
Efp/TRIM25 and Its Related Protein, TRIM47, in Hormone-Dependent Cancers


Abstract
:1. Introduction
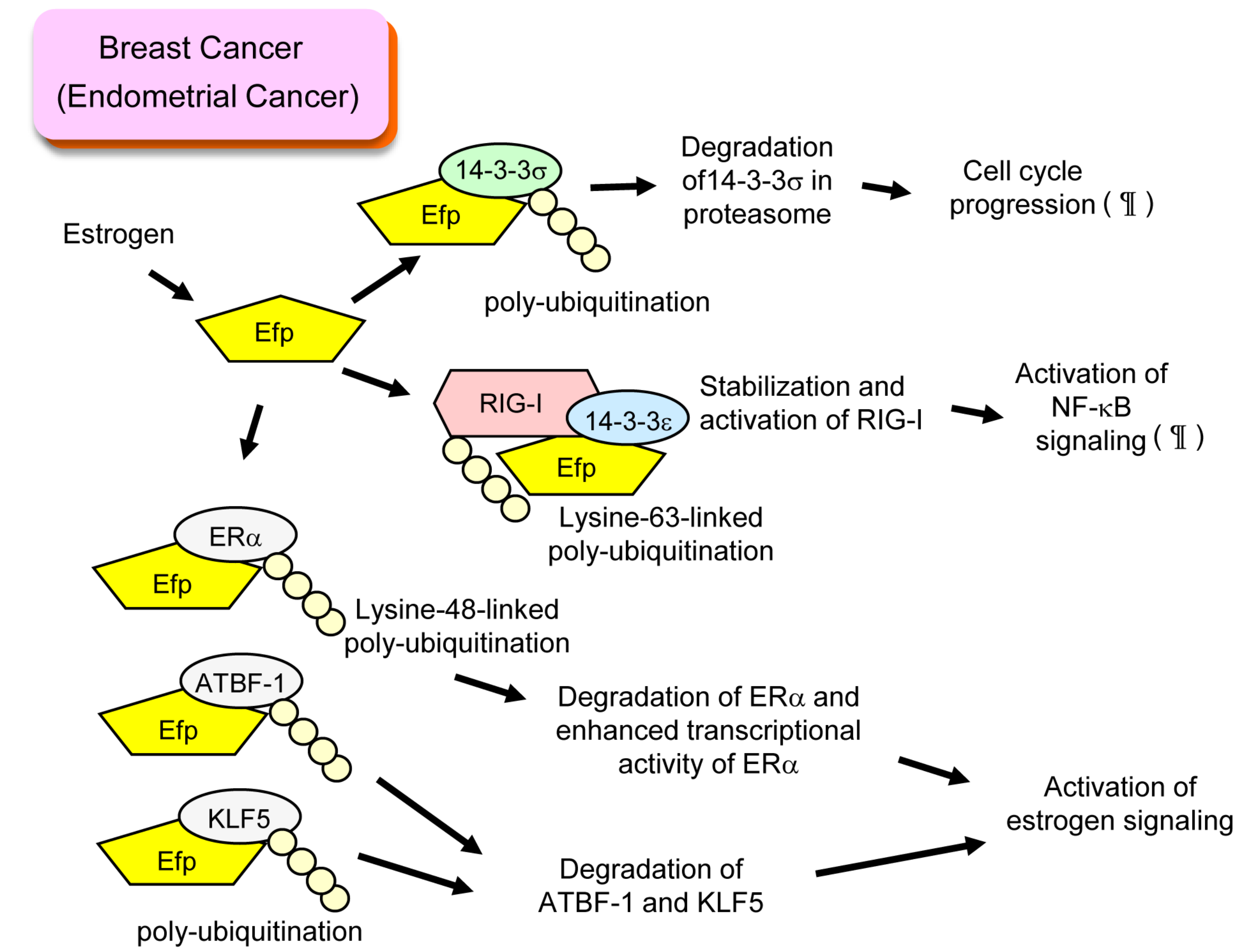
2. Efp and Breast Cancer
3. Efp and Endometrial Cancer
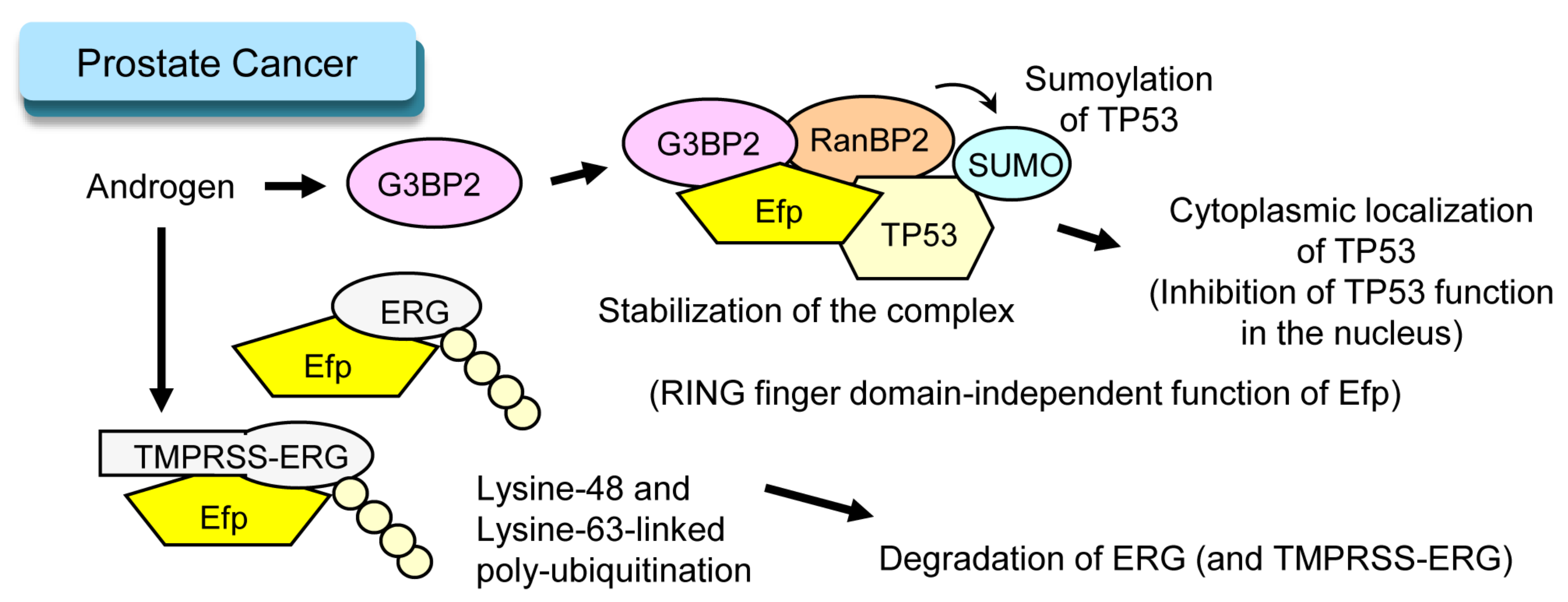
4. Efp and Prostate Cancer
5. Roles of Efp in Other Cancers
6. Roles of Efp in Innate Immunity
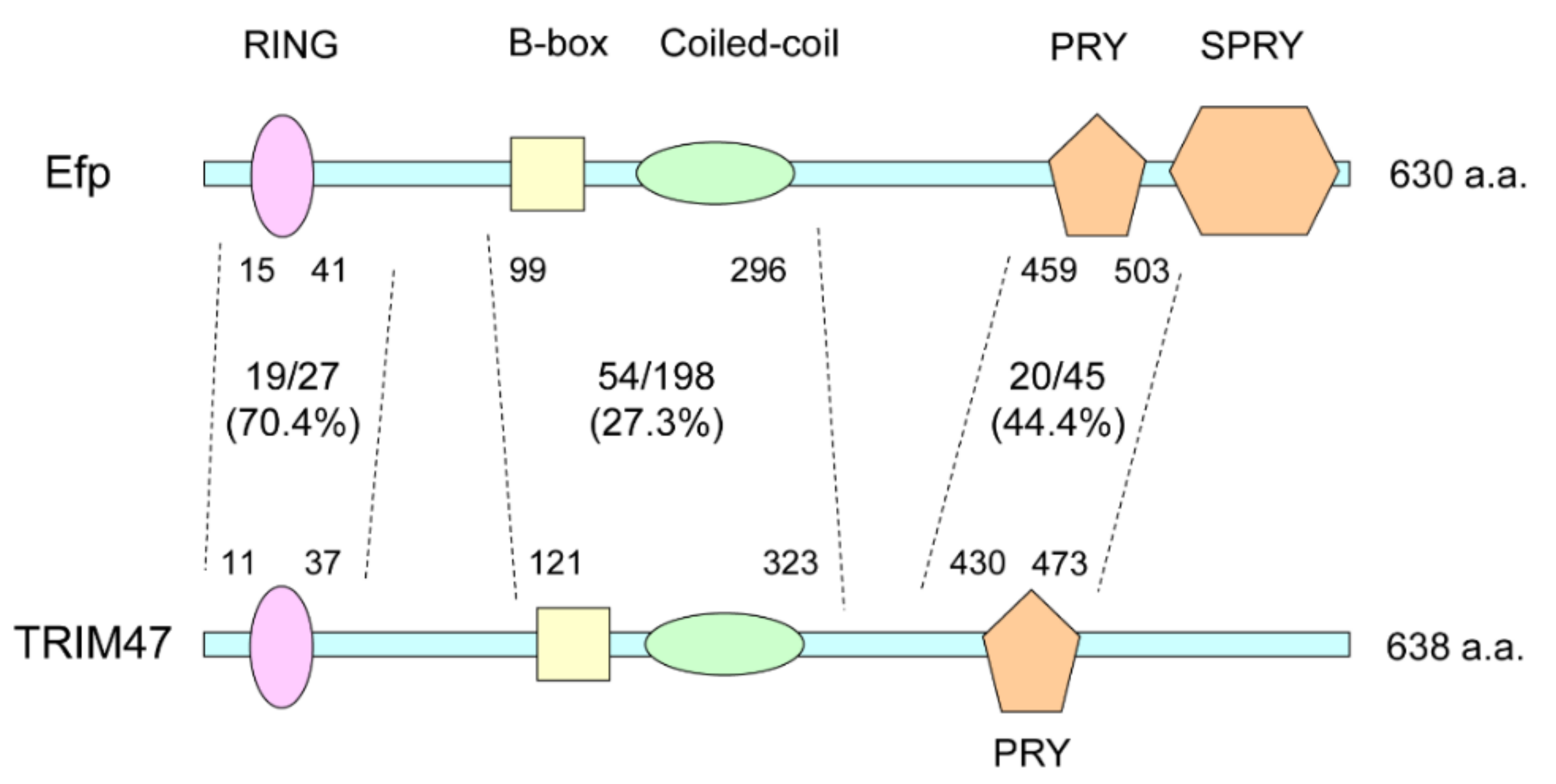
7. TRIM47 and Cancers
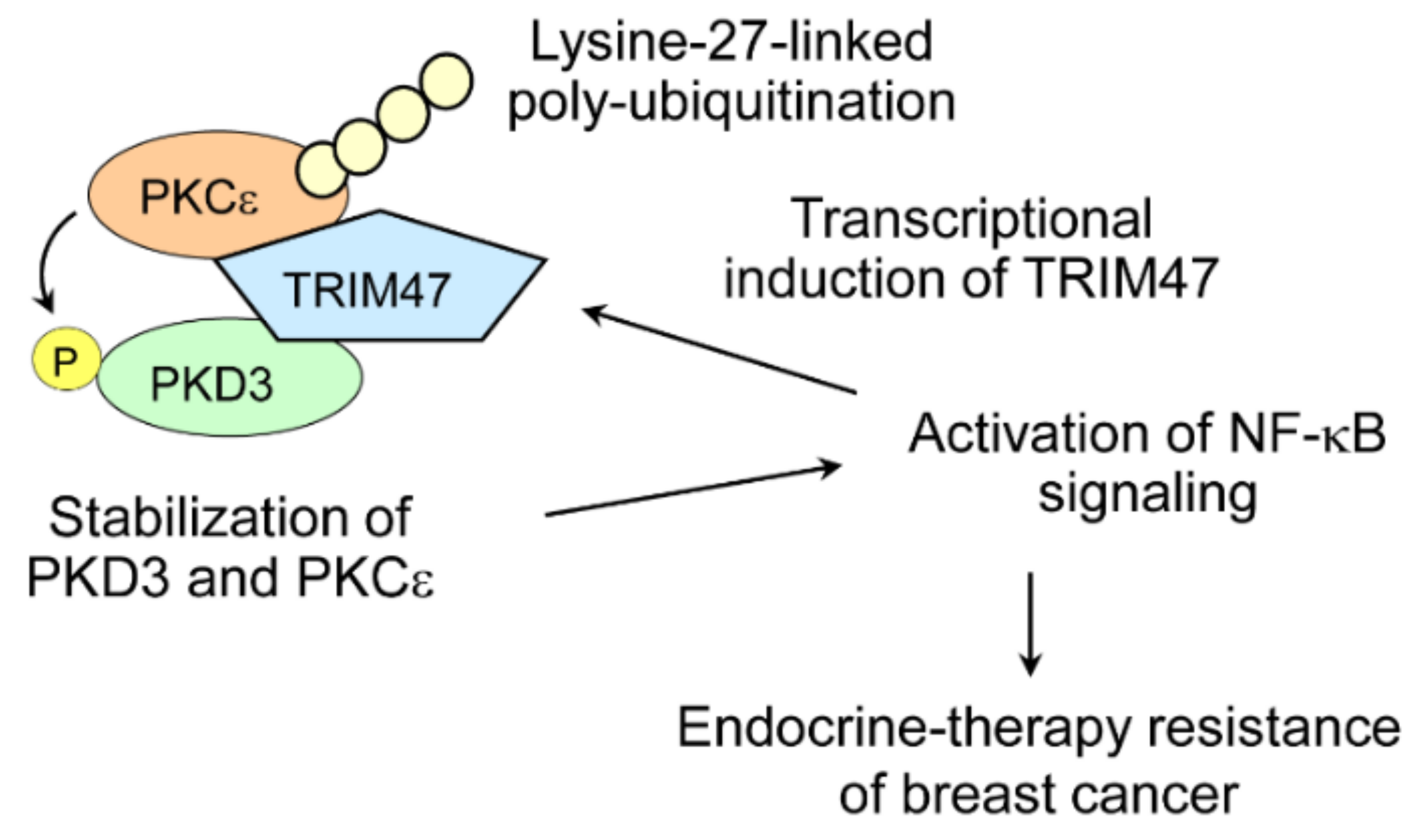
8. TRIM47 and Hormone-Dependent Cancers
9. TRIM47 and Innate Immunity
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Inoue, S.; Orimo, A.; Hosoi, T.; Kondo, S.; Toyoshima, H.; Kondo, T.; Ikegami, A.; Ouchi, Y.; Orimo, H.; Muramatsu, M. Genomic binding-site cloning reveals an estrogen-responsive gene that encodes a RING finger protein. Proc. Natl. Acad. Sci. USA 1993, 90, 11117–11121. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, S. TRIM Family Proteins: Roles in Autophagy, Immunity, and Carcinogenesis. Trends Biochem. Sci. 2017, 42, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Urano, T.; Saito, T.; Tsukui, T.; Fujita, M.; Hosoi, T.; Muramatsu, M.; Ouchi, Y.; Inoue, S. Efp targets 14-3-3 sigma for proteolysis and promotes breast tumour growth. Nature 2002, 417, 871–875. [Google Scholar] [CrossRef]
- Kato, K.; Ahmad, S.; Zhu, Z.; Young, J.M.; Mu, X.; Park, S.; Malik, H.S.; Hur, S. Structural analysis of RIG-I-like receptors reveals ancient rules of engagement between diverse RNA helicases and TRIM ubiquitin ligases. Mol. Cell 2021, 81, 599–613.e8. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Ikeda, K.; Suzuki, T.; Aogi, K.; Horie-Inoue, K.; Inoue, S. TRIM47 activates NF-kappaB signaling via PKC-epsilon/PKD3 stabilization and contributes to endocrine therapy resistance in breast cancer. Proc. Natl. Acad Sci. USA 2021, 118, e2100784118. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. JAMA 2019, 321, 288–300. [Google Scholar] [CrossRef]
- Azuma, K.; Inoue, S. Genomic and non-genomic actions of estrogen: Recent developments. Biomol. Concepts 2012, 3, 365–370. [Google Scholar] [CrossRef]
- Reid, G.; Hübner, M.R.; Métivier, R.; Brand, H.; Denger, S.; Manu, D.; Beaudouin, J.; Ellenberg, J.; Gannon, F. Cyclic, proteasome-mediated turnover of unliganded and liganded ERalpha on responsive promoters is an integral feature of estrogen signaling. Mol. Cell 2003, 11, 695–707. [Google Scholar] [CrossRef]
- Carroll, J.S.; Meyer, C.A.; Song, J.; Li, W.; Geistlinger, T.R.; Eeckhoute, J.; Brodsky, A.S.; Keeton, E.K.; Fertuck, K.C.; Hall, G.F.; et al. Genome-wide analysis of estrogen receptor binding sites. Nat. Genet. 2006, 38, 1289–1297. [Google Scholar] [CrossRef]
