhao [125] | IPA | Tissues; RC-4B/C and GH3 cells | PCAT6 | Up | miR-139-3p | BRD4 | Regulated the progression of PA. |
| Wu [78] | PRLPA | Tissues, GH3 cells | H19 | Down | miR-93a | ATG7 | H19 had a synergistic effect with dopamine agonist treatment on prolactinomas. |
| Yue [126] | PA | Tissues; GH1, RC-4B/C, GH3 and MMQ cell lines | SNHG7 | Up | miR-449a | NA | Associated with poor survival outcomes; increased cell viability, migration, and invasion and decreased apoptosis. |
| Zhu [86] | BIPA | Tissues; GH3 and RAW264.7 cells | NR_033258, SNHG24 | Up | miR-181c-5p, miR-454-3p | NA | TNFα induced osteoclast differentiation, and NR_033258 and SNHG24 regulated TNFα expression. |
| Qiu [119] | PA | HP75 cells | LINC01004 | Up | miR-323a-3p/miR-136-5p | RCN2 | Promoted disease progression. |
| Li [120] | IPA | GH3 and HP75 cells | KCNQ1OT1 | Up | miR-140-5p | RAB11A | Promoted the EMT, cellular stemness, and proliferation and invasion. |
| Wu [115] | PA | Tissues; RC-4B/C, GH3, and MMQ cells | BBOX1-AS1 | Up | miR-361-3p | E2F1 | Promoted cell invasion, apoptosis, and proliferation. |
| Huang [116] | PA | HP75 cells | LINC01116 | Up | miR-744-5p | HOXB8 | Promoted cell proliferation, migration, and EMT process. |
| Li [121] | IPA | Tissues; AtT-20 and GT1-1 cells | LINC00473 | Up | miR-502-3p | KMT5A | Promoted cell proliferation. |
| Zhang [114] | PA | Tissues; HP75 and GH3 cells | TUG1 | Up | miR-187-3p | TESC | Promoted cell proliferation, invasion, migration, and EMT, and inhibited apoptosis. |
| Mao [122] | IPA | Tissues, HP75 cells | SNHG6 | Up | miR-944 | RAB11A | Promoted cell viability, migration, invasion, and EMT. |
| Wang [117] | PA | Tissues; GH3 and MMQ cells | MEG3 | Down | miR-23b-3p | FOXO4 | Promoted cell proliferation, apoptosis, and the EMT process. |
| Zhu [118] | NFPA | Tissues, PDFS cells | MEG3 | Down | miR-376B-3p | HMGA2 | Tumorigenesis and cell apoptosis |
6. Conclusions and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Beylerli, O.; Beeraka, N.M.; Gareev, I.; Pavlov, V.; Yang, G.; Liang, Y.; Aliev, G. MiRNAs as Noninvasive Biomarkers and Therapeutic Agents of Pituitary Adenomas. Int. J. Mol. Sci. 2020, 21, 7287. [Google Scholar] [CrossRef] [PubMed]
- Campana, C.; Nista, F.; Castelletti, L.; Caputo, M.; Lavezzi, E.; Marzullo, P.; Ferrero, A.; Gaggero, G.; Canevari, F.R.; Rossi, D.C.; et al. Clinical and radiological presentation of parasellar ectopic pituitary adenomas: Case series and systematic review of the literature. J. Endocrinol. Investig. 2022, 45, 1465–1481. [Google Scholar] [CrossRef] [PubMed]
- Giuffrida, G.; D’Argenio, V.; Ferraù, F.; Lasorsa, V.A.; Polito, F.; Aliquò, F.; Ragonese, M.; Cotta, O.R.; Alessi, Y.; Oteri, R.; et al. Methylome Analysis in Nonfunctioning and GH-Secreting Pituitary Adenomas. Front. Endocrinol. 2022, 13, 841118. [Google Scholar] [CrossRef] [PubMed]
- Trouillas, J.; Jaffrain-Rea, M.L.; Vasiljevic, A.; Raverot, G.; Roncaroli, F.; Villa, C. How to Classify the Pituitary Neuroendocrine Tumors (PitNET)s in 2020. Cancers 2020, 12, 514. [Google Scholar] [CrossRef]
- Spada, A.; Mantovani, G.; Lania, A.G.; Treppiedi, D.; Mangili, F.; Catalano, R.; Carosi, G.; Sala, E.; Peverelli, E. Pituitary Tumors: Genetic and Molecular Factors Underlying Pathogenesis and Clinical Behavior. Neuroendocrinology 2022, 112, 15–33. [Google Scholar] [CrossRef]
- Chang, M.; Yang, C.; Bao, X.; Wang, R. Genetic and Epigenetic Causes of Pituitary Adenomas. Front. Endocrinol. 2020, 11, 596554. [Google Scholar] [CrossRef]
- Xu, D.; Wang, L. The Involvement of miRNAs in Pituitary Adenomas Pathogenesis and the Clinical Implications. Eur. Neurol. 2022, 85, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Liu, P.; Fan, J.; Luo, Y. Long non-coding RNA H19: Physiological functions and involvements in central nervous system disorders. Neurochem. Int. 2021, 148, 105072. [Google Scholar] [CrossRef]
- DiStefano, J.K.; Gerhard, G.S. Long Noncoding RNAs and Human Liver Disease. Annu. Rev. Pathol. 2022, 17, 1–21. [Google Scholar] [CrossRef]
- An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [CrossRef] [Green Version]
- Kapranov, P.; Cheng, J.; Dike, S.; Nix, D.A.; Duttagupta, R.; Willingham, A.T.; Stadler, P.F.; Hertel, J.; Hackermüller, J.; Hofacker, I.L.; et al. RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science 2007, 316, 1484–1488. [Google Scholar] [CrossRef] [PubMed]
