

Reduced miR-146a-5p Is a Biomarker of Infant Respiratory Diseases Contributing to Immune Dysregulation in Small Airway Epithelial Cells

 , , , ,
, , , ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Sample Collection and Virus Detection
2.3. Nasopharyngeal Aspirate (NPA) Processing
2.4. Small Airway Epithelial Cell Culture
2.5. Gene Expression Assays
2.6. MicroRNA Analysis in NPA Samples
2.7. Statistical Analysis
3. Results
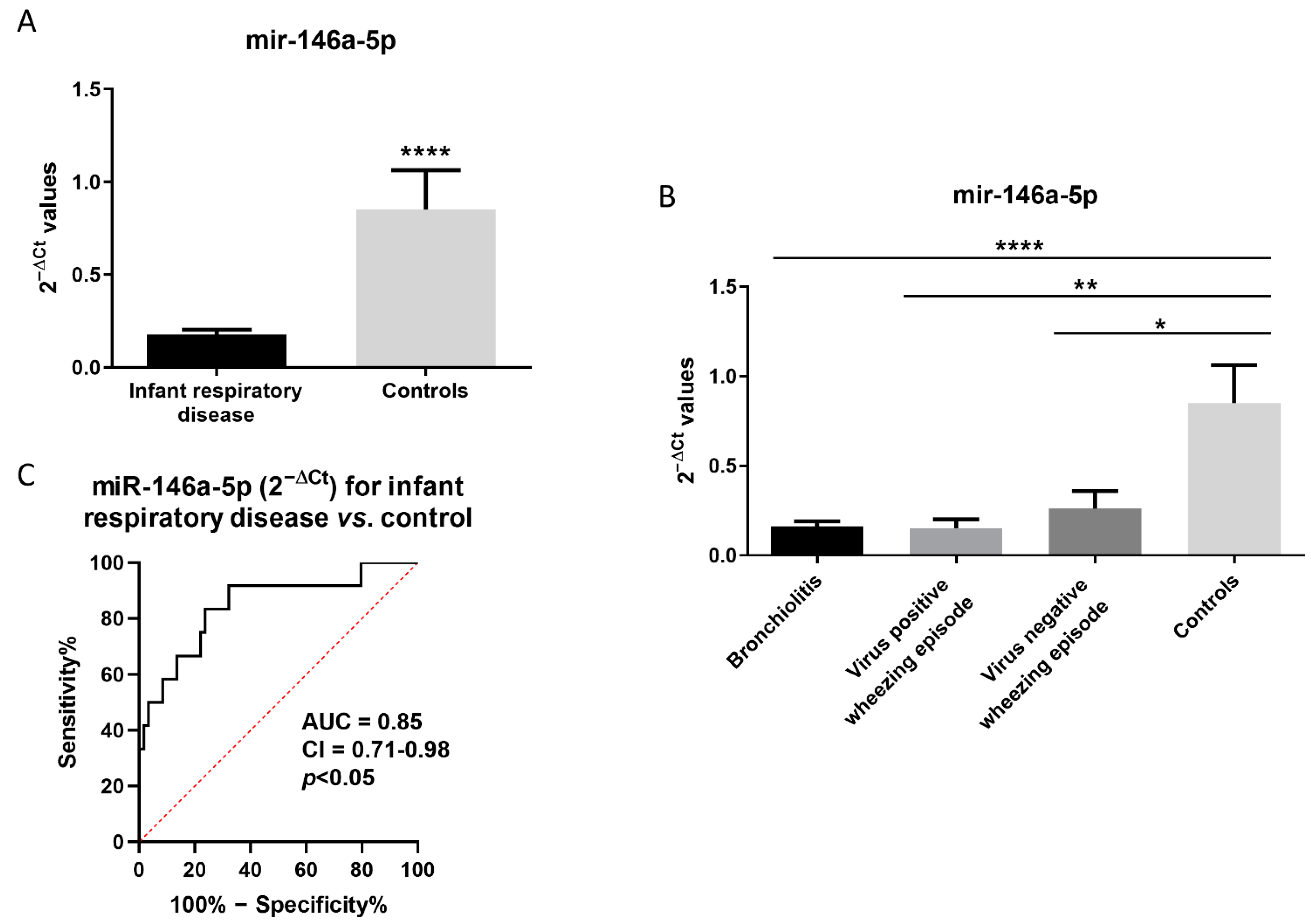
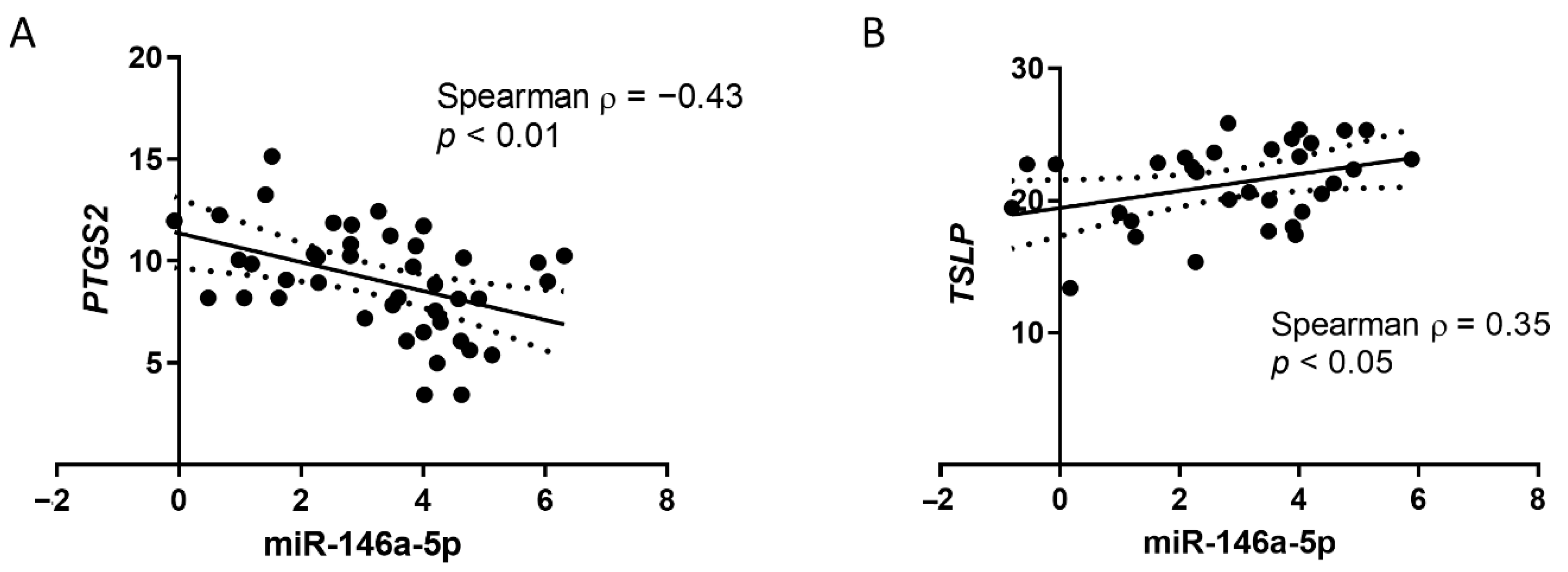
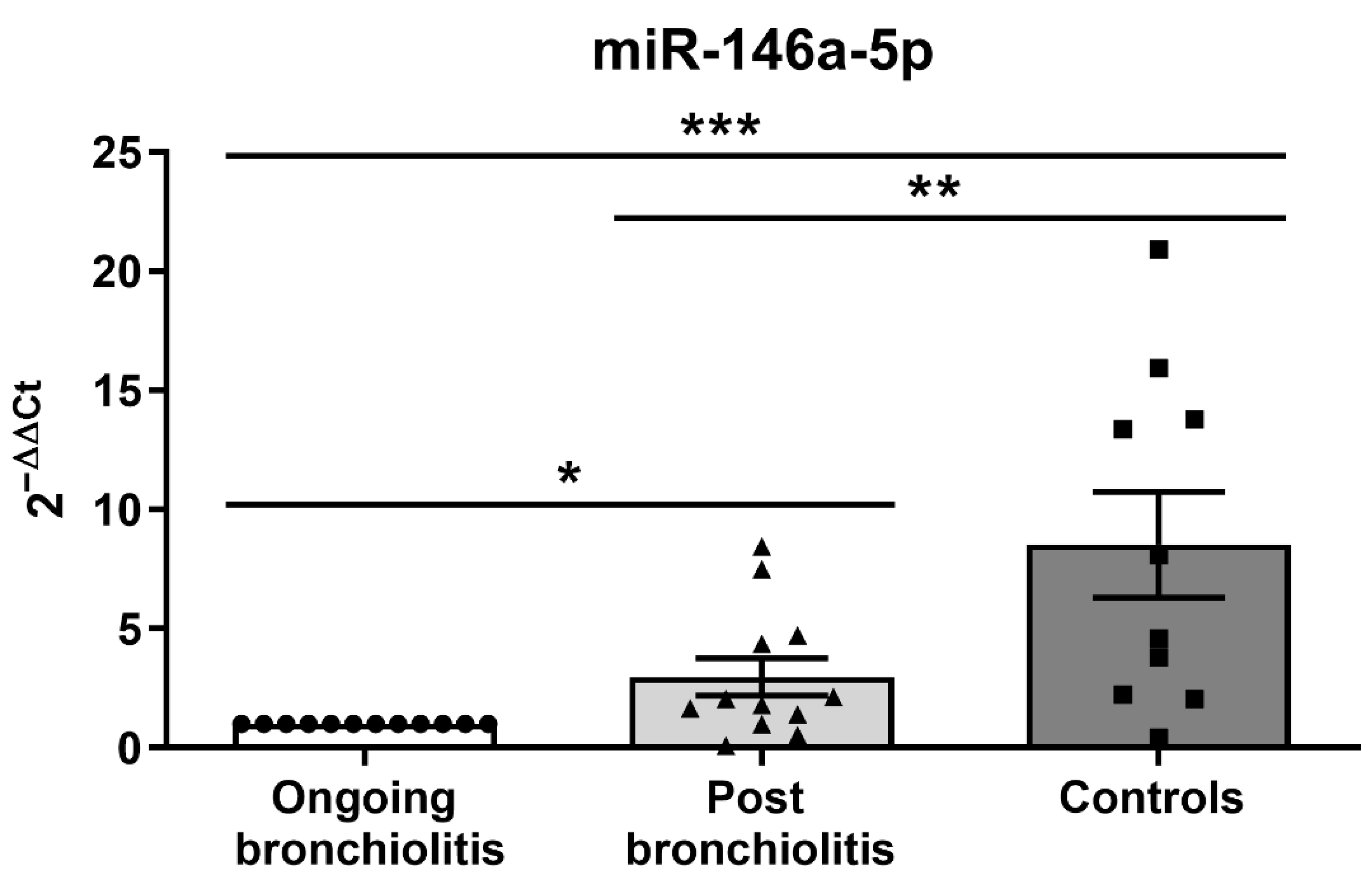
3.1. MiR-146a-5p Is Reduced in NPAs from Infants with Respiratory Diseases and Correlates with Immune-Related Genes
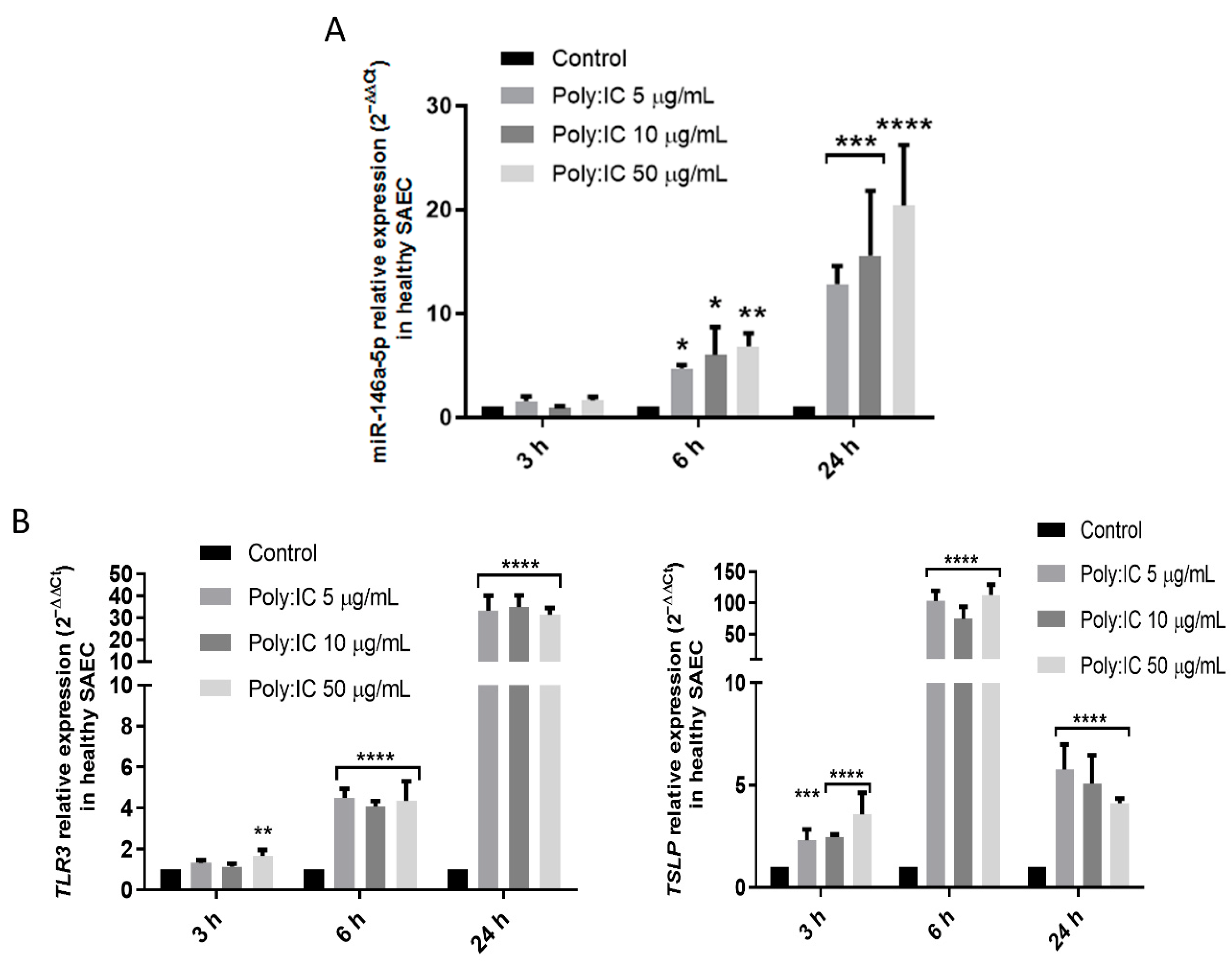
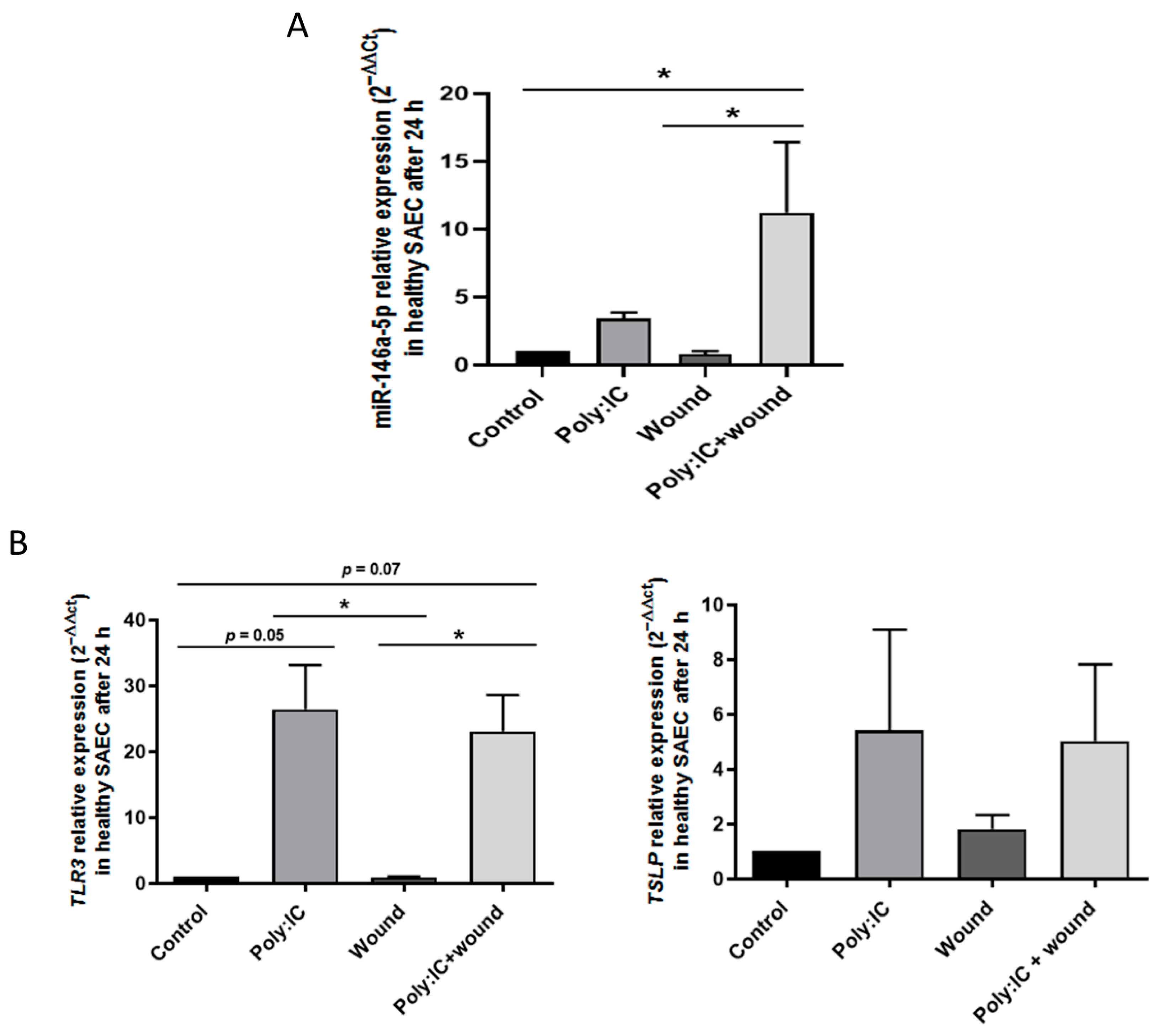
3.2. Viral Stimuli Increase miR-146a-5p Expression in Small Airway Epithelial Cells from Controls
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bousquet, J.; Burney, P.G.; Zuberbier, T.; Cauwenberge, P.V.; Akdis, C.A.; Bindslev-Jensen, C.; Bonini, S.; Fokkens, W.J.; Kauffmann, F.; Kowalski, M.L.; et al. GA2LEN (Global Allergy and Asthma European Network) addresses the allergy and asthma ‘epidemic’. Allergy 2009, 64, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Cutrera, R.; Baraldi, E.; Indinnimeo, L.; Miraglia Del Giudice, M.; Piacentini, G.; Scaglione, F.; Ullmann, N.; Moschino, L.; Galdo, F.; Duse, M. Management of acute respiratory diseases in the pediatric population: The role of oral corticosteroids. Ital. J. Pediatr. 2017, 43, 31. [Google Scholar] [CrossRef]
