
Purkinje Cell Patterning—Insights from Single-Cell Sequencing

Abstract
:1. Introduction
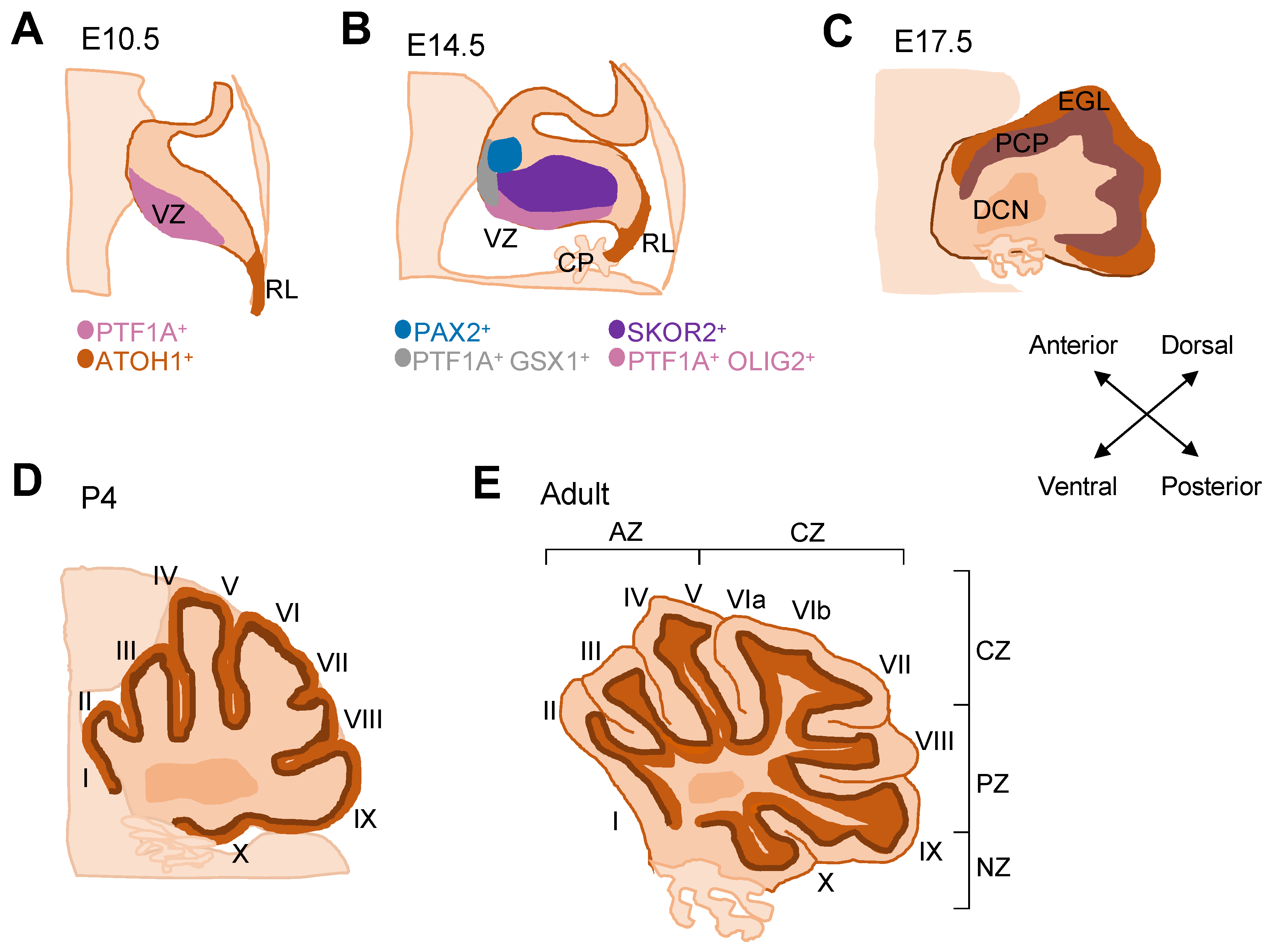
2. Development of the Cerebellum
2.1. Early Cerebellar Development
2.2. Generation of Purkinje Cells
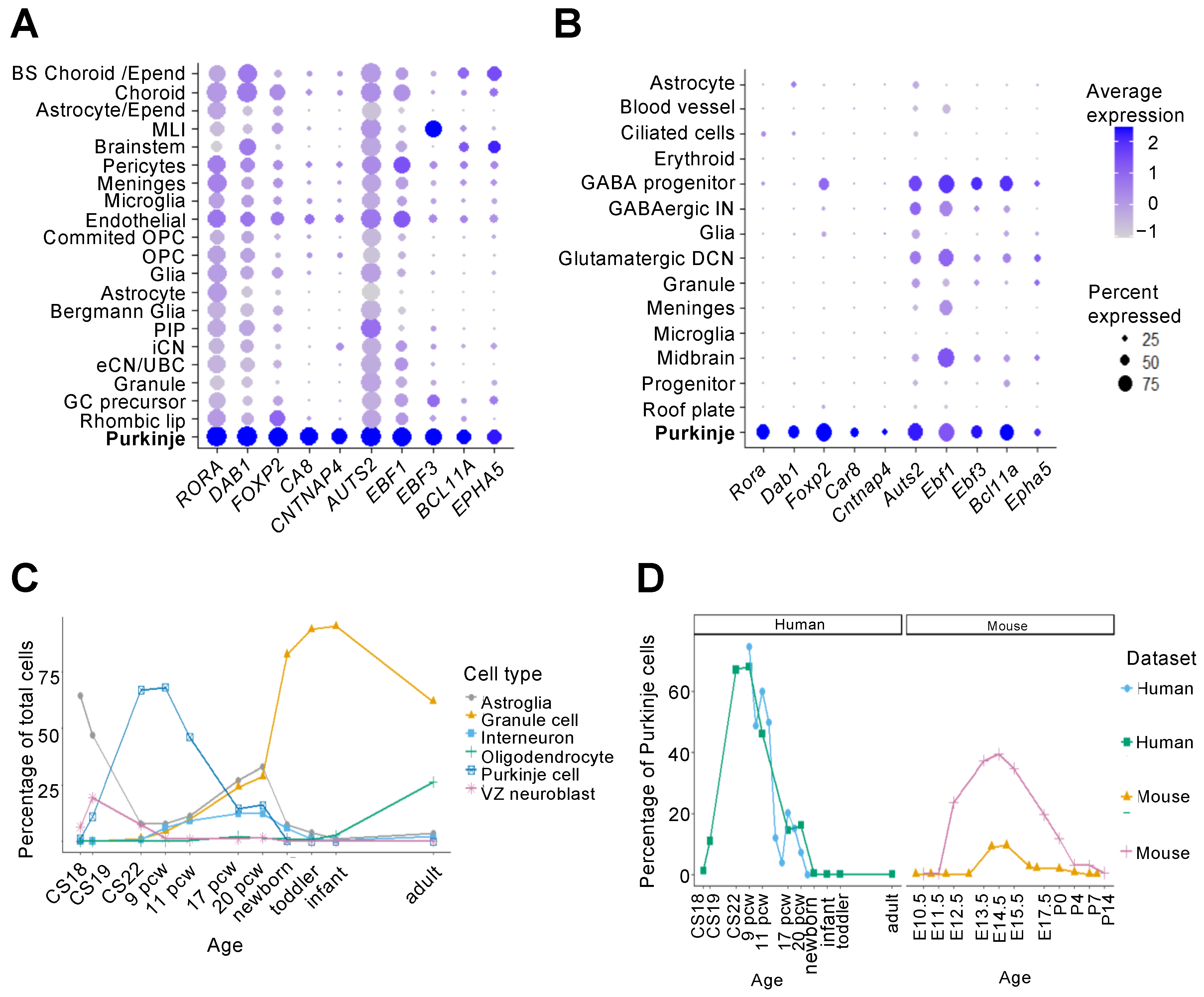
2.3. Recent Transcriptomic Insights into Purkinje Cell Development

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Species | Time Points | Sequencing Method (Library Preparation, Sequencing) | Cell Number 1 | Cell Clusters/Types |
|---|---|---|---|---|---|
| Carter et al., 2018 [25] | Mouse | E10–P10 | scRNA-seq (10x Genomics, Illumina HiSeq 2500) | 39,245 | 48 clusters, 15 cell types |
| Peng et al., 2019 [27] | Mouse | P0, P8 | scRNA-seq (10x Genomics Chromium, Illumina HiSeqX) | 21,119 | 8 cell types |
| Rodriques et al., 2019 [28] | Mouse | Adult | Slide-seq (Illumina NovaSeq) | Not specified | 12 cell types, focus on comparison of Z+ and Z− stripes |
| Vladoiu et al., 2019 [29] | Mouse | E10–P14 | scRNA-seq (10x Genomics Chromium, Illumina 2500) | 62,040 | 30 cell types |
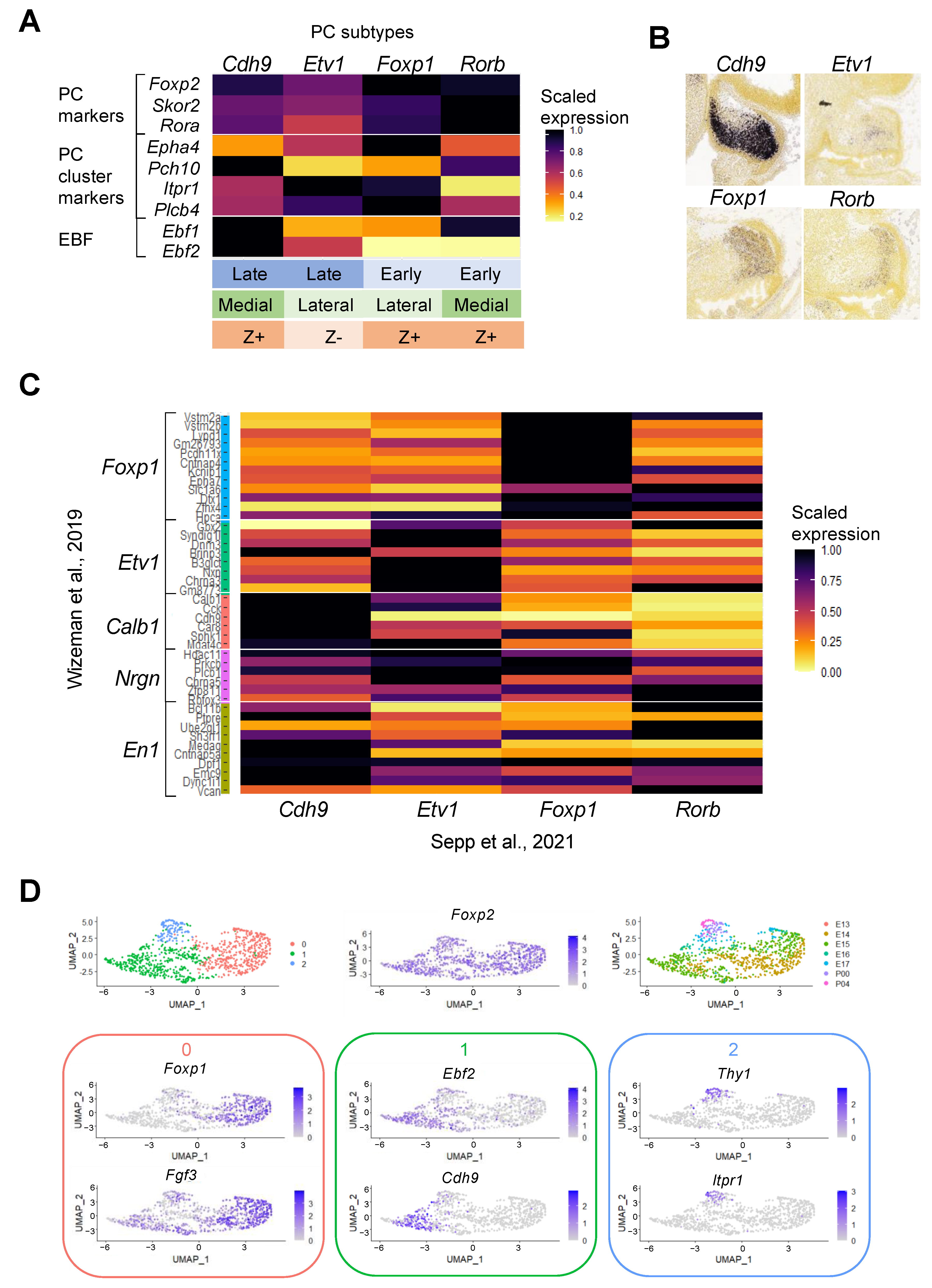
| Wizeman et al., 2019 [30] | Mouse | E13.5 | scRNA-seq (10x Genomics Chromium, Illumina NextSeq 500) | 9326 | 19 clusters |
| Aldinger et al., 2021 [26] | Human | 9–21 pcw | snRNA-seq 2 (SPLiT-seq, Illumina NovaSeq) | 69,174 | 21 cell types |
| Kozareva et al., 2021 [31] | Mouse | Adult (16 different regions) | snRNA-seq (10x Genomics Chromium) | 611,034 | 46 clusters, 18 cell types |
| Sarropoulos et al., 2021 [32] | Mouse | E10.5–P63 | snATAC-seq (10x Genomics, Illumina NextSeq 550) | 91,922 | 12 broad cell types, 42 subtypes and cell states |
| Opossum | P21 & Adult | ||||
| Sepp et al., 2021 [33] | Mouse | E10.5–P63 | snRNA-seq (10x Genomics Chromium, Illumina HiSeq 4000) | 395,736 (115,282 mouse, 180,956 human, 99,498 opossum) | 25 cell types, 44 cell states, (12 cell states further split into 49 subtypes) |
| Human | 7 pcw–adult | ||||
| Opossum | E14–adult | ||||
| Chen et al., 2022 [34] | Mouse | Adult | INTACT snRNA-seq (10x Genomics, Illumina NovaSeq6000) | 52,487 3 | 5 broad cell types (focus on 2 subtypes of Purkinje cells: Z+ and Z−) |
| Khouri-Farah et al., 2022 [35] | Mouse | E12.5–14.5 | scRNA-seq (10x Genomics Chromium) snATAC-seq (10x Genomics) | 31,144 | 26 cell types |
| Gene | avg_log2FC | pct.1 | pct.2 | Gene | avg_log2FC | pct.1 | pct.2 | ||
|---|---|---|---|---|---|---|---|---|---|
| 1 | RORA | 2.31025 | 0.999 | 0.826 | 38 | CADM2 | 0.500989 | 0.917 | 0.766 |
| 2 | DAB1 | 2.254189 | 0.993 | 0.674 | 39 | NRXN3 | 0.493309 | 0.995 | 0.758 |
| 3 | FOXP2 | 1.894787 | 0.966 | 0.418 | 40 | NCAM1 | 0.482498 | 0.949 | 0.785 |
| 4 | CA8 | 1.884526 | 0.895 | 0.148 | 41 | SOX4 | 0.449056 | 0.448 | 0.195 |
| 5 | CNTNAP4 | 1.685789 | 0.801 | 0.171 | 42 | NPTN | 0.443094 | 0.463 | 0.167 |
| 6 | AUTS2 | 1.605596 | 1 | 0.967 | 43 | DCX | 0.429353 | 0.556 | 0.288 |
| 7 | EBF1 | 1.571646 | 0.946 | 0.511 | 44 | KCTD1 | 0.425951 | 0.466 | 0.2 |