- Wery, M.; Kwapisz, M.; Morillon, A. Noncoding RNAs in gene regulation. Wiley Interdiscip. Rev. Syst. Biol. Med. 2011, 3, 728–738. [Google Scholar] [CrossRef] [PubMed]
- Bahreini, F.; Jabbari, P.; Gossing, W.; Aziziyan, F.; Frohme, M.; Rezaei, N. The role of noncoding RNAs in pituitary adenoma. Epigenomics 2021, 13, 1421–1437. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Huang, W.; Wang, X.; Zhang, J.; Tao, T.; Zheng, Y.; Liu, S.; Yang, J.; Chen, Z.S.; Cai, C.Y.; et al. Hsa-miR-3178/RhoB/PI3K/Akt, a novel signaling pathway regulates ABC transporters to reverse gemcitabine resistance in pancreatic cancer. Mol. Cancer 2022, 21, 112. [Google Scholar] [CrossRef]
- Zhang, L.X.; Gao, J.; Long, X.; Zhang, P.F.; Yang, X.; Zhu, S.Q.; Pei, X.; Qiu, B.Q.; Chen, S.W.; Lu, F.; et al. The circular RNA circHMGB2 drives immunosuppression and anti-PD-1 resistance in lung adenocarcinomas and squamous cell carcinomas via the miR-181a-5p/CARM1 axis. Mol. Cancer 2022, 21, 110. [Google Scholar] [CrossRef]
- Slack, F.J.; Chinnaiyan, A.M. The Role of Non-coding RNAs in Oncology. Cell 2019, 179, 1033–1055. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef]
- Meng, F.; Yu, W.; Chen, C.; Guo, S.; Tian, X.; Miao, Y.; Ma, L.; Zhang, X.; Yu, Y.; Huang, L.; et al. A Versatile Electrochemical Biosensor for the Detection of Circulating MicroRNA toward Non-Small Cell Lung Cancer Diagnosis. Small 2022, 18, e2200784. [Google Scholar] [CrossRef]
- Donati, S.; Aurilia, C.; Palmini, G.; Miglietta, F.; Falsetti, I.; Iantomasi, T.; Brandi, M.L. MicroRNAs as Potential Biomarkers in Pituitary Adenomas. Noncoding RNA 2021, 7, 55. [Google Scholar] [CrossRef]
- Tabet, F.; Vickers, K.C.; Cuesta Torres, L.F.; Wiese, C.B.; Shoucri, B.M.; Lambert, G.; Catherinet, C.; Prado-Lourenco, L.; Levin, M.G.; Thacker, S.; et al. HDL-transferred microRNA-223 regulates ICAM-1 expression in endothelial cells. Nat. Commun. 2014, 5, 3292. [Google Scholar] [CrossRef] [Green Version]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Németh, K.; Darvasi, O.; Likó, I.; Szücs, N.; Czirják, S.; Reiniger, L.; Szabó, B.; Krokker, L.; Pállinger, É.; Igaz, P.; et al. Comprehensive analysis of circulating microRNAs in plasma of patients with pituitary adenomas. J. Clin. Endocrinol. Metab. 2019, 104, 4151–4168. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ma, D.; Liu, H.; Wang, J.; Fan, J.; Li, X. MicroRNA-143 shows tumor suppressive effects through inhibition of oncogenic K-Ras in pituitary tumor. Int. J. Clin. Exp. Pathol. 2017, 10, 10969–10978. [Google Scholar]
- Amaral, F.C.; Torres, N.; Saggioro, F.; Neder, L.; Machado, H.R.; Silva, W.A., Jr.; Moreira, A.C.; Castro, M. MicroRNAs differentially expressed in ACTH-secreting pituitary tumors. J. Clin. Endocrinol. Metab. 2009, 94, 320–323. [Google Scholar] [CrossRef]
- Belaya, Z.; Khandaeva, P.; Nonn, L.; Nikitin, A.; Solodovnikov, A.; Sitkin, I.; Grigoriev, A.; Pikunov, M.; Lapshina, A.; Rozhinskaya, L.; et al. Circulating Plasma microRNA to Differentiate Cushing’s Disease From Ectopic ACTH Syndrome. Front. Endocrinol. 2020, 11, 331. [Google Scholar] [CrossRef]
- Lu, B.; Liu, G.L.; Yu, F.; Li, W.J.; Xiang, X.X.; Xiao, H.Z. MicroRNA-16/VEGFR2/p38/NF-κB signaling pathway regulates cell growth of human pituitary neoplasms. Oncol. Rep. 2018, 39, 1235–1244. [Google Scholar] [CrossRef]
- Di Ieva, A.; Butz, H.; Niamah, M.; Rotondo, F.; De Rosa, S.; Sav, A.; Yousef, G.M.; Kovacs, K.; Cusimano, M.D. MicroRNAs as biomarkers in pituitary tumors. Neurosurgery 2014, 75, 181–189, discussion 188–189. [Google Scholar] [CrossRef]
- Bottoni, A.; Piccin, D.; Tagliati, F.; Luchin, A.; Zatelli, M.C.; degli Uberti, E.C. miR-15a and miR-16-1 down-regulation in pituitary adenomas. J. Cell Physiol. 2005, 204, 280–285. [Google Scholar] [CrossRef]