- Ralston, S.L.; Lieberthal, A.S.; Meissner, H.C.; Alverson, B.K.; Baley, J.E.; Gadomski, A.M.; Johnson, D.W.; Light, M.J.; Maraqa, N.F.; Mendonca, E.A.; et al. Clinical practice guideline: The diagnosis, management, and prevention of bronchiolitis. Pediatrics 2014, 134, e1474–e1502. [Google Scholar] [CrossRef] [PubMed]
- Lieberthal, A.S.; Bauchner, H.; Hall, C.B.; Johnson, D.W.; Kotagal, U.; Light, M.J.; Mason, W.; Meissner, H.C.; Phelan, K.J.; Zorc, J.J.; et al. Diagnosis and management of bronchiolitis. Pediatrics 2006, 118, 1774–1793. [Google Scholar] [CrossRef]
- Mazur, N.I.; Martinón-Torres, F.; Baraldi, E.; Fauroux, B.; Greenough, A.; Heikkinen, T.; Manzoni, P.; Mejias, A.; Nair, H.; Papadopoulos, N.G.; et al. Lower respiratory tract infection caused by respiratory syncytial virus: Current management and new therapeutics. Lancet. Respir. Med. 2015, 3, 888–900. [Google Scholar] [CrossRef]
- Øymar, K.; Skjerven, H.O.; Mikalsen, I.B. Acute bronchiolitis in infants, a review. Scand. J. Trauma. Resusc. Emerg. Med. 2014, 22, 23. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Garcia, M.L.; Calvo, C.; Ruiz, S.; Pozo, F.; Del Pozo, V.; Remedios, L.; Exposito, N.; Tellez, A.; Casas, I. Role of viral coinfections in asthma development. PLoS ONE 2017, 12, e0189083. [Google Scholar] [CrossRef]
- Wu, P.; Hartert, T.V. Evidence for a causal relationship between respiratory syncytial virus infection and asthma. Expert Rev Anti Infect Ther. 2014, 9, 731–745. [Google Scholar] [CrossRef]