| 8 | EBF3 | 1.171463 | 0.825 | 0.252 | 45 | EBF2 | 0.409443 | 0.233 | 0.084 |
| 9 | BCL11A | 1.045316 | 0.744 | 0.202 | 46 | SRGAP1 | 0.392077 | 0.72 | 0.453 |
| 10 | EPHA5 | 0.91761 | 0.727 | 0.268 | 47 | DLGAP4 | 0.391699 | 0.533 | 0.269 |
| 11 | NTM | 0.909009 | 0.985 | 0.797 | 48 | FSTL5 | 0.379821 | 0.667 | 0.373 |
| 12 | CELF2 | 0.863531 | 0.987 | 0.836 | 49 | PHACTR1 | 0.379253 | 0.796 | 0.572 |
| 13 | DNER | 0.80226 | 0.773 | 0.343 | 50 | UBE2E2 | 0.351005 | 0.686 | 0.448 |
| 14 | MEF2C | 0.769929 | 0.555 | 0.099 | 51 | PKIA | 0.338708 | 0.48 | 0.251 |
| 15 | LRRC3B | 0.769179 | 0.566 | 0.132 | 52 | PRKAR2B | 0.329412 | 0.411 | 0.194 |
| 16 | FOXP1 | 0.76819 | 0.65 | 0.309 | 53 | SLIT2 | 0.327772 | 0.543 | 0.317 |
| 17 | TENM2 | 0.767349 | 0.95 | 0.755 | 54 | KIDINS220 | 0.324566 | 0.61 | 0.389 |
| 18 | PPP2R2B | 0.764795 | 0.83 | 0.447 | 55 | ZNRF1 | 0.314214 | 0.431 | 0.209 |
| 19 | KCNIP1 | 0.759729 | 0.744 | 0.306 | 56 | CADM1 | 0.313522 | 0.624 | 0.413 |
| 20 | ATP2B2 | 0.736813 | 0.65 | 0.247 | 57 | SYT16 | 0.310313 | 0.268 | 0.058 |
| 21 | EPHA7 | 0.736577 | 0.685 | 0.365 | 58 | MYT1L | 0.306548 | 0.901 | 0.65 |
| 22 | FOXP4 | 0.707523 | 0.461 | 0.046 | 59 | ARHGAP20 | 0.291987 | 0.269 | 0.093 |
| 23 | XPR1 | 0.700962 | 0.724 | 0.343 | 60 | WNT7B | 0.29075 | 0.216 | 0.026 |
| 24 | PCDH17 | 0.690897 | 0.566 | 0.187 | 61 | CACNG2 | 0.290066 | 0.339 | 0.14 |
| 24 | CNTN5 | 0.667829 | 0.928 | 0.687 | 62 | LYPD1 | 0.289998 | 0.221 | 0.03 |
| 26 | NRP2 | 0.667035 | 0.434 | 0.077 | 63 | GNG2 | 0.289336 | 0.481 | 0.276 |
| 27 | KITLG | 0.659408 | 0.455 | 0.091 | 64 | CRMP1 | 0.282869 | 0.489 | 0.28 |
| 28 | EPHA4 | 0.653552 | 0.482 | 0.124 | 65 | GAD1 | 0.275746 | 0.351 | 0.128 |
| 29 | RBMS1 | 0.627325 | 0.595 | 0.207 | 66 | CMIP | 0.267756 | 0.653 | 0.46 |
| 30 | CTNNA2 | 0.6253 | 0.982 | 0.801 | 67 | PCP4 | 0.264464 | 0.206 | 0.04 |
| 31 | ANK2 | 0.60591 | 0.966 | 0.724 | 68 | NSG1 | 0.263876 | 0.257 | 0.076 |
| 32 | CTTNBP2 | 0.58162 | 0.609 | 0.232 | 69 | SLC1A6 | 0.261146 | 0.197 | 0.029 |
| 33 | SPOCK3 | 0.56822 | 0.538 | 0.215 | 70 | RAB3C | 0.260151 | 0.373 | 0.215 |
| 34 | NTRK3 | 0.531684 | 0.772 | 0.481 | 71 | CHD3 | 0.25663 | 0.349 | 0.165 |
| 35 | PID1 | 0.516531 | 0.44 | 0.174 | 72 | NNAT | 0.256533 | 0.346 | 0.182 |
| 36 | LHX1 | 0.512295 | 0.402 | 0.073 | 73 | EVL | 0.251227 | 0.529 | 0.335 |
| 37 | MACROD2 | 0.511606 | 0.889 | 0.667 |
3. Purkinje Cell Complexity
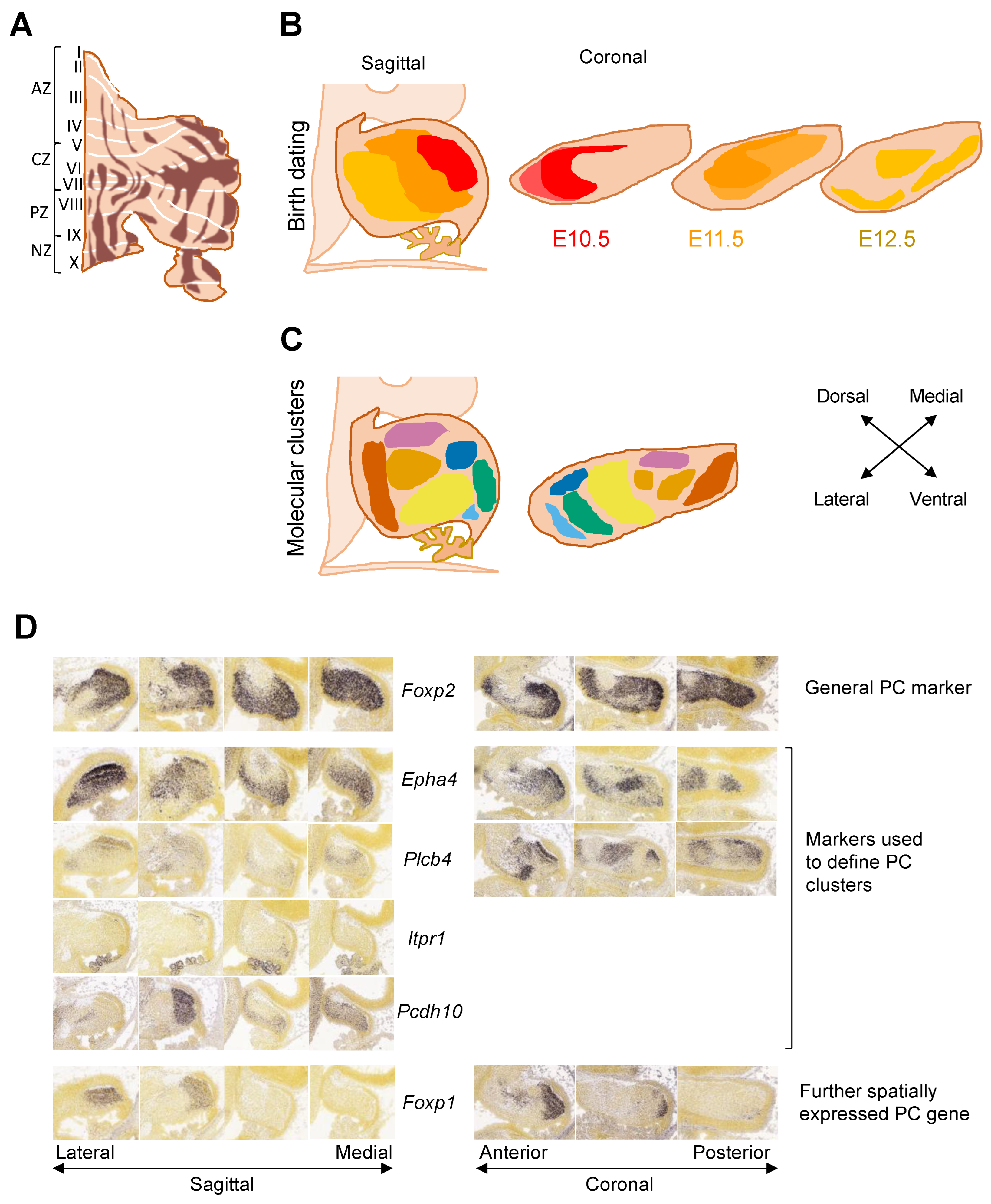
3.1. Adult Cerebellar Topography
3.2. From Cells to Clusters to Stripes
3.3. Understanding Purkinje Cell Identity Using Single-Cell Transcriptomics
4. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Overwalle, F.; Manto, M.; Cattaneo, Z.; Clausi, S.; Ferrari, C.; Gabrieli, J.D.E.; Guell, X.; Heleven, E.; Lupo, M.; Ma, Q.; et al. Consensus Paper: Cerebellum and Social Cognition. Cerebellum 2020, 19, 833–868. [Google Scholar] [CrossRef] [PubMed]