- He, Z.; Chen, L.; Hu, X.; Tang, J.; He, L.; Hu, J.; Fei, F.; Wang, Q. Next-generation sequencing of microRNAs reveals a unique expression pattern in different types of pituitary adenomas. Endocr. J. 2019, 66, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gao, L.; Guo, X.; Feng, C.; Deng, K.; Lian, W.; Xing, B. Identification of microRNAs associated with the aggressiveness of prolactin pituitary tumors using bioinformatic analysis. Oncol. Rep. 2019, 42, 533–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Gao, L.; Guo, X.; Feng, C.; Deng, K.; Lian, W.; Xing, B. [Corrigendum] Identification of microRNAs associated with the aggressiveness of prolactin pituitary tumors using bioinformatic analysis. Oncol. Rep. 2021, 46, 130. [Google Scholar] [CrossRef] [PubMed]
- Vicchio, T.M.; Aliquò, F.; Ruggeri, R.M.; Ragonese, M.; Giuffrida, G.; Cotta, O.R.; Spagnolo, F.; Torre, M.L.; Alibrandi, A.; Asmundo, A.; et al. MicroRNAs expression in pituitary tumors: Differences related to functional status, pathological features, and clinical behavior. J. Endocrinol. Investig. 2020, 43, 947–958. [Google Scholar] [CrossRef]
- Lou, X.; Cai, Y.; Zheng, H.; Zhang, Y. MicroRNA-146b-5p/EPHA7 axis regulates cell invasion, metastasis, proliferation, and temozolomide-induced chemoresistance via regulation of IRAK4/TRAF6/NF-κB signaling pathway in aggressive pituitary adenoma. Histol. Histopathol. 2022, 37, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, J.; Zhang, C.; Wang, P.; Huang, C.; Peng, H. MiR-219a-2-3p suppresses cell proliferation and promotes apoptosis by targeting MDM2/p53 in pituitary adenomas cells. Biosci. Biotechnol. Biochem. 2020, 84, 911–918. [Google Scholar] [CrossRef] [PubMed]
- Russo, F.; Di Bella, S.; Vannini, F.; Berti, G.; Scoyni, F.; Cook, H.V.; Santos, A.; Nigita, G.; Bonnici, V.; Laganà, A.; et al. miRandola 2017: A curated knowledge base of non-invasive biomarkers. Nucleic. Acids Res. 2018, 46, D354-d359. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Cheng, J.; Li, B.; Nie, D.; Li, C.; Gui, S.; Wang, H.; Zhang, Y. Up-regulation of the expressions of MiR-149-5p and MiR-99a-3p in exosome inhibits the progress of pituitary adenomas. Cell Biol. Toxicol. 2021, 37, 633–651. [Google Scholar] [CrossRef]
- Katznelson, L.; Laws, E.R., Jr.; Melmed, S.; Molitch, M.E.; Murad, M.H.; Utz, A.; Wass, J.A. Acromegaly: An endocrine society clinical practice guideline. J. Clin. Endocrinol. Metab. 2014, 99, 3933–3951. [Google Scholar] [CrossRef]
- Cansu, G.B.; Yılmaz, N.; Yanıkoğlu, A.; Özdem, S.; Yıldırım, A.B.; Süleymanlar, G.; Altunbaş, H.A. Assessment of Diastolic Dysfunction, Arterial Stiffness, and Carotid Intima-Media Thickness in Patients with Acromegaly. Endocr. Pract. 2017, 23, 536–545. [Google Scholar] [CrossRef]
- Wassenaar, M.J.; Biermasz, N.R.; Bijsterbosch, J.; Pereira, A.M.; Meulenbelt, I.; Smit, J.W.; Roelfsema, F.; Kroon, H.M.; Romijn, J.A.; Kloppenburg, M. Arthropathy in long-term cured acromegaly is characterised by osteophytes without joint space narrowing: A comparison with generalised osteoarthritis. Ann. Rheum. Dis. 2011, 70, 320–325. [Google Scholar] [CrossRef]
- Xiong, Y.; Tang, Y.; Fan, F.; Zeng, Y.; Li, C.; Zhou, G.; Hu, Z.; Zhang, L.; Liu, Z. Exosomal hsa-miR-21-5p derived from growth hormone-secreting pituitary adenoma promotes abnormal bone formation in acromegaly. Transl. Res. 2020, 215, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Li, J.; Feng, J.; Li, Z.; Liu, Q.; Lv, P.; Wang, F.; Gao, H.; Zhang, Y. Identification of Serum miRNA-423-5p Expression Signature in Somatotroph Adenomas. Int. J. Endocrinol. 2019, 2019, 8516858. [Google Scholar] [CrossRef] [PubMed]
- Mossakowska, B.J.; Kober, P.; Rusetska, N.; Boresowicz, J.; Maksymowicz, M.; Pękul, M.; Zieliński, G.; Styk, A.; Kunicki, J.; Mandat, T.; et al. Difference in miRNA Expression in Functioning and Silent Corticotroph Pituitary Adenomas Indicates the Role of miRNA in the Regulation of Corticosteroid Receptors. Int. J. Mol. Sci. 2022, 23, 2867. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, Y.; Zhou, Y.; Zhang, Q.; Xu, C. Potential biomarkers of miRNA in non-functional pituitary adenomas. World J. Surg. Oncol. 2021, 19, 270. [Google Scholar] [CrossRef]
- Chen, Y.; Li, B.; Feng, J.; Fang, Q.; Cheng, J.; Xie, W.; Li, C.; Cheng, S.; Zhang, Y.; Gao, H. JAG1, Regulated by microRNA-424-3p, Involved in Tumorigenesis and Epithelial-Mesenchymal Transition of High Proliferative Potential-Pituitary Adenomas. Front. Oncol. 2020, 10, 567021. [Google Scholar] [CrossRef]