- Hah, N.; Danko, C.G.; Core, L.; Waterfall, J.J.; Siepel, A.; Lis, J.T.; Kraus, W.L. A rapid, extensive, and transient transcriptional response to estrogen signaling in breast cancer cells. Cell 2011, 145, 622–634. [Google Scholar] [CrossRef] [Green Version]
- Altucci, L.; Addeo, R.; Cicatiello, L.; Dauvois, S.; Parker, M.G.; Truss, M.; Beato, M.; Sica, V.; Bresciani, F.; Weisz, A. 17beta-estradiol induces cyclin D1 gene transcription, p36D1-p34cdk4 complex activation and p105Rb phosphorylation during mitogenic stimulation of G(1)-arrested human breast cancer cells. Oncogene 1996, 12, 2315–2324. [Google Scholar]
- Cheng, M.; Michalski, S.; Kommagani, R. Role for Growth Regulation by Estrogen in Breast Cancer 1 (GREB1) in Hormone-Dependent Cancers. Int. J. Mol. Sci. 2018, 19, 2543. [Google Scholar] [CrossRef] [Green Version]
- Bhan, A.; Hussain, I.; Ansari, K.I.; Kasiri, S.; Bashyal, A.; Mandal, S.S. Antisense transcript long noncoding RNA (lncRNA) HOTAIR is transcriptionally induced by estradiol. J. Mol. Biol. 2013, 425, 3707–3722. [Google Scholar] [CrossRef] [Green Version]
- Migliaccio, A.; Piccolo, D.; Castoria, G.; Di Domenico, M.; Bilancio, A.; Lombardi, M.; Gong, W.; Beato, M.; Auricchio, F. Activation of the Src/p21ras/Erk pathway by progesterone receptor via cross-talk with estrogen receptor. EMBO J. 1998, 17, 2008–2018. [Google Scholar] [CrossRef] [Green Version]
- Azuma, K.; Urano, T.; Horie-Inoue, K.; Hayashi, S.; Sakai, R.; Ouchi, Y.; Inoue, S. Association of estrogen receptor alpha and histone deacetylase 6 causes rapid deacetylation of tubulin in breast cancer cells. Cancer Res. 2009, 69, 2935–2940. [Google Scholar] [CrossRef] [Green Version]
- Orimo, A.; Inoue, S.; Ikeda, K.; Noji, S.; Muramatsu, M. Molecular cloning, structure, and expression of mouse estrogen-responsive finger protein Efp. Co-localization with estrogen receptor mRNA in target organs. J. Biol. Chem. 1995, 270, 24406–24413. [Google Scholar] [CrossRef] [Green Version]
- Grice, G.L.; Nathan, J.A. The recognition of ubiquitinated proteins by the proteasome. Cell. Mol. Life Sci. 2016, 73, 3497–3506. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Urano, T.; Tsukui, T.; Horie-Inoue, K.; Moriya, T.; Ishida, T.; Muramatsu, M.; Ouchi, Y.; Sasano, H.; Inoue, S. Estrogen-responsive finger protein as a new potential biomarker for breast cancer. Clin. Cancer Res. 2005, 11, 6148–6154. [Google Scholar] [CrossRef] [Green Version]
- Ueyama, K.; Ikeda, K.; Sato, W.; Nakasato, N.; Horie-Inoue, K.; Takeda, S.; Inoue, S. Knockdown of Efp by DNA-modified small interfering RNA inhibits breast cancer cell proliferation and in vivo tumor growth. Cancer Gene Ther. 2010, 17, 624–632. [Google Scholar] [CrossRef]