- Stoodley, C.J.; Schmahmann, J.D. Evidence for Topographic Organization in the Cerebellum of Motor Control versus Cognitive and Affective Processing. Cortex 2010, 46, 831–844. [Google Scholar] [CrossRef] [PubMed]
- Schmahmann, J.D.; Guell, X.; Stoodley, C.J.; Halko, M.A. The Theory and Neuroscience of Cerebellar Cognition. Annu. Rev. Neurosci. 2019, 42, 337–364. [Google Scholar] [CrossRef] [PubMed]
- Stoodley, C.J.; Limperopoulos, C. Structure–Function Relationships in the Developing Cerebellum: Evidence from Early-Life Cerebellar Injury and Neurodevelopmental Disorders. Semin. Fetal Neonatal Med. 2016, 21, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Sathyanesan, A.; Zhou, J.; Scafidi, J.; Heck, D.H.; Sillitoe, R.V.; Gallo, V. Emerging Connections between Cerebellar Development, Behaviour and Complex Brain Disorders. Nat. Rev. Neurosci. 2019, 20, 298–313. [Google Scholar] [CrossRef]
- van der Heijden, M.E.; Sillitoe, R.V. Interactions between Purkinje Cells and Granule Cells Coordinate the Development of Functional Cerebellar Circuits. Neuroscience 2021, 462, 4–21. [Google Scholar] [CrossRef]
- Brochu, G.; Maler, L.; Hawkes, R. Zebrin II: A Polypeptide Antigen Expressed Selectively by Purkinje Cells Reveals Compartments in Rat and Fish Cerebellum. J. Comp. Neurol. 1990, 291, 538–552. [Google Scholar] [CrossRef]
- Cerminara, N.L.; Lang, E.J.; Sillitoe, R.V.; Apps, R. Redefining the Cerebellar Cortex. Nat. Publ. Gr. 2015, 16, 79–93. [Google Scholar] [CrossRef]
- White, J.J.; Sillitoe, R.V. Development of the Cerebellum: From Gene Expression Patterns to Circuit Maps. Wiley Interdiscip. Rev. Dev. Biol. 2013, 2, 149–164. [Google Scholar] [CrossRef]
- Butts, T.; Green, M.J.; Wingate, R.J.T. Development of the Cerebellum: Simple Steps to Make a ‘Little Brain’. Development 2014, 141, 4031–4041. [Google Scholar] [CrossRef] [Green Version]
- Leto, K.; Arancillo, M.; Becker, E.B.E.; Buffo, A.; Chiang, C.; Ding, B.; Dobyns, W.B.; Dusart, I.; Haldipur, P.; Hatten, M.E.; et al. Consensus Paper: Cerebellar Development. Cerebellum 2016, 15, 789–828. [Google Scholar] [CrossRef] [PubMed]
- Wang, V.Y.; Zoghbi, H.Y. Genetic Regulation of Cerebellar Development. Nat. Rev. Neurosci. 2001, 2, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Herculano-Houzel, S. Coordinated Scaling of Cortical and Cerebellar Numbers of Neurons. Front. Neuroanat. 2010, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Crossley, P.H.; Martinez, S.; Martin, G.R. Midbrain Development Induced by FGF8 in the Chick Embryo. Nature 1996, 380, 66–68. [Google Scholar] [CrossRef] [PubMed]
- Martinez, S.; Crossley, P.H.; Cobos, I.; Rubenstein, J.L.R.; Martin, G.R. FGF8 Induces Formation of an Ectopic Isthmic Organizer and Isthmocerebellar Development via a Repressive Effect on Otx2 Expression. Development 1999, 126, 1189–1200. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Losos, K.; Joyner, A.L. FGF8 Can Activate Gbx2 and Transform Regions of the Rostral Mouse Brain into a Hindbrain Fate. Development 1999, 126, 4827–4838. [Google Scholar] [CrossRef]