- Wei, D.; Yu, Z.; Cheng, Y.; Jiawei, H.; Jian, G.; Hua, G.; Guilan, D. Dysregulated miR-137 and its target EGFR contribute to the progression of pituitary adenomas. Mol. Cell. Endocrinol. 2021, 520, 111083. [Google Scholar] [CrossRef]
- Su, W.J.; Wang, J.S.; Ye, M.D.; Chen, W.L.; Liao, C.X. Expression and clinical significance of miR-193a-3p in invasive pituitary adenomas. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 7673–7680. [Google Scholar] [CrossRef]
- Cai, F.; Dai, C.; Chen, S.; Wu, Q.; Liu, X.; Hong, Y.; Wang, Z.; Li, L.; Yan, W.; Wang, R.; et al. CXCL12-regulated miR-370-3p functions as a tumor suppressor gene by targeting HMGA2 in nonfunctional pituitary adenomas. Mol. Cell. Endocrinol. 2019, 488, 25–35. [Google Scholar] [CrossRef]
- Song, W.; Qian, L.; Jing, G.; Jie, F.; Xiaosong, S.; Chunhui, L.; Yangfang, L.; Guilin, L.; Gao, H.; Yazhuo, Z. Aberrant expression of the sFRP and WIF1 genes in invasive non-functioning pituitary adenomas. Mol. Cell. Endocrinol. 2018, 474, 168–175. [Google Scholar] [CrossRef]
- Leone, V.; Langella, C.; D’Angelo, D.; Mussnich, P.; Wierinckx, A.; Terracciano, L.; Raverot, G.; Lachuer, J.; Rotondi, S.; Jaffrain-Rea, M.L.; et al. Mir-23b and miR-130b expression is downregulated in pituitary adenomas. Mol. Cell. Endocrinol. 2014, 390, 1–7. [Google Scholar] [CrossRef]
- Gentilin, E.; Tagliati, F.; Filieri, C.; Molè, D.; Minoia, M.; Rosaria Ambrosio, M.; Degli Uberti, E.C.; Zatelli, M.C. miR-26a plays an important role in cell cycle regulation in ACTH-secreting pituitary adenomas by modulating protein kinase Cδ. Endocrinology 2013, 154, 1690–1700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trivellin, G.; Butz, H.; Delhove, J.; Igreja, S.; Chahal, H.S.; Zivkovic, V.; McKay, T.; Patócs, A.; Grossman, A.B.; Korbonits, M. MicroRNA miR-107 is overexpressed in pituitary adenomas and inhibits the expression of aryl hydrocarbon receptor-interacting protein in vitro. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E708–E719. [Google Scholar] [CrossRef] [PubMed]
- Butz, H.; Likó, I.; Czirják, S.; Igaz, P.; Korbonits, M.; Rácz, K.; Patócs, A. MicroRNA profile indicates downregulation of the TGFβ pathway in sporadic non-functioning pituitary adenomas. Pituitary 2011, 14, 112–124. [Google Scholar] [CrossRef]
- Butz, H.; Likó, I.; Czirják, S.; Igaz, P.; Khan, M.M.; Zivkovic, V.; Bálint, K.; Korbonits, M.; Rácz, K.; Patócs, A. Down-regulation of Wee1 kinase by a specific subset of microRNA in human sporadic pituitary adenomas. J. Clin. Endocrinol. Metab. 2010, 95, E181–E191. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.R.; Asa, S.L.; Siomi, H.; Siomi, M.C.; Yoshimoto, K.; Yamada, S.; Wang, E.L.; Rahman, M.M.; Inoue, H.; Itakura, M.; et al. Overexpression of HMGA2 relates to reduction of the let-7 and its relationship to clinicopathological features in pituitary adenomas. Mod. Pathol. 2009, 22, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Lyu, L.; Li, H.; Chen, C.; Yu, Y.; Wang, L.; Yin, S.; Hu, Y.; Jiang, S.; Ye, F.; Zhou, P. Exosomal miRNA Profiling is a Potential Screening Route for Non-Functional Pituitary Adenoma. Front. Cell Dev. Biol. 2021, 9, 771354. [Google Scholar] [CrossRef]
- He, C.; Yang, J.; Ding, J.; Li, S.; Wu, H.; Zhou, F.; Teng, L.; Yang, J. MiR-448 targets BLC2 and inhibits the growth of pituitary adenoma cells. Biochem. Cell Biol. 2020, 98, 511–517. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, T.; Wang, Q.; Gao, H. Overexpression of microRNA-34a Attenuates Proliferation and Induces Apoptosis in Pituitary Adenoma Cells via SOX7. Mol. Ther. Oncolytics. 2018, 10, 40–47. [Google Scholar] [CrossRef]
- Fan, X.; Mao, Z.; He, D.; Liao, C.; Jiang, X.; Lei, N.; Hu, B.; Wang, X.; Li, Z.; Lin, Y.; et al. Expression of somatostatin receptor subtype 2 in growth hormone-secreting pituitary adenoma and the regulation of miR-185. J. Endocrinol. Investig. 2015, 38, 1117–1128. [Google Scholar] [CrossRef]
- Liao, C.; Chen, W.; Fan, X.; Jiang, X.; Qiu, L.; Chen, C.; Zhu, Y.; Wang, H. MicroRNA-200c inhibits apoptosis in pituitary adenoma cells by targeting the PTEN/Akt signaling pathway. Oncol. Res. 2013, 21, 129–136. [Google Scholar] [CrossRef]
- Ulitsky, I.; Bartel, D.P. lincRNAs: Genomics, evolution, and mechanisms. Cell 2013, 154, 26–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Chang, H.Y. Genome regulation by long noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef] [PubMed]
