
Therapeutic Potential of Human Immature Dental Pulp Stem Cells Observed in Mouse Model for Acquired Aplastic Anemia

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Aspects
2.2. Cell Culture and Characterization
2.3. Developing Mice Model for AA
2.4. Cell Therapy
2.5. Immunofluorescence (IF) Analyses
2.6. Histological and Immunohistochemistry (IHC-P) Analyses
2.7. Flow Cytometry (FC) Analysis
2.8. BM Colony-Forming Unit Assay (CFU)
2.9. Blood Cell Count
2.10. Statistical Analysis
3. Results
3.1. Immunophenotyping of hIDPSC
3.2. Short- and Long-Term Benefits Observed in BM of an Irradiated Mouse after hIDPSC Transplantation
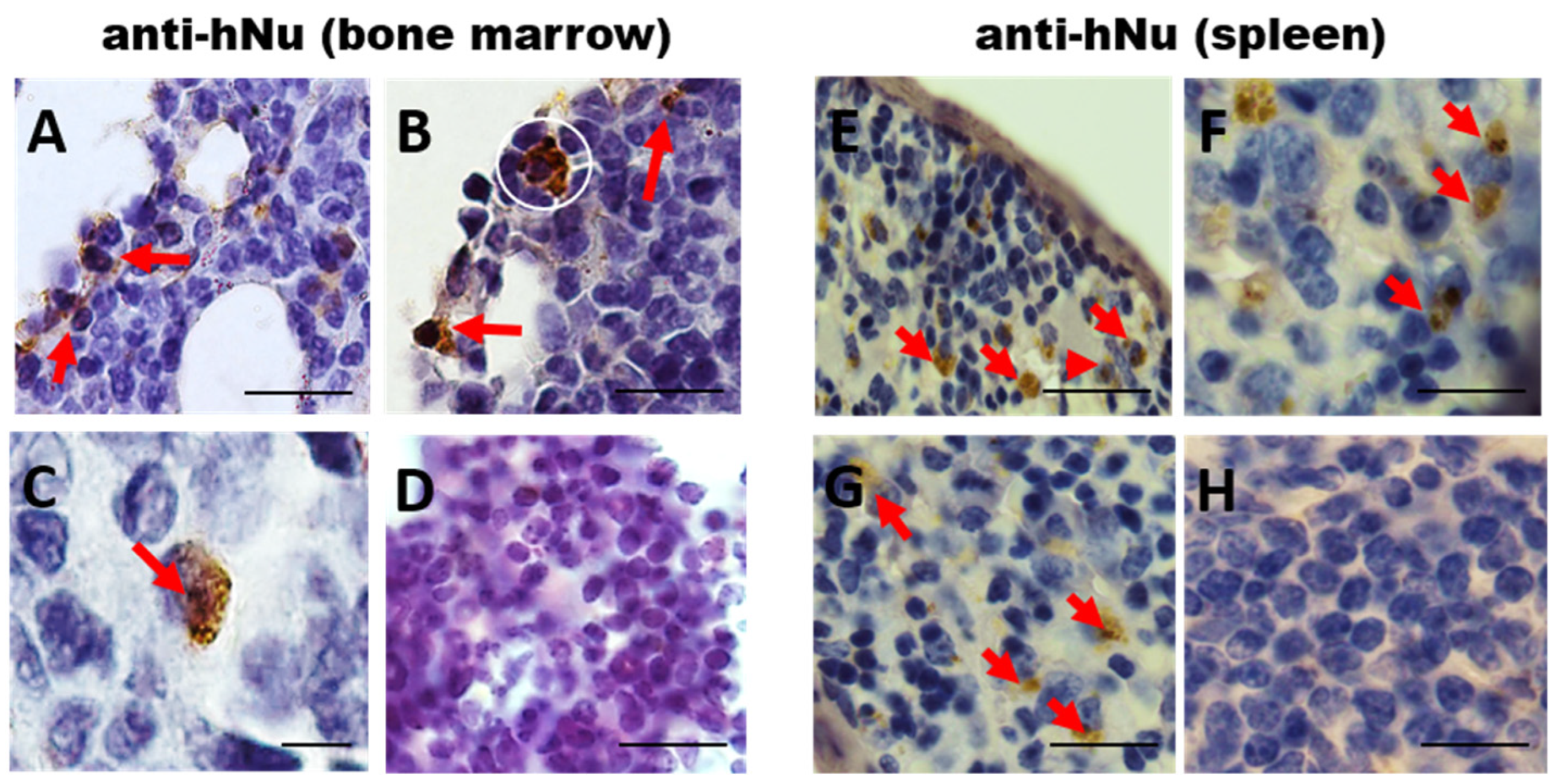
3.3. Long-Term hIDPSC Homing in Irradiated BM
3.4. Evaluation of Expression of Endogenous Stromal and Hematopoietic Cell Markers after hIDPSC Transplantation
3.5. CFU-f Assay
3.6. Monitoring of Hematological Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Shallis, R.M.; Ahmad, R.; Zeidan, A.M. Aplastic anemia: Etiology, molecular pathogenesis, and emerging concepts. Eur. J. Haematol. 2018, 101, 711–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Y.; Katsanis, E. The complex pathophysiology of acquired aplastic anaemia. Clin. Exp. Immunol. 2015, 180, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacigalupo, A.; Benintende, G. Bone marrow transplantation for acquired aplastic anemia: What’s new. Best Pract. Res. Clin. Haematol. 2021, 34, 101284. [Google Scholar] [CrossRef]
- Méndez-Ferrer, S.; Michurina, T.V.; Ferraro, F.; Mazloom, A.R.; MacArthur, B.; Lira, S.A.; Scadden, D.T.; Ma’Ayan, A.; Enikolopov, G.; Frenette, P.S. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 2010, 466, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Greenbaum, A.; Hsu, Y.M.S.; Day, R.B.; Schuettpelz, L.G.; Christopher, M.J.; Borgerding, J.N.; Link, D. C CXCL12 in early mesenchymal progenitors is required for haematopoietic stem-cell mainte-nance. Nature 2013, 495, 227–230. [Google Scholar] [CrossRef] [Green Version]
- Broglie, L.; Margolis, D.; Medin, J.A. Yin and Yang of mesenchymal stem cells and aplastic anemia. World J. Stem Cells 2017, 9, 219–226. [Google Scholar] [CrossRef]
- Man, Y.; Yao, X.; Yang, T.; Wang, Y. Hematopoietic Stem Cell Niche During Homeostasis, Malignancy, and Bone Marrow Transplantation. Front. Cell Dev. Biol. 2021, 9, 621214. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, W.; Ran, Q.; Xiang, Y.; Zhong, J.F.; Li, S.C.; Li, Z. The Differentiation Balance of Bone Marrow Mesenchymal Stem Cells Is Crucial to Hematopoiesis. Stem Cells Int. 2018, 2018, 1540148. [Google Scholar] [CrossRef]
- Pontikoglou, C.; Deschaseaux, F.; Sensebé, L.; Papadaki, H.A. Bone marrow mesenchymal stem cells: Biological properties and their role in hema-topoiesis and hematopoietic stem cell transplantation. Stem Cell Rev. Rep. 2011, 7, 569–589. [Google Scholar] [CrossRef]
- Fattizzo, B.; Giannotta, J.A.; Barcellini, W. Mesenchymal Stem Cells in Aplastic Anemia and Myelodys-plastic Syndromes: The “Seed and Soil” Crosstalk. Int. J. Mol. Sci. 2020, 21, 5438. [Google Scholar] [CrossRef]