- MacHold, R.; Fishell, G. Math1 Is Expressed in Temporally Discrete Pools of Cerebellar Rhombic-Lip Neural Progenitors. Neuron 2005, 48, 17–24. [Google Scholar] [CrossRef]
- Hoshino, M.; Nakamura, S.; Mori, K.; Kawauchi, T.; Terao, M.; Nishimura, Y.V.; Fukuda, A.; Fuse, T.; Matsuo, N.; Sone, M.; et al. Ptf1a, a BHLH Transcriptional Gene, Defines GABAergic Neuronal Fates in Cerebellum. Neuron 2005, 47, 201–213. [Google Scholar] [CrossRef]
- Pascual, M.; Abasolo, I.; Meur, A.M.-L.; Martínez, A.; Del Rio, J.A.; Wright, C.V.E.; Real, F.X.; Soriano, E. Cerebellar GABAergic Progenitors Adopt an External Granule Cell-like Phenotype in the Absence of Ptf1a Transcription Factor Expression. Proc. Natl. Acad. Sci. USA 2007, 104, 5193–5198. [Google Scholar] [CrossRef]
- Seto, Y.; Nakatani, T.; Masuyama, N.; Taya, S.; Kumai, M.; Minaki, Y.; Hamaguchi, A.; Inoue, Y.U.; Inoue, T.; Miyashita, S.; et al. Temporal Identity Transition from Purkinje Cell Progenitors to GABAergic Interneuron Progenitors in the Cerebellum. Nat. Commun. 2014, 5, 3337. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, M.; Mikoshiba, K. Mediolateral Compartmentalization of the Cerebellum Is Determined on the “Birth Date” of Purkinje Cells. J. Neurosci. 2003, 23, 11342–11351. [Google Scholar] [CrossRef] [PubMed]
- Beekhof, G.C.; Osório, C.; White, J.J.; Van Zoomeren, S.; Van der Stok, H.; Xiong, B.; Nettersheim, I.H.M.S.; Mak, W.A.; Runge, M.; Fiocchi, F.R.; et al. Differential Spatiotemporal Development of Purkinje Cell Populations and Cerebellum-Dependent Sensorimotor Behaviors. Elife 2021, 10, e63668. [Google Scholar] [CrossRef] [PubMed]
- Haldipur, P.; Millen, K.J.; Aldinger, K.A. Human Cerebellar Development and Transcriptomics: Implications for Neurodevelopmental Disorders. Annu. Rev. Neurosci. 2022, 45, 515–531. [Google Scholar] [CrossRef]
- Haldipur, P.; Aldinger, K.A.; Bernardo, S.; Deng, M.; Timms, A.E.; Overman, L.M.; Winter, C.; Lisgo, S.N.; Razavi, F.; Silvestri, E.; et al. Spatiotemporal Expansion of Primary Progenitor Zones in the Developing Human Cerebellum. Science 2019, 366, 454–460. [Google Scholar] [CrossRef]
- Carter, R.A.; Bihannic, L.; Rosencrance, C.; Hadley, J.L.; Tong, Y.; Phoenix, T.N.; Natarajan, S.; Easton, J.; Northcott, P.A.; Gawad, C. A Single-Cell Transcriptional Atlas of the Developing Murine Cerebellum. Curr. Biol. 2018, 28, 2910–2920.e2. [Google Scholar] [CrossRef] [PubMed]
- Aldinger, K.A.; Thomson, Z.; Phelps, I.G.; Haldipur, P.; Deng, M.; Timms, A.E.; Hirano, M.; Santpere, G.; Roco, C.; Rosenberg, A.B.; et al. Spatial and Cell Type Transcriptional Landscape of Human Cerebellar Development. Nat. Neurosci. 2021, 24, 1163–1175. [Google Scholar] [CrossRef]
- Peng, J.; Sheng, A.L.; Xiao, Q.; Shen, L.; Ju, X.C.; Zhang, M.; He, S.T.; Wu, C.; Luo, Z.G. Single-Cell Transcriptomes Reveal Molecular Specializations of Neuronal Cell Types in the Developing Cerebellum. J. Mol. Cell Biol. 2019, 11, 636–648. [Google Scholar] [CrossRef]