- Engreitz, J.M.; Haines, J.E.; Perez, E.M.; Munson, G.; Chen, J.; Kane, M.; McDonel, P.E.; Guttman, M.; Lander, E.S. Local regulation of gene expression by lncRNA promoters, transcription and splicing. Nature 2016, 539, 452–455. [Google Scholar] [CrossRef]
- Willingham, A.T.; Orth, A.P.; Batalov, S.; Peters, E.C.; Wen, B.G.; Aza-Blanc, P.; Hogenesch, J.B.; Schultz, P.G. A strategy for probing the function of noncoding RNAs finds a repressor of NFAT. Science 2005, 309, 1570–1573. [Google Scholar] [CrossRef] [PubMed]
- Carrieri, C.; Cimatti, L.; Biagioli, M.; Beugnet, A.; Zucchelli, S.; Fedele, S.; Pesce, E.; Ferrer, I.; Collavin, L.; Santoro, C.; et al. Long non-coding antisense RNA controls Uchl1 translation through an embedded SINEB2 repeat. Nature 2012, 491, 454–457. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kopp, F.; Chang, T.C.; Sataluri, A.; Chen, B.; Sivakumar, S.; Yu, H.; Xie, Y.; Mendell, J.T. Noncoding RNA NORAD Regulates Genomic Stability by Sequestering PUMILIO Proteins. Cell 2016, 164, 69–80. [Google Scholar] [CrossRef]
- Tripathi, V.; Ellis, J.D.; Shen, Z.; Song, D.Y.; Pan, Q.; Watt, A.T.; Freier, S.M.; Bennett, C.F.; Sharma, A.; Bubulya, P.A.; et al. The nuclear-retained noncoding RNA MALAT1 regulates alternative splicing by modulating SR splicing factor phosphorylation. Mol. Cell 2010, 39, 925–938. [Google Scholar] [CrossRef]
- Chen, C.K.; Blanco, M.; Jackson, C.; Aznauryan, E.; Ollikainen, N.; Surka, C.; Chow, A.; Cerase, A.; McDonel, P.; Guttman, M. Xist recruits the X chromosome to the nuclear lamina to enable chromosome-wide silencing. Science 2016, 354, 468–472. [Google Scholar] [CrossRef]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef]
- Trimarchi, T.; Bilal, E.; Ntziachristos, P.; Fabbri, G.; Dalla-Favera, R.; Tsirigos, A.; Aifantis, I. Genome-wide mapping and characterization of Notch-regulated long noncoding RNAs in acute leukemia. Cell 2014, 158, 593–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, J.F.; Yin, Q.F.; Chen, T.; Zhang, Y.; Zhang, X.O.; Wu, Z.; Zhang, S.; Wang, H.B.; Ge, J.; Lu, X.; et al. Human colorectal cancer-specific CCAT1-L lncRNA regulates long-range chromatin interactions at the MYC locus. Cell Res. 2014, 24, 513–531. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Bond, A.M.; Vangompel, M.J.; Sametsky, E.A.; Clark, M.F.; Savage, J.C.; Disterhoft, J.F.; Kohtz, J.D. Balanced gene regulation by an embryonic brain ncRNA is critical for adult hippocampal GABA circuitry. Nat. Neurosci. 2009, 12, 1020–1027. [Google Scholar] [CrossRef]
- Tsai, M.C.; Manor, O.; Wan, Y.; Mosammaparast, N.; Wang, J.K.; Lan, F.; Shi, Y.; Segal, E.; Chang, H.Y. Long noncoding RNA as modular scaffold of histone modification complexes. Science 2010, 329, 689–693. [Google Scholar] [CrossRef]
- Li, W.; Notani, D.; Ma, Q.; Tanasa, B.; Nunez, E.; Chen, A.Y.; Merkurjev, D.; Zhang, J.; Ohgi, K.; Song, X.; et al. Functional roles of enhancer RNAs for oestrogen-dependent transcriptional activation. Nature 2013, 498, 516–520. [Google Scholar] [CrossRef]
- Zhang, X.; Ji, S.; Cai, G.; Pan, Z.; Han, R.; Yuan, Y.; Xu, S.; Yang, J.; Hu, X.; Chen, M.; et al. H19 Increases IL-17A/IL-23 Releases via Regulating VDR by Interacting with miR675-5p/miR22-5p in Ankylosing Spondylitis. Mol. Ther. Nucleic. Acids 2020, 19, 393–404. [Google Scholar] [CrossRef]
- Wu, Z.; Zheng, Y.; Xie, W.; Li, Q.; Zhang, Y.; Ren, B.; Cai, L.; Cheng, Y.; Tang, H.; Su, Z.; et al. The long noncoding RNA-H19/miRNA-93a/ATG7 axis regulates the sensitivity of pituitary adenomas to dopamine agonists. Mol. Cell. Endocrinol. 2020, 518, 111033. [Google Scholar] [CrossRef]
- Zhu, H.B.; Li, B.; Guo, J.; Miao, Y.Z.; Shen, Y.T.; Zhang, Y.Z.; Zhao, P.; Li, C.Z. LncRNA MEG8 promotes TNF-α expression by sponging miR-454-3p in bone-invasive pituitary adenomas. Aging 2021, 13, 14342–14354. [Google Scholar] [CrossRef]
- Du, Q.; Hoover, A.R.; Dozmorov, I.; Raj, P.; Khan, S.; Molina, E.; Chang, T.C.; de la Morena, M.T.; Cleaver, O.B.; Mendell, J.T.; et al. MIR205HG Is a Long Noncoding RNA that Regulates Growth Hormone and Prolactin Production in the Anterior Pituitary. Dev. Cell 2019, 49, 618–631.e615. [Google Scholar] [CrossRef]