- Gonzaga, V.F.; Wenceslau, C.V.; Lisboa, G.S.; Frare, E.O.; Kerkis, I. Mesenchymal Stem Cell Benefits Observed in Bone Marrow Failure and Acquired Aplastic Anemia. Stem Cells Int. 2017, 2017, 8076529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huo, J.; Zhang, L.; Ren, X.; Li, C.; Li, X.; Dong, P.; Zheng, X.; Huang, J.; Shao, Y.; Ge, M.; et al. Multifaceted characterization of the signatures and efficacy of mesenchymal stem/stromal cells in acquired aplastic anemia. Stem Cell Res. Ther. 2020, 11, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, M.B.; Moncivais, K.; Caplan, A. Mesenchymal stem cells: Environmentally responsive therapeutics for regenerative medicine. Exp. Mol. Med. 2013, 45, e54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caplan, A.I. There is no “Stem cell mess”. Tissue Eng. B Rev. 2019, 25, 291–293. [Google Scholar] [CrossRef]
- Caplan, A.I. Adult mesenchymal stem cells: When, where, and how. Stem Cells Int. 2015, 2015, 628767. [Google Scholar] [CrossRef] [Green Version]
- Caplan, A.I. Mesenchymal Stem Cells: Time to Change the Name! Stem Cells Transl. Med. 2017, 6, 1445–1451. [Google Scholar] [CrossRef] [Green Version]
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal stem cell perspective: Cell biology to clinical progress. NPJ Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Tziafas, D.; Smith, A.; Lesot, H. Designing new treatment strategies in vital pulp therapy. J. Dent. 1999, 28, 77–92. [Google Scholar] [CrossRef]
- Dimitrova-Nakov, S.; Baudry, A.; Harichane, Y.; Kellermann, O.; Goldberg, M. Pulp Stem Cells: Implication in Reparative Dentin Formation. J. Endod. 2014, 40, S13–S18. [Google Scholar] [CrossRef]
- Kerkis, I.; Kerkis, A.; Dozortsev, D.; Stukart-Parsons, G.C.; Massironi, S.M.G.; Pereira, L.V.; Caplan, A.I.; Cerruti, H.F. Isolation and Characterization of a Population of Immature Dental Pulp Stem Cells Expressing OCT-4 and Other Embryonic Stem Cell Markers. Cells Tissues Organs 2006, 184, 105–116. [Google Scholar] [CrossRef]
- Kerkis, I.; Caplan, A.I. Stem Cells in Dental Pulp of Deciduous Teeth. Tissue Eng. B Rev. 2012, 18, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Junqueira, L.C.; Carneiro, J. Histologia Básica. In Histologia Básica, 6th ed.; Guanabara Koogan: Rio de Janeiro, Brazil, 1985; 512p. [Google Scholar]
- Vieira, D.P. Avaliação dos Efeitos da Inibição de Cadeias Inflamatórias e da Suplementação Exógena de CXCL12 na Hematopoiese de Modelos Experimentais Expostos a Doses Letais ou Subletais de Radiação Gama. Ph.D. Thesis, Univestity of Sao Paulo, Sao Paulo, Brazil, 2007. [Google Scholar]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-C.; Chang, K.-C.; Tsai, S.-J.; Chang, H.-H.; Lin, C.-P. Neurogenic differentiation of dental pulp stem cells to neuron-like cells in dopaminergic and motor neuronal inductive media. J. Formos. Med. Assoc. 2014, 113, 956–965. [Google Scholar] [CrossRef] [Green Version]