- Rodriques, S.G.; Stickels, R.R.; Goeva, A.; Martin, C.A.; Murray, E.; Vanderburg, C.R.; Welch, J.; Chen, L.M.; Chen, F.; Macosko, E.Z. Slide-Seq: A Scalable Technology for Measuring Genome-Wide Expression at High Spatial Resolution. Science 2019, 363, 1463–1467. [Google Scholar] [CrossRef]
- Vladoiu, M.C.; El-Hamamy, I.; Donovan, L.K.; Farooq, H.; Holgado, B.L.; Sundaravadanam, Y.; Ramaswamy, V.; Hendrikse, L.D.; Kumar, S.; Mack, S.C.; et al. Childhood Cerebellar Tumours Mirror Conserved Fetal Transcriptional Programs. Nature 2019, 572, 67–73. [Google Scholar] [CrossRef]
- Wizeman, J.W.; Guo, Q.; Wilion, E.M.; Li, J.Y.H. Specification of Diverse Cell Types during Early Neurogenesis of the Mouse Cerebellum. Elife 2019, 8, e42388. [Google Scholar] [CrossRef]
- Kozareva, V.; Martin, C.; Osorno, T.; Rudolph, S.; Guo, C.; Vanderburg, C.; Nadaf, N.; Regev, A.; Regehr, W.G.; Macosko, E. A Transcriptomic Atlas of Mouse Cerebellar Cortex Comprehensively Defines Cell Types. Nature 2021, 598, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Sarropoulos, I.; Sepp, M.; Frömel, R.; Leiss, K.; Trost, N.; Leushkin, E.; Okonechnikov, K.; Joshi, P.; Giere, P.; Kutscher, L.M.; et al. Developmental and Evolutionary Dynamics of Cis-Regulatory Elements in Mouse Cerebellar Cells. Science 2021, 373, 983. [Google Scholar] [CrossRef] [PubMed]
- Sepp, M.; Leiss, K.; Sarropoulos, I.; Murat, F.; Okonechnikov, K.; Joshi, P.; Leushkin, E.; Mbengue, N.; Schneider, C.; Schmidt, J.; et al. Cellular Development and Evolution of the Mammalian Cerebellum. bioRxiv 2021. [Google Scholar] [CrossRef]
- Chen, X.; Du, Y.; Broussard, G.J.; Kislin, M.; Yuede, C.M.; Zhang, S.; Dietmann, S.; Gabel, H.; Zhao, G.; Wang, S.S.-H.; et al. Transcriptomic Mapping Uncovers Purkinje Neuron Plasticity Driving Learning. Nature 2022, 605, 722–727. [Google Scholar] [CrossRef]
- Khouri-Farah, N.; Guo, Q.; Morgan, K.; Shin, J.; Li, J.Y.H. Integrated Single-Cell Transcriptomic and Epigenetic Study of Cell State Transition and Lineage Commitment in Embryonic Mouse Cerebellum. Sci. Adv. 2022, 8, eabl9156. [Google Scholar] [CrossRef] [PubMed]
- Aldinger, K.A.; Timms, A.E.; Thomson, Z.; Mirzaa, G.M.; Bennett, J.T.; Rosenberg, A.B.; Roco, C.M.; Hirano, M.; Abidi, F.; Haldipur, P.; et al. Redefining the Etiologic Landscape of Cerebellar Malformations. Am. J. Hum. Genet. 2019, 105, 606–615. [Google Scholar] [CrossRef]
- Cunningham, F.; Allen, J.E.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Bennett, R.; et al. Ensembl 2022. Nucleic Acids Res. 2022, 50, D988–D995. [Google Scholar] [CrossRef]
- Hawkes, R. Cerebellar Patterning Defects in Mutant Mice. Front. Neurosci. 2021, 15, 787425. [Google Scholar] [CrossRef]
- Márquez, B.T.; Cook, A.A.; Rice, M.; Smileski, A.; Vieira-Lomasney, K.; Charron, F.; McKinney, R.A.; Watt, A.J. Molecular Identity and Location Influence Purkinje Cell Vulnerability in Autosomal-Recessive Spastic Ataxia of Charlevoix-Saguenay Mice. Front. Cell. Neurosci. 2021, 15, 485. [Google Scholar] [CrossRef]