- Cesana, M.; Cacchiarelli, D.; Legnini, I.; Santini, T.; Sthandier, O.; Chinappi, M.; Tramontano, A.; Bozzoni, I. A long noncoding RNA controls muscle differentiation by functioning as a competing endogenous RNA. Cell 2011, 147, 358–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, B.K.; Pfeifer, K.; Dutta, A. The H19 long noncoding RNA gives rise to microRNAs miR-675-3p and miR-675-5p to promote skeletal muscle differentiation and regeneration. Genes Dev. 2014, 28, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, C.; Wang, J.; Song, G.; Zhao, Z.; Wang, H.; Wang, W.; Li, H.; Li, Z.; Miao, Y.; et al. Genome-wide analysis of differentially expressed lncRNAs and mRNAs in primary gonadotrophin adenomas by RNA-seq. Oncotarget 2017, 8, 4585–4606. [Google Scholar] [CrossRef] [PubMed]
- Xing, W.; Qi, Z.; Huang, C.; Zhang, N.; Zhang, W.; Li, Y.; Qiu, M.; Fang, Q.; Hui, G. Genome-wide identification of lncRNAs and mRNAs differentially expressed in non-functioning pituitary adenoma and construction of an lncRNA-mRNA co-expression network. Biol. Open 2019, 8, bio037127. [Google Scholar] [CrossRef]
- Peng, C.; Wang, S.; Yu, J.; Deng, X.; Ye, H.; Chen, Z.; Yao, H.; Cai, H.; Li, Y.; Yuan, Y. lncRNA-mRNA Expression Patterns in Invasive Pituitary Adenomas: A Microarray Analysis. Biomed. Res. Int. 2022, 2022, 1380485. [Google Scholar] [CrossRef]
- Zhu, H.; Guo, J.; Shen, Y.; Dong, W.; Gao, H.; Miao, Y.; Li, C.; Zhang, Y. Functions and Mechanisms of Tumor Necrosis Factor-α and Noncoding RNAs in Bone-Invasive Pituitary Adenomas. Clin. Cancer Res. 2018, 24, 5757–5766. [Google Scholar] [CrossRef]
- Byun, H.M.; Wong, H.L.; Birnstein, E.A.; Wolff, E.M.; Liang, G.; Yang, A.S. Examination of IGF2 and H19 loss of imprinting in bladder cancer. Cancer Res. 2007, 67, 10753–10758. [Google Scholar] [CrossRef]
- Yoshimizu, T.; Miroglio, A.; Ripoche, M.A.; Gabory, A.; Vernucci, M.; Riccio, A.; Colnot, S.; Godard, C.; Terris, B.; Jammes, H.; et al. The H19 locus acts in vivo as a tumor suppressor. Proc. Natl. Acad. Sci. USA 2008, 105, 12417–12422. [Google Scholar] [CrossRef]
- Jiang, X.; Yan, Y.; Hu, M.; Chen, X.; Wang, Y.; Dai, Y.; Wu, D.; Wang, Y.; Zhuang, Z.; Xia, H. Increased level of H19 long noncoding RNA promotes invasion, angiogenesis, and stemness of glioblastoma cells. J. Neurosurg. 2016, 124, 129–136. [Google Scholar] [CrossRef]
- Lu, T.; Yu, C.; Ni, H.; Liang, W.; Yan, H.; Jin, W. Expression of the long non-coding RNA H19 and MALAT-1 in growth hormone-secreting pituitary adenomas and its relationship to tumor behavior. Int. J. Dev. Neurosci. 2018, 67, 46–50. [Google Scholar] [CrossRef]
- Liang, W.; Zou, Y.; Qin, F.; Chen, J.; Xu, J.; Huang, S.; Chen, J.; Dai, S. sTLR4/MD-2 complex inhibits colorectal cancer migration and invasiveness in vitro and in vivo by lncRNA H19 down-regulation. Acta Biochim. Biophys. Sin. 2017, 49, 1035–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.R.; Yan, L.; Liu, Y.T.; Cao, L.; Guo, Y.H.; Zhang, Y.; Yao, H.; Cai, L.; Shang, H.B.; Rui, W.W.; et al. Inhibition of mTORC1 by lncRNA H19 via disrupting 4E-BP1/Raptor interaction in pituitary tumours. Nat. Commun. 2018, 9, 4624. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.T.; Tang, H.; Xie, W.Q.; Yao, H.; Gu, W.T.; Zheng, Y.Z.; Shang, H.B.; Wang, Y.; Wei, Y.X.; et al. Exosome-Transmitted lncRNA H19 Inhibits the Growth of Pituitary Adenoma. J. Clin. Endocrinol. Metab. 2019, 104, 6345–6356. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Guo, J.; Wang, D.; Fang, Q.; Liu, Y.; Xie, W.; Zhang, Y.; Li, C. A Novel Three-LncRNA Signature Predicting Tumor Recurrence in Nonfunctioning Pituitary Adenomas. Front. Genet. 2021, 12, 754503. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Duan, J.; Zhou, D. Long-noncoding RNA IFNG-AS1 exerts oncogenic properties by interacting with epithelial splicing regulatory protein 2 (ESRP2) in pituitary adenomas. Pathol. Res. Pract. 2018, 214, 2054–2061. [Google Scholar] [CrossRef]
- Fu, D.; Zhang, Y.; Cui, H. Long noncoding RNA CCAT2 is activated by E2F1 and exerts oncogenic properties by interacting with PTTG1 in pituitary adenomas. Am. J. Cancer Res. 2018, 8, 245–255. [Google Scholar]