- Itoh, Y.; Sasaki, J.I.; Hashimoto, M.; Katata, C.; Hayashi, M.; Imazato, S. Pulp Regeneration by 3-dimensional Dental Pulp Stem Cell Constructs. J. Dent. Res. 2018, 97, 1137–1143. [Google Scholar] [CrossRef] [PubMed]
- Kerkis, I.; Araldi, R.P.; Wenceslau, C.V.; Mendes, T.B. Advances in Cellular and Cell-Free Therapy Medicinal Products for Huntington Disease Treatment. In From Pathophysiology to Treatment of Huntington’s Disease; Szejko, M.N., Ed.; IntechOpen: London, UK, 2022; Available online: https://www.intechopen.com/online-first/80659 (accessed on 16 May 2022). [CrossRef]
- Araldi, R.P.; D’Amelio, F.; Vigerelli, H.; De Melo, T.C.; Kerkis, I. Stem Cell-Derived Exosomes as Therapeutic Approach for Neurodegenerative Disorders: From Biology to Biotechnology. Cells 2020, 9, 2663. [Google Scholar] [CrossRef] [PubMed]
- Tatullo, M.; Codispoti, B.; Sied, J.; Makeeva, I.; Paduano, F.; Marrelli, M.; Spagnuolo, G. Stem Cells-based and Molecular-based Approaches in Regenerative Dentistry: A Topical Review. Curr. Stem Cell Res. Ther. 2019, 14, 607–616. [Google Scholar] [CrossRef]
- Mortada, I.; Mortada, R.; Al Bazzal, M. Dental Pulp Stem Cells and Neurogenesis. Stem Cells Biol. Eng. 2017, 1083, 63–75. [Google Scholar] [CrossRef]
- da Silva, J.M.; Araldi, R.P.; Colozza-Gama, G.A.; Pagani, E.; Sid, A.; Valverde, C.W.; Kerkis, I. Human Immature Dental Pulp Stem Cells Did Not Graft into a Preexisting Human Lung Adenocarcinoma. Case Rep. Oncol. 2022, 15, 413–422. [Google Scholar] [CrossRef]
- Macedo, J.; Pagani, E.; Wenceslau, C.V.; Ferrara, L.; Kerkis, I. A phase i clinical trial on intravenous administration of immature human dental pulp stem cells (nestacell hdtm) to huntington’s disease patients. Cytotherapy 2021, 23, 4. [Google Scholar] [CrossRef]
- Araldi, R.P.; Prezoto, B.C.; Gonzaga, V.; Policiquio, B.; Mendes, T.B.; D’Amélio, F.; Vigerelli, H.; Viana, M.; Valverde, C.W.; Pagani, E.; et al. Advanced cell therapy with low tissue factor loaded product NestaCell® does not confer thrombogenic risk for critically ill COVID-19 heparin-treated patients. Biomed. Pharmacother. 2022, 149, 112920. [Google Scholar] [CrossRef]
- Lizier, N.F.; Kerkis, A.; Gomes, C.M.; Hebling, J.; Oliveira, C.F.; Caplan, A.I.; Kerkis, I. Scaling-up of dental pulp stem cells isolated from multiple niches. PLoS ONE 2012, 7, e39885. [Google Scholar]
- Che n, J. Animal models for acquired bone marrow failure syndromes. Clin. Med. Res. 2005, 3, 102–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCabe, A.; Smith, J.N.; Costello, A.; Maloney, J.; Katikaneni, D.; MacNamara, K.C. Hematopoietic stem cell loss and hematopoietic failure in severe aplastic anemia is driven by mac-rophages and aberrant podoplanin expression. Haematologica 2018, 103, 1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.T.; Zhao, J.M.; Chu, J.X. Experimental study of low dose irradiation for treatment of immuno-mediated aplastic anemia in mice. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2007, 15, 510–514. [Google Scholar] [PubMed]