- Gack, M.U.; Shin, Y.C.; Joo, C.H.; Urano, T.; Liang, C.; Sun, L.; Takeuchi, O.; Akira, S.; Chen, Z.; Inoue, S.; et al. TRIM25 RING-finger E3 ubiquitin ligase is essential for RIG-I-mediated antiviral activity. Nature 2007, 446, 916–920. [Google Scholar] [CrossRef]
- Liu, H.M.; Loo, Y.M.; Horner, S.M.; Zornetzer, G.A.; Katze, M.G.; Gale, M., Jr. The mitochondrial targeting chaperone 14-3-3epsilon regulates a RIG-I translocon that mediates membrane association and innate antiviral immunity. Cell Host Microbe 2012, 11, 528–537. [Google Scholar] [CrossRef] [Green Version]
- Sato, J.; Azuma, K.; Kinowaki, K.; Ikeda, K.; Ogura, T.; Takazawa, Y.; Kawabata, H.; Kitagawa, M.; Inoue, S. Combined Use of Immunoreactivities of RIG-I with Efp/TRIM25 for Predicting Prognosis of Patients with Estrogen Receptor-positive Breast Cancer. Clin. Breast Cancer 2021, 21, 399–407.e2. [Google Scholar] [CrossRef]
- Nakajima, A.; Maruyama, S.; Bohgaki, M.; Miyajima, N.; Tsukiyama, T.; Sakuragi, N.; Hatakeyama, S. Ligand-dependent transcription of estrogen receptor alpha is mediated by the ubiquitin ligase EFP. Biochem. Biophys. Res. Commun. 2007, 357, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.W.; Sikriwal, D.; Dong, X.; Guo, P.; Sun, X.; Dong, J.T. Oestrogen causes degradation of KLF5 by inducing the E3 ubiquitin ligase EFP in ER-positive breast cancer cells. Biochem. J. 2011, 437, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.Y.; Fu, X.; Fan, S.; Guo, P.; Su, D.; Dong, J.T. Oestrogen causes ATBF1 protein degradation through the oestrogen-responsive E3 ubiquitin ligase EFP. Biochem. J. 2012, 444, 581–590. [Google Scholar] [CrossRef] [Green Version]
- Guo, P.; Dong, X.Y.; Zhao, K.W.; Sun, X.; Li, Q.; Dong, J.T. Estrogen-induced interaction between KLF5 and estrogen receptor (ER) suppresses the function of ER in ER-positive breast cancer cells. Int. J. Cancer. 2010, 126, 81–89. [Google Scholar] [CrossRef]
- Dong, X.Y.; Sun, X.; Guo, P.; Li, Q.; Sasahara, M.; Ishii, Y.; Dong, J.T. ATBF1 inhibits estrogen receptor (ER) function by selectively competing with AIB1 for binding to the ER in ER-positive breast cancer cells. J. Biol. Chem. 2010, 285, 32801–32809. [Google Scholar] [CrossRef] [Green Version]
- Sato, W.; Ikeda, K.; Urano, T.; Abe, Y.; Nakasato, N.; Horie-Inoue, K.; Takeda, S.; Inoue, S. Efp promotes in vitro and in vivo growth of endometrial cancer cells along with the activation of nuclear factor-κB signaling. PLoS ONE 2018, 13, e0208351. [Google Scholar] [CrossRef] [Green Version]