- Li, Z.; Li, C.; Liu, C.; Yu, S.; Zhang, Y. Expression of the long non-coding RNAs MEG3, HOTAIR, and MALAT-1 in non-functioning pituitary adenomas and their relationship to tumor behavior. Pituitary 2015, 18, 42–47. [Google Scholar] [CrossRef]
- Cheng, S.; Guo, J.; Zhang, Y.; Li, Z.; Li, C. Identification of a multidimensional transcriptome signature predicting tumor regrowth of clinically non-functioning pituitary adenoma. Int. J. Oncol. 2020, 57, 804–812. [Google Scholar] [CrossRef]
- Zhang, Y.; Tan, Y.; Wang, H.; Xu, M.; Xu, L. Long Non-Coding RNA Plasmacytoma Variant Translocation 1 (PVT1) Enhances Proliferation, Migration, and Epithelial-Mesenchymal Transition (EMT) of Pituitary Adenoma Cells by Activating β-Catenin, c-Myc, and Cyclin D1 Expression. Med. Sci. Monit. 2019, 25, 7652–7659. [Google Scholar] [CrossRef]
- Liu, C.X.; Chen, L.L. Circular RNAs: Characterization, cellular roles, and applications. Cell 2022, 185, 2016–2034. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, S.; Du, Q.; Bian, P.; Chen, Y.; Liu, Z.; Zheng, J.; Sai, K.; Mou, Y.; Chen, Z.; et al. CircVPS13C promotes pituitary adenoma growth by decreasing the stability of IFITM1 mRNA via interacting with RRBP1. Oncogene 2022, 41, 1550–1562. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.K.; Schimmel, K.; Zhao, L.; Chen, C.K.; Dua, K.; Nicolls, M.R.; Spiekerkoetter, E. The role of circular RNAs in pulmonary hypertension. Eur. Respir. J. 2022, 9, 2200012. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Yang, Y.; Kang, M.; Wang, Y.; Wang, Y.; Bi, Y.; He, S.; Shimamoto, F. m(6)A demethylase ALKBH5 inhibits pancreatic cancer tumorigenesis by decreasing WIF-1 RNA methylation and mediating Wnt signaling. Mol. Cancer 2020, 19, 3. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.; Zheng, H.; Wu, Z.; Chen, M.; Huang, Y. Circular RNA-protein interactions: Functions, mechanisms, and identification. Theranostics 2020, 10, 3503–3517. [Google Scholar] [CrossRef] [PubMed]
- Vo, J.N.; Cieslik, M.; Zhang, Y.; Shukla, S.; Xiao, L.; Zhang, Y.; Wu, Y.M.; Dhanasekaran, S.M.; Engelke, C.G.; Cao, X.; et al. The Landscape of Circular RNA in Cancer. Cell 2019, 176, 869–881.e813. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, Q.; Bao, C.; Li, S.; Guo, W.; Zhao, J.; Chen, D.; Gu, J.; He, X.; Huang, S. Circular RNA is enriched and stable in exosomes: A promising biomarker for cancer diagnosis. Cell Res. 2015, 25, 981–984. [Google Scholar] [CrossRef]
- Yu, J.; Xu, Q.G.; Wang, Z.G.; Yang, Y.; Zhang, L.; Ma, J.Z.; Sun, S.H.; Yang, F.; Zhou, W.P. Circular RNA cSMARCA5 inhibits growth and metastasis in hepatocellular carcinoma. J. Hepatol. 2018, 68, 1214–1227. [Google Scholar] [CrossRef]
- Jahani, S.; Nazeri, E.; Majidzadeh, A.K.; Jahani, M.; Esmaeili, R. Circular RNA; a new biomarker for breast cancer: A systematic review. J. Cell. Physiol. 2020, 235, 5501–5510. [Google Scholar] [CrossRef]
- Guo, J.; Wang, Z.; Miao, Y.; Shen, Y.; Li, M.; Gong, L.; Wang, H.; He, Y.; Gao, H.; Liu, Q.; et al. A two-circRNA signature predicts tumour recurrence in clinical non-functioning pituitary adenoma. Oncol. Rep. 2019, 41, 113–124. [Google Scholar] [CrossRef]
- Du, Q.; Zhang, W.; Feng, Q.; Hao, B.; Cheng, C.; Cheng, Y.; Li, Y.; Fan, X.; Chen, Z. Comprehensive circular RNA profiling reveals that hsa_circ_0001368 is involved in growth hormone-secreting pituitary adenoma development. Brain Res. Bull. 2020, 161, 65–77. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, N.; Zhang, S.; Zhou, P.; Lv, L.; Richard, S.A.; Ma, W.; Chen, C.; Wang, X.; Huang, S.; et al. Differential circular RNA expression profiles of invasive and non-invasive non-functioning pituitary adenomas: A microarray analysis. Medicine 2019, 98, e16148. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, D.; Wan, D.; Ma, Q.; Liu, Q.; Li, J.; Li, Z.; Gao, Y.; Jiang, G.; Ma, L.; et al. Circular RNA In Invasive and Recurrent Clinical Nonfunctioning Pituitary Adenomas: Expression Profiles and Bioinformatic Analysis. World Neurosurg. 2018, 117, e371–e386. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Yang, F.; Fan, H.; Wang, H.; Wang, Q.; Yang, J.; Song, T. Long non-coding RNA TUG1/microRNA-187-3p/TESC axis modulates progression of pituitary adenoma via regulating the NF-κB signaling pathway. Cell Death Dis. 2021, 12, 524. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Zhou, S.; Zheng, Y.; Pan, Z.; Chen, Y.; Wang, X. LncRNA BBOX1-AS1 promotes pituitary adenoma progression via sponging miR-361-3p/E2F1 axis. Anticancer Drugs 2022, 33, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Cai, M.; Chen, C.; Ling, C.; Zhang, B.; Zheng, W.; Luo, L. LINC01116 boosts the progression of pituitary adenoma via regulating miR-744-5p/HOXB8 pathway. Mol. Cell. Endocrinol. 2021, 536, 111350. [Google Scholar] [CrossRef]