- Sarna, J.R.; Larouche, M.; Marzban, H.; Sillitoe, R.V.; Rancourt, D.E.; Hawkes, R. Patterned Purkinje Cell Degeneration in Mouse Models of Niemann-Pick Type C Disease. J. Comp. Neurol. 2003, 456, 279–291. [Google Scholar] [CrossRef]
- Apps, R.; Hawkes, R.; Aoki, S.; Bengtsson, F.; Brown, A.M.; Chen, G.; Ebner, T.J.; Isope, P.; Jörntell, H.; Lackey, E.P.; et al. Cerebellar Modules and Their Role as Operational Cerebellar Processing Units. Cerebellum 2018, 17, 654–682. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Morita, N.; Furuichi, T.; Sugihara, I. Clustered Fine Compartmentalization of the Mouse Embryonic Cerebellar Cortex and Its Rearrangement into the Postnatal Striped Configuration. J. Neurosci. 2012, 32, 15688–15703. [Google Scholar] [CrossRef]
- Sillitoe, R.V.; Marzban, H.; Larouche, M.; Zahedi, S.; Affanni, J.; Hawkes, R. Conservation of the Architecture of the Anterior Lobe Vermis of the Cerebellum across Mammalian Species. Prog. Brain Res. 2005, 148, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, C.L.; Krueger-Naug, A.M.; Currie, R.W.; Hawkes, R. Constitutive Expression of the 25-Kda Heat Shock Protein Hsp25 Reveals Novel Parasagittal Bands of Purkinje Cells in the Adult Mouse Cerebellar Cortex. J. Comp. Neurol. 2000, 416, 383–397. [Google Scholar] [CrossRef]
- White, J.J.; Sillitoe, R.V. Postnatal Development of Cerebellar Zones Revealed by Neurofilament Heavy Chain Protein Expression. Front. Neuroanat. 2013, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Brandenburg, C.; Smith, L.A.; Kilander, M.B.C.; Bridi, M.S.; Lin, Y.C.; Huang, S.; Blatt, G.J. Parvalbumin Subtypes of Cerebellar Purkinje Cells Contribute to Differential Intrinsic Firing Properties. Mol. Cell. Neurosci. 2021, 115, 103650. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, N.; Schwarting, G.A.; Herrup, K.; Hawkes, R.; Yamamoto, M. Compartmentation in Mammalian Cerebellum: Zebrin II and P-Path Antibodies Define Three Classes of Sagittally Organized Bands of Purkinje Cells. Proc. Natl. Acad. Sci. USA 1992, 89, 5006–5010. [Google Scholar] [CrossRef]
- Hawkes, R. The Ferdinando Rossi Memorial Lecture: Zones and Stripes—Pattern Formation in the Cerebellum. Cerebellum 2018, 17, 12–16. [Google Scholar] [CrossRef]
- Nguyen-Minh, V.; Tran-Anh, K.; Sugihara, I. Heterogeneity of Intrinsic Excitability in Purkinje Cells Linked with Longitudinal Zebrin Zones in the Mouse Cerebellum. bioarXiv 2020. [Google Scholar] [CrossRef]
- Zhou, H.; Lin, Z.; Voges, K.; Ju, C.; Gao, Z.; Bosman, L.W.J.; Ruigrok, T.J.H.; Hoebeek, F.E.; Zeeuw, C.I. De Cerebellar Modules Operate at Different Frequencies. Elife 2014, 3, e02536. [Google Scholar] [CrossRef]
- Wu, B.; Blot, F.G.C.; Wong, A.B.; Osório, C.; Adolfs, Y.; Pasterkamp, R.J.; Hartmann, J.; Becker, E.B.E.; Boele, H.J.; De Zeeuw, C.I.; et al. TRPC3 is a Major Contributor to Functional Heterogeneity of Cerebellar Purkinje Cells. Elife 2019, 8, e45590. [Google Scholar] [CrossRef] [PubMed]