- Hu, K.X.; Sun, Q.Y.; Guo, M.; Ai, H.S. The radiation protection and therapy effects of mesenchymal stem cells in mice with acute radiation injury. Br. J. Radiol. 2010, 83, 52–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, S.; Ren, H.; Shi, H.; Liu, W. Allogeneic compact bone-derived mesenchymal stem cell transplantation increases survival of mice ex-posed to lethal total body irradiation: A potential immunological mechanism. Chin. Med. J. 2014, 127, 475–482. [Google Scholar]
- Diaz, M.F.; Horton, P.D.; Dumbali, S.P.; Kumar, A.; Livingston, M.; Skibber, M.A.; Wenzel, P.L. Bone marrow stromal cell therapy improves survival after radiation injury but does not restore endog-enous hematopoiesis. Sci. Rep. 2020, 10, 22211. [Google Scholar] [CrossRef]
- Hérodin, F.; Drouet, M. Cytokine-based treatment of accidentally irradiated victims and new approaches. Exp. Hematol. 2005, 33, 1071–1080. [Google Scholar] [CrossRef]
- López, M.; Martín, M. Medical management of the acute radiation syndrome. Rep. Pract. Oncol. Radiother. 2011, 16, 138–146. [Google Scholar] [CrossRef] [Green Version]
- Kroemeke, A.; Sobczyk-Kruszelnicka, M.; Kwissa-Gajewska, Z. Everyday life following hematopoietic stem cell transplantation: Decline in physical symptoms within the first month and change-related predictors. Qual. Life Res. 2017, 27, 125–135. [Google Scholar] [CrossRef] [Green Version]
- Rios, C.; Jourdain, J.R.; DiCarlo, A.L. Cellular therapies for treatment of radiation injury after a mass casualty incident. Radiat. Res. 2017, 188, 242–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenceslau, C.V.; Miglino, M.A.; Martins, D.S.; Ambrósio, C.E.; Lizier, N.F.; Pignatari, G.C.; Kerkis, I. Mesenchymal Progenitor Cells from Canine Fetal Tissues: Yolk Sac, Liver, and Bone Marrow. Tissue Eng. A 2011, 17, 2165–2176. [Google Scholar] [CrossRef] [PubMed]
- Zoeller, M. CD44, hyaluronan, the hematopoietic stem cell, and leukemiainitiating cells. Front. Immunol. 2015, 6, 235. [Google Scholar]
- Garg, S.; Madkaikar, M.; Ghosh, K. Investigating Cell Surface Markers on Normal Hematopoietic Stem Cells in Three Different Niche Conditions. Int. J. Stem Cells 2013, 6, 129–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirth, F.; Lubosch, A.; Hamelmann, S.; Nakchbandi, I.A. Fibronectin and Its Receptors in Hematopoiesis. Cells 2020, 9, 2717. [Google Scholar] [CrossRef]
- Xie, L.; Zeng, X.; Hu, J.; Chen, Q. Characterization of nestin, a selective marker for bone marrow derived mesenchymal stem cells. Stem Cells Int. 2015, 2015, 762098. [Google Scholar] [CrossRef] [Green Version]
- Beneš, P.; Macečková, V.; Zdráhal, Z.; Konečná, H.; Zahradníčková, E.; Mužík, J.; Šmarda, J. Role of vimentin in regulation of monocyte/macrophage differentiation. Differentiation 2006, 74, 265–276. [Google Scholar] [CrossRef]
- Cao, H.; Heazlewood, S.Y.; Williams, B.; Cardozo, D.; Nigro, J.; Oteiza, A.; Nilsson, S.K. The role of CD44 in fetal and adult hematopoietic stem cell regulation. Haematologica 2015, 101, 26–37. [Google Scholar] [CrossRef] [Green Version]