- Shiba, S.; Ikeda, K.; Suzuki, T.; Shintani, D.; Okamoto, K.; Horie-Inoue, K.; Hasegawa, K.; Inoue, S. Hormonal Regulation of Patient-Derived Endometrial Cancer Stem-like Cells Generated by Three-Dimensional Culture. Endocrinology 2019, 160, 1895–1906. [Google Scholar] [CrossRef]
- Yang, C.; Ikeda, K.; Horie-Inoue, K.; Sato, W.; Hasegawa, K.; Takeda, S.; Itakura, A.; Inoue, S. Transcriptomic analysis of hormone-sensitive patient-derived endometrial cancer spheroid culture defines Efp as a proliferation modulator. Biochem. Biophys. Res. Commun. 2021, 548, 204–210. [Google Scholar] [CrossRef]
- Nevedomskaya, E.; Baumgart, S.J.; Haendler, B. Recent Advances in Prostate Cancer Treatment and Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashikari, D.; Takayama, K.; Tanaka, T.; Suzuki, Y.; Obinata, D.; Fujimura, T.; Urano, T.; Takahashi, S.; Inoue, S. Androgen induces G3BP2 and SUMO-mediated p53 nuclear export in prostate cancer. Oncogene 2017, 36, 6272–6281. [Google Scholar] [CrossRef] [PubMed]
- Takayama, K.I.; Suzuki, T.; Tanaka, T.; Fujimura, T.; Takahashi, S.; Urano, T.; Ikeda, K.; Inoue, S. TRIM25 enhances cell growth and cell survival by modulating p53 signals via interaction with G3BP2 in prostate cancer. Oncogene 2018, 37, 2165–2180. [Google Scholar] [CrossRef]
- Tomlins, S.A.; Rhodes, D.R.; Perner, S.; Dhanasekaran, S.M.; Mehra, R.; Sun, X.W.; Varambally, S.; Cao, X.; Tchinda, J.; Kuefer, R.; et al. Recurrent fusion of TMPRSS2 and ETS transcription factor genes in prostate cancer. Science 2005, 310, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Kollipara, R.K.; Humphries, C.G.; Ma, S.H.; Hutchinson, R.; Li, R.; Siddiqui, J.; Tomlins, S.A.; Raj, G.V.; Kittler, R. The ubiquitin ligase TRIM25 targets ERG for degradation in prostate cancer. Oncotarget 2016, 7, 64921–64931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zang, H.L.; Ren, S.N.; Cao, H.; Tian, X.F. The ubiquitin ligase TRIM25 inhibits hepatocellular carcinoma progression by targeting metastasis associated 1 protein. IUBMB Life 2017, 69, 795–801. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Li, X.; Cui, K.; Liu, C.; Wu, M.; Prochownik, E.V.; Li, Y. The MAP3K13-TRIM25-FBXW7alpha axis affects c-Myc protein stability and tumor development. Cell Death Differ. 2020, 7, 420–433. [Google Scholar] [CrossRef]
- Liu, Y.; Tao, S.; Liao, L.; Li, Y.; Li, H.; Li, Z.; Lin, L.; Wan, X.; Yang, X.; Chen, L. TRIM25 promotes the cell survival and growth of hepatocellular carcinoma through targeting Keap1-Nrf2 pathway. Nat. Commun. 2020, 11, 348. [Google Scholar] [CrossRef]