- Vibulyaseck, S.; Fujita, H.; Luo, Y.; Tran, A.K.; Oh-Nishi, A.; Ono, Y.; Hirano, S.; Sugihara, I. Spatial Rearrangement of Purkinje Cell Subsets Forms the Transverse and Longitudinal Compartmentalization in the Mouse Embryonic Cerebellum. J. Comp. Neurol. 2017, 525, 2971–2990. [Google Scholar] [CrossRef] [PubMed]
- Tran-Anh, K.; Zhang, J.; Nguyen-Minh, V.T.; Fujita, H.; Hirata, T.; Sugihara, I. Common Origin of the Cerebellar Dual Somatotopic Areas Revealed by Tracking Embryonic Purkinje Cell Clusters with Birthdate Tagging. eNeuro 2020, 7, 1–23. [Google Scholar] [CrossRef]
- Morales, D.; Hatten, M.E. Molecular Markers of Neuronal Progenitors in the Embryonic Cerebellar Anlage. J. Neurosci. 2006, 26, 12226–12236. [Google Scholar] [CrossRef]
- Miyata, T.; Ono, Y.; Okamoto, M.; Masaoka, M.; Sakakibara, A.; Kawaguchi, A.; Hashimoto, M.; Ogawa, M. Migration, Early Axonogenesis, and Reelin-Dependent Layer-Forming Behavior of Early/Posterior-Born Purkinje Cells in the Developing Mouse Lateral Cerebellum. Neural Dev. 2010, 5, 23. [Google Scholar] [CrossRef]
- Fujita, H.; Sugihara, I. FoxP2 Expression in the Cerebellum and Inferior Olive: Development of the Transverse Stripe-Shaped Expression Pattern in the Mouse Cerebellar Cortex. J. Comp. Neurol. 2012, 520, 656–677. [Google Scholar] [CrossRef] [PubMed]
- Tissir, F.; Goffinet, A.M. Reelin and Brain Development. Nat. Rev. Neurosci. 2003, 4, 496–505. [Google Scholar] [CrossRef] [PubMed]
- D’Arcangelo, G.; Miao, G.G.; Chen, S.C.; Scares, H.D.; Morgan, J.I.; Curran, T. A Protein Related to Extracellular Matrix Proteins Deleted in the Mouse Mutant Reeler. Nature 1995, 374, 719–723. [Google Scholar] [CrossRef]
- Sillitoe, R.; Gopal, N.; Joyner, A.L. Embryonic Origins of ZebrinII Parasagittal Stripes and Establishment of Topographic Purkinje Cell Projections. Neuroscience 2009, 162, 574–588. [Google Scholar] [CrossRef]
- Cheng, Y.; Sudarov, A.; Szulc, K.U.; Sgaier, S.K.; Stephen, D.; Turnbull, D.H.; Joyner, A.L. The Engrailed Homeobox Genes Determine the Different Foliation Patterns in the Vermis and Hemispheres of the Mammalian Cerebellum. Development 2010, 137, 519–529. [Google Scholar] [CrossRef] [Green Version]
- Sillitoe, R.V.; Stephen, D.; Lao, Z.; Joyner, A.L. Engrailed Homeobox Genes Determine the Organization of Purkinje Cell Sagittal Stripe Gene Expression in the Adult Cerebellum. J. Neurosci. 2008, 28, 12150–12162. [Google Scholar] [CrossRef]
- Wilson, S.L.; Kalinovsky, A.; Orvis, G.D.; Joyner, A.L. Spatially Restricted and Developmentally Dynamic Expression of Engrailed Genes in Multiple Cerebellar Cell Types. Cerebellum 2011, 10, 356–372. [Google Scholar] [CrossRef] [PubMed]
- Redies, C.; Neudert, F.; Lin, J. Cadherins in Cerebellar Development: Translation of Embryonic Patterning into Mature Functional Compartmentalization. Cerebellum 2011, 10, 393–408. [Google Scholar] [CrossRef] [PubMed]
- Casoni, F.; Croci, L.; Cremona, O.; Hawkes, R.; Consalez, G.G. Early Purkinje Cell Development and the Origins of Cerebellar Patterning. In Development of the Cerebellum from Molecular Aspects to Diseases; Springer: Berlin/Heidelberg, Germany, 2017; pp. 67–86. [Google Scholar] [CrossRef]