- Garcia-Garcia, M.L.; Gonzalez-Carrasco, E.; Bracamonte, T.; Molinero, M.; Pozo, F.; Casas, I.; Calvo, C. Impact of Prematurity and Severe Viral Bronchiolitis on Asthma Development at 6-9 Years. J. Asthma Allergy 2020, 13, 343–353. [Google Scholar] [CrossRef]
- Mallol, J.; García-Marcos, L.; Solé, D.; Brand, P.; Baeza-Bacab, M.; Leite, Á.M.; Bessa, O.C.; Prestes, E.X.; Sarinho, E.; Medeiros, D.; et al. International prevalence of recurrent wheezing during the first year of life: Variability, treatment patterns and use of health resources. Thorax 2010, 65, 1004–1009. [Google Scholar] [CrossRef] [Green Version]
- Sastre, B.; García-García, M.L.; Cañas, J.A.; Calvo, C.; Rodrigo-Muñoz, J.M.; Casas, I.; Mahíllo-Fernández, I.; del Pozo, V. Bronchiolitis and recurrent wheezing are distinguished by type 2 innate lymphoid cells and immune response. Pediatr. Allergy Immunol. 2021, 32, 51–59. [Google Scholar] [CrossRef]
- Chen, S.; Gu, W.; Wu, M.; Hao, C.; Zhu, C.; Shao, X.; Wang, Y. Risk factors for recurrent wheezing after bronchiolitis in infants: 2-year follow up in China. BMC Infect. Dis. 2021, 21, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Yarza, E.G.; Moreno-Galdó, A.; Ramilo, O.; Rubí, T.; Escribano, A.; Torres, A.; Sardón, O.; Oliva, C.; Pérez, G.; Cortell, I.; et al. Risk factors for bronchiolitis, recurrent wheezing, and related hospitalization in preterm infants during the first year of life. Pediatr. Allergy Immunol. 