- Chen, J.J.; Ren, Y.L.; Shu, C.J.; Zhang, Y.; Chen, M.J.; Xu, J.; Li, J.; Li, A.P.; Chen, D.Y.; He, J.D.; et al. JP3, an antiangiogenic peptide, inhibits growth and metastasis of gastric cancer through TRIM25/SP1/MMP2 axis. J. Exp. Clin. Cancer Res. 2020, 39, 118. [Google Scholar] [CrossRef]
- Yuan, J.; Song, Y.; Pan, W.; Li, Y.; Xu, Y.; Xie, M.; Shen, Y.; Zhang, N.; Liu, J.; Hua, H.; et al. LncRNA SLC26A4-AS1 suppresses the MRN complex-mediated DNA repair signaling and thyroid cancer metastasis by destabilizing DDX5. Oncogene 2020, 39, 6664–6676. [Google Scholar] [CrossRef]
- Wong, D.; Sogerer, L.; Lee, S.S.; Wong, V.; Lum, A.; Levine, A.B.; Marra, M.A.; Yip, S. TRIM25 promotes Capicua degradation independently of ERK in the absence of ATXN1L. BMC Biol. 2020, 18, 154. [Google Scholar] [CrossRef]
- Dissanayake, K.; Toth, R.; Blakey, J.; Olsson, O.; Campbell, D.G.; Prescott, A.R.; MacKintosh, C. ERK/p90(RSK)/14-3-3 signalling has an impact on expression of PEA3 Ets transcription factors via the transcriptional repressor capicua. Biochem. J. 2011, 433, 515–525. [Google Scholar] [CrossRef] [Green Version]
- He, Y.M.; Zhou, X.M.; Jiang, S.Y.; Zhang, Z.B.; Cao, B.Y.; Liu, J.B.; Zeng, Y.Y.; Zhao, J.; Mao, X.L. TRIM25 activates AKT/mTOR by inhibiting PTEN via K63-linked polyubiquitination in non-small cell lung cancer. Acta Pharmacol. Sin. 2022, 43, 681–691. [Google Scholar] [CrossRef]
- Nasrullah, U.; Haeussler, K.; Biyanee, A.; Wittig, I.; Pfeilschifter, J.; Eberhardt, W. Identification of TRIM25 as a Negative Regulator of Caspase-2 Expression Reveals a Novel Target for Sensitizing Colon Carcinoma Cells to Intrinsic Apoptosis. Cells 2019, 8, 1622. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Sun, Q.; Liu, K.; Zhang, L.; Lin, N.; You, K.; Liu, M.; Kon, N.; Tian, F.; Mao, Z.; et al. OTUD5 cooperates with TRIM25 in transcriptional regulation and tumor progression via deubiquitination activity. Nat. Commun. 2020, 11, 4184. [Google Scholar] [CrossRef]
- Li, M.M.; Lau, Z.; Cheung, P.; Aguilar, E.G.; Schneider, W.M.; Bozzacco, L.; Molina, H.; Buehler, E.; Takaoka, A.; Rice, C.M.; et al. TRIM25 Enhances the Antiviral Action of Zinc-Finger Antiviral Protein (ZAP). PLoS Pathog. 2017, 13, e1006145. [Google Scholar] [CrossRef]
- Zheng, X.; Wang, X.; Tu, F.; Wang, Q.; Fan, Z.; Gao, G. TRIM25 Is Required for the Antiviral Activity of Zinc Finger Antiviral Protein. J. Virol. 2017, 91, e00088-e17. [Google Scholar] [CrossRef] [Green Version]
- Brisse, M.; Ly, H. Comparative Structure and Function Analysis of the RIG-I-Like Receptors: RIG-I and MDA5. Front. Immunol. 2019, 10, 1586. [Google Scholar] [CrossRef]
- Oshiumi, H.; Matsumoto, M.; Hatakeyama, S.; Seya, T. Riplet/RNF135, a RING finger protein, ubiquitinates RIG-I to promote interferon-beta induction during the early phase of viral infection. J. Biol. Chem. 2009, 284, 807–817. [Google Scholar] [CrossRef] [Green Version]