- Wang, X.; Li, X.; Wang, Z. lncRNA MEG3 inhibits pituitary tumor development by participating in cell proliferation, apoptosis and EMT processes. Oncol. Rep. 2021, 45, 1–11. [Google Scholar] [CrossRef]
- Zhu, D.; Xiao, Z.; Wang, Z.; Hu, B.; Duan, C.; Zhu, Z.; Gao, N.; Zhu, Y.; Wang, H. MEG3/MIR-376B-3P/HMGA2 axis is involved in pituitary tumor invasiveness. J. Neurosurg. 2020, 134, 499–511. [Google Scholar] [CrossRef]
- Qiu, P.; Bi, J.; Liu, J.; Lai, C.; Li, X. Long non-coding RNA LINC01004 promotes malignant behaviors of pituitary adenoma via miR-323a-3p/136-5p/RCN2 axis. Pathol. Res. Pract. 2022, 234, 153884. [Google Scholar] [CrossRef]
- Li, Z.; Ren, R.; Wang, L.; Wang, Z.; Zong, X.; Sun, P.; Zhu, C.; Guo, M.; Guo, G.; Hu, G.; et al. lncRNA KCNQ1OT1 Promotes EMT, Angiogenesis, and Stemness of Pituitary Adenoma by Upregulation of RAB11A. J. Oncol. 2022, 2022, 4474476. [Google Scholar] [CrossRef]
- Li, J.; Qian, Y.; Zhang, C.; Wang, W.; Qiao, Y.; Song, H.; Li, L.; Guo, J.; Lu, D.; Deng, X. LncRNA LINC00473 is involved in the progression of invasive pituitary adenoma by upregulating KMT5A via ceRNA-mediated miR-502-3p evasion. Cell Death Dis. 2021, 12, 580. [Google Scholar] [CrossRef] [PubMed]
- Mao, D.; Jie, Y.; Lv, Y. LncRNA SNHG6 Induces Epithelial-Mesenchymal Transition of Pituitary Adenoma Via Suppressing MiR-944. Cancer Biother. Radiopharm. 2022, 37, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Hu, B.; Feng, Y.; Wang, Z.; Wang, X.; Zhu, D.; Zhu, Y.; Jiang, X.; Wang, H. circOMA1-Mediated miR-145-5p Suppresses Tumor Growth of Nonfunctioning Pituitary Adenomas by Targeting TPT1. J. Clin. Endocrinol. Metab. 2019, 104, 2419–2434. [Google Scholar] [CrossRef]
- Cheng, J.; Nie, D.; Li, B.; Gui, S.; Li, C.; Zhang, Y.; Zhao, P. CircNFIX promotes progression of pituitary adenoma via CCNB1 by sponging miR-34a -5p. Mol. Cell. Endocrinol. 2021, 525, 111140. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Cheng, J.; Li, B.; Nie, D.; Wang, H.; Li, C.; Gui, S.; Zhang, Y. LncRNA PCAT6 regulates the progression of pituitary adenomas by regulating the miR-139-3p/BRD4 axis. Cancer Cell Int. 2021, 21, 14. [Google Scholar] [CrossRef] [PubMed]
- Yue, X.; Dong, C.; Ye, Z.; Zhu, L.; Zhang, X.; Wang, X.; Mo, F.; Li, Z.; Pan, B. LncRNA SNHG7 sponges miR-449a to promote pituitary adenomas progression. Metab. Brain Dis. 2021, 36, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Zheng, X.; Tang, X.; Zang, Z.; Li, B.; He, S.; Shen, R.; Yang, H.; Li, S. Potential biomarkers and lncRNA-mRNA regulatory networks in invasive growth hormone-secreting pituitary adenomas. J. Endocrinol. Investig. 2021, 44, 1947–1959. [Google Scholar] [CrossRef]
- Kang, Y.; He, W.; Tulley, S.; Gupta, G.P.; Serganova, I.; Chen, C.R.; Manova-Todorova, K.; Blasberg, R.; Gerald, W.L.; Massagué, J. Breast cancer bone metastasis mediated by the Smad tumor suppressor pathway. Proc. Natl. Acad. Sci. USA 2005, 102, 13909–13914. [Google Scholar] [CrossRef]
- Inoue, J.; Inazawa, J. Cancer-associated miRNAs and their therapeutic potential. J. Hum. Genet. 2021, 66, 937–945. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, W.; Cao, L.; Jia, Y.; Xiao, Y.; Zhang, X.; Gui, S. Emerging Roles of miRNA, lncRNA, circRNA, and Their Cross-Talk in Pituitary Adenoma. Cells 2022, 11, 2920. https://doi.org/10.3390/cells11182920
Wu W, Cao L, Jia Y, Xiao Y, Zhang X, Gui S. Emerging Roles of miRNA, lncRNA, circRNA, and Their Cross-Talk in Pituitary Adenoma. Cells. 2022; 11(18):2920. https://doi.org/10.3390/cells11182920
Chicago/Turabian StyleWu, Wentao, Lei Cao, Yanfei Jia, Youchao Xiao, Xu Zhang, and Songbai Gui. 2022. "Emerging Roles of miRNA, lncRNA, circRNA, and Their Cross-Talk in Pituitary Adenoma" Cells 11, no. 18: 2920. https://doi.org/10.3390/cells11182920