2015, 26, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Pasanen, A.; Karjalainen, M.K.; Bont, L.; Piippo-Savolainen, E.; Ruotsalainen, M.; Goksör, E.; Kumawat, K.; Hodemaekers, H.; Nuolivirta, K.; Jartti, T.; et al. Genome-Wide Association Study of Polymorphisms Predisposing to Bronchiolitis. Sci. Reports 2017, 7, srep41653. [Google Scholar] [CrossRef] [PubMed]
- Drysdale, S.B.; Prendergast, M.; Alcazar, M.; Wilson, T.; Smith, M.; Zuckerman, M.; Broughton, S.; Rafferty, G.F.; Johnston, S.L.; Hodemaekers, H.M.; et al. Genetic predisposition of RSV infection-related respiratory morbidity in preterm infants. Eur. J. Pediatr. 2014, 173, 905–912. [Google Scholar] [CrossRef]
- Cañas, J.A.; Rodrigo-Muñoz, J.M.; Sastre, B.; Gil-Martinez, M.; Redondo, N.; del Pozo, V. MicroRNAs as Potential Regulators of Immune Response Networks in Asthma and Chronic Obstructive Pulmonary Disease. Front. Immunol. 2020, 11, 1. [Google Scholar] [CrossRef]
- Saba, R.; Sorensen, D.L.; Booth, S.A. MicroRNA-146a: A dominant, negative regulator of the innate immune response. Front. Immunol. 2014, 5, 578. [Google Scholar] [CrossRef]
- Mcconnochie, K.M. Bronchiolitis: What’s in the Name? Am. J. Dis. Child. 1983, 137, 11–13. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Jia, Y.; Wei, Y. Modulators of MicroRNA Function in the Immune System. Int. J. Mol. Sci. 2020, 21, 2357. [Google Scholar] [CrossRef]