- Kuniyoshi, K.; Takeuchi, O.; Pandey, S.; Satoh, T.; Iwasaki, H.; Akira, S.; Kawai, T. Pivotal role of RNA-binding E3 ubiquitin ligase MEX3C in RIG-I-mediated antiviral innate immunity. Proc. Natl. Acad. Sci. USA 2014, 111, 5646–5651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Li, Q.; Mao, A.P.; Hu, M.M.; Shu, H.B. TRIM4 modulates type I interferon induction and cellular antiviral response by targeting RIG-I for K63-linked ubiquitination. J. Mol. Cell. Biol. 2014, 6, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Kouwaki, T.; Fukushima, Y.; Oshiumi, H. Regulation of RIG-I Activation by K63-Linked Polyubiquitination. Front. Immunol. 2018, 8, 1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.; Jiang, M.; Liu, L.; Yang, Z.; Ma, Z.; Liu, S.; Ma, Y.; Zhang, L.; Cao, X. The long noncoding RNA Lnczc3h7a promotes a TRIM25-mediated RIG-I antiviral innate immune response. Nat. Immunol. 2019, 20, 812–823. [Google Scholar] [CrossRef]
- Choudhury, N.R.; Heikel, G.; Trubitsyna, M.; Kubik, P.; Nowak, J.S.; Webb, S.; Granneman, S.; Spanos, C.; Rappsilber, J.; Castello, A.; et al. RNA-binding activity of TRIM25 is mediated by its PRY/SPRY domain and is required for ubiquitination. BMC Biol. 2017, 15, 105. [Google Scholar] [CrossRef] [Green Version]
- Gori Savellini, G.; Anichini, G.; Gandolfo, C.; Cusi, M.G. SARS-CoV-2 N Protein Targets TRIM25-Mediated RIG-I Activation to Suppress Innate Immunity. Viruses 2021, 13, 1439. [Google Scholar] [CrossRef]
- Vandeputte, D.A.; Meije, C.B.; van Dartel, M.; Leenstra, S.; Ijlst-Keizers, H.; Das, P.K.; Troost, D.; Bosch, D.A.; Baas, F.; Hulsebos, T.J. GOA, a novel gene encoding a ring finger B-box coiled-coil protein, is overexpressed in astrocytoma. Biochem. Biophys. Res. Commun. 2001, 286, 574–579. [Google Scholar] [CrossRef]
- Freudenberger, P.; Schmidt, R.; Schmidt, H. Genetics of age-related white matter lesions from linkage to genome wide association studies. J. Neurol. Sci. 2012, 322, 82–86. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.X.; Huang, Z.; Yang, X.; Wang, X.; Zhao, L.P.; Wang, P.X.; Zhang, X.J.; Alves-Bezerra, M.; Cai, L.; Zhang, P.; et al. The deubiquitinating enzyme cylindromatosis mitigates nonalcoholic steatohepatitis. Nat. Med. 2018, 24, 213–223. [Google Scholar] [CrossRef]
- Qian, Y.; Wang, Z.; Lin, H.; Lei, T.; Zhou, Z.; Huang, W.; Wu, X.; Zuo, L.; Wu, J.; Liu, Y.; et al. TRIM47 is a novel endothelial activation factor that aggravates lipopolysaccharide-induced acute lung injury in mice via K63-linked ubiquitination of TRAF2. Signal. Transduct. Target. Ther. 2022, 7, 148. [Google Scholar] [CrossRef]