- Croci, L.; Chung, S.; Masserdotti, G.; Gianola, S.; Bizzoca, A.; Gennarini, G.; Corradi, A.; Rossi, F.; Hawkes, R.; Consalez, G.G. A Key Role for the HLH Transcription Factor EBF2 COE2, O/E-3 in Purkinje Neuron Migration and Cerebellar Cortical Topography. Development 2006, 2729, 2719–2729. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Marzban, H.; Croci, L. Purkinje Cell Subtype Specification in the Cerebellar Cortex: Early B-Cell Factor 2 Acts to Repress the Zebrin-II Positive Purkinje Cell Phenotype. Neuroscience 2008, 153, 721–732. [Google Scholar] [CrossRef]
- Badaloni, A.; Casoni, F.; Croci, L.; Chiara, F.; Bizzoca, A.; Gennarini, G.; Cremona, O.; Hawkes, R.; Consalez, G.G. Dynamic Expression and New Functions of Early B Cell Factor 2 in Cerebellar Development. Cerebellum 2019, 18, 999–1010. [Google Scholar] [CrossRef]
- Ståhl, P.L.; Salmén, F.; Vickovic, S.; Lundmark, A.; Navarro, J.F.; Magnusson, J.; Giacomello, S.; Asp, M.; Westholm, J.O.; Huss, M.; et al. Visualization and Analysis of Gene Expression in Tissue Sections by Spatial Transcriptomics. Science 2016, 353, 78–82. [Google Scholar] [CrossRef]
- Lipovsek, M.; Bardy, C.; Cadwell, C.R.; Hadley, K.; Kobak, D.; Tripathy, S.J. Patch-Seq: Past, Present, and Future. J. Neurosci. 2021, 41, 937–946. [Google Scholar] [CrossRef]
- Muguruma, K.; Nishiyama, A.; Kawakami, H.; Hashimoto, K.; Sasai, Y. Self-Organization of Polarized Cerebellar Tissue in 3D Culture of Human Pluripotent Stem Cells. Cell Rep. 2015, 10, 537–550. [Google Scholar] [CrossRef] [Green Version]
- Nayler, S.; Agarwal, D.; Curion, F.; Bowden, R.; Becker, E.B.E. Single-Cell Sequencing of Human IPSC-Derived Cerebellar Organoids Shows Recapitulation of Cerebellar Development. Sci. Rep. 2021, 11, 1–30. [Google Scholar] [CrossRef]
- Buchholz, D.E.; Carroll, T.; Kocabas, A.; Zhu, X.; Behesti, H.; Faust, P.; Stalbow, L.; Fang, Y.; Hatten, M.E. Novel Genetic Features of Human and Mouse Purkinje Cell Differentiation Defined by Comparative Transcriptomics. Pnas 2020, 117, 15085–15095. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.P.; Bekman, E.P.; Fernandes, T.G.; Vaz, S.H.; Rodrigues, C.A.V.; Diogo, M.M.; Cabral, J.M.S.; Carmo-Fonseca, M. Maturation of Human Pluripotent Stem Cell-Derived Cerebellar Neurons in the Absence of Co-Culture. Front. Bioeng. Biotechnol. 2020, 8, 70. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Apsley, E.J.; Becker, E.B.E. Purkinje Cell Patterning—Insights from Single-Cell Sequencing. Cells 2022, 11, 2918. https://doi.org/10.3390/cells11182918
Apsley EJ, Becker EBE. Purkinje Cell Patterning—Insights from Single-Cell Sequencing. Cells. 2022; 11(18):2918. https://doi.org/10.3390/cells11182918
Chicago/Turabian StyleApsley, Elizabeth J., and Esther B. E. Becker. 2022. "Purkinje Cell Patterning—Insights from Single-Cell Sequencing" Cells 11, no. 18: 2918. https://doi.org/10.3390/cells11182918