- Fei, Y.; Chaulagain, A.; Wang, T.; Chen, Y.; Liu, J.; Yi, M.; Wang, Y.; Huang, Y.; Lin, L.; Chen, S.; et al. MiR-146a down-regulates inflammatory response by targeting TLR3 and TRAF6 in Coxsackievirus B infection. RNA 2020, 26, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Bandiera, S.; Pernot, S.; El Saghire, H.; Durand, S.C.; Thumann, C.; Crouchet, E.; Ye, T.; Fofana, I.; Oudot, M.A.; Barths, J.; et al. Hepatitis C Virus-Induced Upregulation of MicroRNA miR-146a-5p in Hepatocytes Promotes Viral Infection and Deregulates Metabolic Pathways Associated with Liver Disease Pathogenesis. J. Virol. 2016, 90, 6387–6400. [Google Scholar] [CrossRef]
- Alexander, M.; Hu, R.; Runtsch, M.C.; Kagele, D.A.; Mosbruger, T.L.; Tolmachova, T.; Seabra, M.C.; Round, J.L.; Ward, D.M.; O’Connell, R.M. Exosome-delivered microRNAs modulate the inflammatory response to endotoxin. Nat. Commun. 2015 61 2015, 6, 7321. [Google Scholar] [CrossRef] [PubMed]
- Iyer, A.; Zurolo, E.; Prabowo, A.; Fluiter, K.; Spliet, W.G.M.; van Rijen, P.C.; Gorter, J.A.; Aronica, E. MicroRNA-146a: A Key Regulator of Astrocyte-Mediated Inflammatory Response. PLoS ONE 2012, 7, e44789. [Google Scholar] [CrossRef]
- Matysiak, M.; Fortak-Michalska, M.; Szymanska, B.; Orlowski, W.; Jurewicz, A.; Selmaj, K. MicroRNA-146a Negatively Regulates the Immunoregulatory Activity of Bone Marrow Stem Cells by Targeting Prostaglandin E2 Synthase-2. J. Immunol. 2013, 190, 5102–5109. [Google Scholar] [CrossRef]
- Kivihall, A.; Aab, A.; Soja, J.; Sładek, K.; Sanak, M.; Altraja, A.; Jakiela, B.; Bochenek, G.; Rebane, A. Reduced expression of miR-146a in human bronchial epithelial cells alters neutrophil migration. Clin. Transl. Allergy 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Comer, B.S.; Camoretti-Mercado, B.; Kogut, P.C.; Halayko, A.J.; Solway, J.; Gerthoffer, W.T. MicroRNA-146a and microRNA-146b expression and anti-inflammatory function in human airway smooth muscle. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2014, 307, L727–L734. [Google Scholar] [CrossRef]
- Panganiban, R.P.; Wang, Y.; Howrylak, J.; Chinchilli, V.M.; Craig, T.J.; August, A.; Ishmael, F.T. Circulating microRNAs as biomarkers in patients with allergic rhinitis and asthma. J. Allergy Clin. Immunol. 2016, 137, 1423–1432. [Google Scholar] [CrossRef]
- Hammad Mahmoud Hammad, R.; Hamed, D.H.E.D.; Eldosoky, M.A.E.R.; Ahmad, A.A.E.S.; Osman, H.M.; Abd Elgalil, H.M.; Mahmoud Hassan, M.M. Plasma microRNA-21, microRNA-146a and IL-13 expression in asthmatic children. Innate Immun. 2018, 24, 171–179. [Google Scholar] [CrossRef]
- Weidner, J.; Ekerljung, L.; Malmhäll, C.; Miron, N.; Rådinger, M. Circulating microRNAs correlate to clinical parameters in individuals with allergic and non-allergic asthma. Respir. Res. 2020, 21, 1–12. [Google Scholar] [CrossRef]
- Lambert, K.A.; Roff, A.N.; Panganiban, R.P.; Douglas, S.; Ishmael, F.T. MicroRNA-146a is induced by inflammatory stimuli in airway epithelial cells and augments the anti-inflammatory effects of glucocorticoids. PLoS ONE 2018, 13, e0205434. [Google Scholar] [CrossRef] [PubMed]
- Laanesoo, A.; Urgard, E.; Periyasamy, K.; Laan, M.; Bochkov, Y.A.; Aab, A.; Magilnick, N.; Pooga, M.; Gern, J.E.; Johnston, S.L.; et al. Dual role of the miR-146 family in rhinovirus-induced airway inflammation and allergic asthma exacerbation. Clin. Transl. Med. 2021, 11, e427. [Google Scholar] [CrossRef] [PubMed]