- Liang, Q.; Tang, C.; Tang, M.; Zhang, Q.; Gao, Y.; Ge, Z. TRIM47 is up-regulated in colorectal cancer, promoting ubiquitination and degradation of SMAD4. J. Exp. Clin. Cancer Res. 2019, 38, 159. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Ding, C.; Zhuang, H.; Hu, W. TRIM47 Promotes the Development of Glioma by Ubiquitination and Degradation of FOXO1. Onco. Targets Ther. 2020, 13, 13401–13411. [Google Scholar] [CrossRef]
- Li, L.; Yu, Y.; Zhang, Z.; Guo, Y.; Yin, T.; Wu, H.; Yang, M. TRIM47 accelerates aerobic glycolysis and tumor progression through regulating ubiquitination of FBP1 in pancreatic cancer. Pharmacol. Res. 2021, 166, 105429. [Google Scholar] [CrossRef]
- Chen, J.X.; Xu, D.; Cao, J.W.; Zuo, L.; Han, Z.T.; Tian, Y.J.; Chu, C.M.; Zhou, W.; Pan, X.W.; Cui, X.G. TRIM47 promotes malignant progression of renal cell carcinoma by degrading P53 through ubiquitination. Cancer Cell Int. 2021, 21, 129. [Google Scholar] [CrossRef]
- Fujimura, T.; Inoue, S.; Urano, T.; Takayama, K.; Yamada, Y.; Ikeda, K.; Obinata, D.; Ashikari, D.; Takahashi, S.; Homma, Y. Increased Expression of Tripartite Motif (TRIM) 47 Is a Negative Prognostic Predictor in Human Prostate Cancer. Clin. Genitourin Cancer 2016, 14, 298–303. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, C.; Xie, Z.; Lu, H. Knockdown of TRIM47 inhibits breast cancer tumorigenesis and progression through the inactivation of PI3K/Akt pathway. Chem. Biol. Interact. 2020, 317, 108960. [Google Scholar] [CrossRef]
- Zhou, Y.; Eppenberger-Castori, S.; Marx, C.; Yau, C.; Scott, G.K.; Eppenberger, U.; Benz, C.C. Activation of nuclear factor-kappaB (NFkappaB) identifies a high-risk subset of hormone-dependent breast cancers. Int. J. Biochem. Cell. Biol. 2005, 37, 1130–1144. [Google Scholar] [CrossRef]
- Frasor, J.; Weaver, A.; Pradhan, M.; Dai, Y.; Miller, L.D.; Lin, C.Y.; Stanculescu, A. Positive cross-talk between estrogen receptor and NF-kappaB in breast cancer. Cancer Res. 2009, 69, 8918–8925. [Google Scholar] [CrossRef] [Green Version]
- Garg, R.; Blando, J.; Perez, C.J.; Wang, H.; Benavides, F.J.; Kazanietz, M.G. Activation of nuclear factor kappaB (NF-kappaB) in prostate cancer is mediated by protein kinase C epsilon (PKCepsilon). J. Biol. Chem. 2012, 287, 37570–37582. [Google Scholar] [CrossRef] [Green Version]
- Zou, Z.; Zeng, F.; Xu, W.; Wang, C.; Ke, Z.; Wang, Q.J.; Deng, F. PKD2 and PKD3 promote prostate cancer cell invasion by modulating NF-κB- and HDAC1-mediated expression and activation of uPA. J. Cell. Sci. 2012, 125, 4800–4811. [Google Scholar]