- Miyata, R.; Kakuki, T.; Nomura, K.; Ohkuni, T.; Ogasawara, N.; Takano, K.I.; Konno, T.; Kohno, T.; Sawada, N.; Himi, T.; et al. Poly(I:C) induced microRNA-146a regulates epithelial barrier and secretion of proinflammatory cytokines in human nasal epithelial cells. Eur. J. Pharmacol. 2015, 761, 375–382. [Google Scholar] [CrossRef]
- Chahar, H.S.; Corsello, T.; Kudlicki, A.S.; Komaravelli, N.; Casola, A. Respiratory Syncytial Virus Infection Changes Cargo Composition of Exosome Released from Airway Epithelial Cells. Sci. Reports 2018, 8, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Liu, X.; Wu, X.; Chen, M.; Yu, W. MiR-146a alleviates lung injury caused by RSV infection in young rats by targeting TRAF-6 and regulating JNK/ERKMAPK signaling pathways. Sci. Reports 2022, 12, 1–12. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Controls (n = 10) | Bronchiolitis (n = 41) | Viral-Positive, Wheezing Episode (n = 10) | Viral-Negative, Wheezing Episode (n = 10) | |
|---|---|---|---|---|
| Age (mo) | 4.9 ± 0.9 | 3.5 ± 0.4 | 33.3 ± 11.1 | 16.9 ± 3.8 |
| Male (%) | 60.0 (6/10) | 51.2 (21/41) | 77.8 (7/9) | 40.0 (4/10) |
| Prematurity (%) | NA | 4.1 (1/24) | 11.1 (1/9) | 0.0 (0/10) |
| Hospital stay (d) | NA | 5.1 ± 0.6 | 2.6 ± 0.5 | 2.7 ± 0.5 |
| Temperature > 37.9 °C (%) | NA | 12.5 (3/24) | 55.6 (5/9) | 33.3 (3/9) |
| Hypoxia (SatO2 < 95%) (%) | NA | 79.2 (19/24) | 88.8 (8/9) | 55.5 (5/9) |