- Chen, J.; Deng, F.; Singh, S.V.; Wang, Q.J. Protein kinase D3 (PKD3) contributes to prostate cancer cell growth and survival through a PKCepsilon/PKD3 pathway downstream of Akt and ERK 1/2. Cancer Res. 2008, 68, 3844–3853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, S.; Li, G.; Li, G.; Hou, C.; Zheng, Y.; Tang, L.; Gao, Y.; Mo, R.; Li, Y.; Wang, R.; et al. Ubiquitination and degradation of NF90 by Tim-3 inhibits antiviral innate immunity. elife 2021, 10, e66501. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate Proteins | Poly-Ubiquitination Code | Cancer | Function | References |
|---|---|---|---|---|
| 14-3-3σ | unknown | Breast Cancer | degradation | [3] |
| RIG-I | Lys-63-linked | Breast Cancer 1 | activation | [21,23] |
| ERα | Lys-48-linked | Breast Cancer | degradation/ activation | [24] |
| KLF5 | unknown | Breast Cancer | degradation | [25] |
| ATBF-1 | unknown | Breast Cancer | degradation | [26] |
| ERG | Lys-48, Lys-63-linked | Prostate Cancer | degradation | [36] |
| MTA-1 | unknown | Hepatocellular carcinoma | degradation | [37] |
| FBXW7α | Lys-48-linked | Hepatocellular carcinoma | degradation | [38] |
| Keap1 | unknown | Hepatocellular carcinoma | degradation | [39] |
| Sp1 | unknown | Gastric cancer | degradation | [40] |
| DDX5 | unknown | Thyroid cancer | degradation | [41] |
| Capicua | unknown | Brain tumors | degradation | [42] |
| PTEN | Lys-63-linked | Non-small cell Lung carcinoma | prevention of translocation | [44] |
| ZAP | Lys-48, Lys-63-linked | N/A 2 | enhancing activity 3 | [47,48] |
| Substrate Proteins | Poly-Ubiquitination Code | Cancer | Function | References |
|---|---|---|---|---|
| CYLD | unknown | N/A 1 | degradation | [59] |
| TRAF2 | Lys-63-linked | N/A 2 | activation | [60] |
| SMAD4 | unknown | Colorectal cancer | degradation | [61] |
| FOXO1 | unknown | Glioma | degradation | [62] |
| FBP1 | unknown | Pancreatic cancer | degradation | [63] |
| TP53 | unknown | Renal cell carcinoma | degradation | [64] |
| PKCε | Lys-27-linked | Breast cancer | stabilization | [5] |
| NF90 | Lys-48-linked | N/A 3 | degradation | [72] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azuma, K.; Inoue, S. Efp/TRIM25 and Its Related Protein, TRIM47, in Hormone-Dependent Cancers. Cells 2022, 11, 2464. https://doi.org/10.3390/cells11152464
Azuma K, Inoue S. Efp/TRIM25 and Its Related Protein, TRIM47, in Hormone-Dependent Cancers. Cells. 2022; 11(15):2464. https://doi.org/10.3390/cells11152464
Chicago/Turabian StyleAzuma, Kotaro, and Satoshi Inoue. 2022. "Efp/TRIM25 and Its Related Protein, TRIM47, in Hormone-Dependent Cancers" Cells 11, no. 15: 2464. https://doi.org/10.3390/cells11152464