| Neonatal admission (%) | NA | 0.0 (0/24) | 11.1 (1/9) | 0.0 (0/9) |
| Neonatal CPAP (%) | NA | 0.0 (0/22) | 11.1 (1/9) | 0.0 (0/9) |
| Antibiotic treatment (%) | NA | 8.3 (2/24) | 11.1 (1/9) | 22.2 (2/9) |
| Virus (%) | NA | 95.1 (39/41) | 100.0 (10/10) | 0.0 (0/10) |
| RSV | NA | 58.5 (24/41) | 50.0 (5/10) | NA |
| HRV | NA | 34.1 (14/41) | 60.0 (6/19) | NA |
| Coinfection | NA | 24.4 (10/41) | 20.0 (2/19) | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigo-Muñoz, J.M.; Gil-Martínez, M.; Lorente-Sorolla, C.; Sastre, B.; García-García, M.L.; Calvo, C.; Casas, I.; del Pozo, V. Reduced miR-146a-5p Is a Biomarker of Infant Respiratory Diseases Contributing to Immune Dysregulation in Small Airway Epithelial Cells. Cells 2022, 11, 2746. https://doi.org/10.3390/cells11172746
Rodrigo-Muñoz JM, Gil-Martínez M, Lorente-Sorolla C, Sastre B, García-García ML, Calvo C, Casas I, del Pozo V. Reduced miR-146a-5p Is a Biomarker of Infant Respiratory Diseases Contributing to Immune Dysregulation in Small Airway Epithelial Cells. Cells. 2022; 11(17):2746. https://doi.org/10.3390/cells11172746
Chicago/Turabian StyleRodrigo-Muñoz, José M., Marta Gil-Martínez, Clara Lorente-Sorolla, Beatriz Sastre, María Luz García-García, Cristina Calvo, Inmaculada Casas, and Victoria del Pozo. 2022. "Reduced miR-146a-5p Is a Biomarker of Infant Respiratory Diseases Contributing to Immune Dysregulation in Small Airway Epithelial Cells" Cells 11, no. 17: 2746. https://doi.org/10.3390/cells11172746