- Panvini, F.M.; Pacini, S.; Montali, M.; Barachini, S.; Mazzoni, S.; Morganti, R.; Ciancia, E.M.; Carnicelli, V.; Petrini, M. High NESTIN Expression Marks the Endosteal Capillary Network in Human Bone Marrow. Front. Cell Dev. Biol. 2020, 8, 596452. [Google Scholar] [CrossRef]
- Anthony, B.A.; Link, D.C. Regulation of hematopoietic stem cells by bone marrow stromal cells. Trends Immunol. 2013, 35, 32–37. [Google Scholar] [CrossRef] [Green Version]
- Morrison, S.J.; Scadden, D.T. The bone marrow niche for haematopoietic stem cells. Nature 2014, 505, 327–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhou, S.; Zhou, Y.; Feng, F.; Wang, Q.; Zhu, X.; Zhang, X. Adipose-derived mesenchymal stem cells (ADSCs) with the potential to ameliorate platelet recovery, enhance megakaryopoiesis, and inhibit apoptosis of bone marrow cells in a mouse model of radiation-induced thrombocyto-penia. Cell Transplant. 2016, 25, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Gan, J.; Meng, F.; Zhou, X.; Li, C.; He, Y.; Zeng, X.; Li, T. Hematopoietic recovery of acute radiation syndrome by human superoxide dismutase-expressing umbilical cord mes-enchymal stromal cells. Cytotherapy 2015, 17, 403–417. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, H.; Lv, Y.-B.; Wang, Q.; Xie, Z.-J.; Ma, L.-H.; He, J.; Xue, W.; Yu, S.; Guo, J.; et al. Intraperitoneal Injection of Multiplacentas Pooled Cells Treatment on a Mouse Model with Aplastic Anemia. Stem Cells Int. 2016, 2016, 3279793. [Google Scholar] [CrossRef]
- Horwitz, E.M.; Gordon, P.L.; Koo, W.K.; Marx, J.C.; Neel, M.D.; McNall, R.Y.; Hofmann, T. Isolated allogeneic bone marrow-derived mesenchymal cells engraft and stimulate growth in children with osteogenesis imperfecta: Implications for cell therapy of bone. Proc. Natl. Acad. Sci. USA 2002, 99, 8932–8937. [Google Scholar] [CrossRef] [Green Version]
- Jaganathan, B.G.; Tisato, V.; Vulliamy, T.; Dokal, I.; Marsh, J.; Dazzi, F.; Bonnet, D. Effects of MSC co-injection on the reconstitution of aplastic anemia patient following hem-atopoietic stem cell transplantation. Leukemia 2010, 24, 1791–1795. [Google Scholar] [CrossRef]
- Xiao, Y.; Jiang, Z.J.; Pang, Y.; Li, L.; Gao, Y.; Xiao, H.W.; Liu, Q. Efficacy and safety of mesenchymal stromal cell treatment from related donors for patients with re-fractory aplastic anemia. Cytotherapy 2013, 15, 760–766. [Google Scholar] [CrossRef]
- Sackstein, R. The biology of CD44 and HCELL in hematopoiesis: The ‘step 2-bypass pathway’ and other emerging perspectives. Curr. Opin. Hematol. 2011, 18, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Oatley, M.; Bölükbası, V.; Svensson, V.; Shvartsman, M.; Ganter, K.; Zirngibl, K.; Pavlovich, P.V.; Milchevskaya, V.; Foteva, V.; Natarajan, K.N.; et al. Single-cell transcriptomics identifies CD44 as a marker and regulator of endothelial to haematopoietic transition. Nat. Commun. 2020, 11, 586. [Google Scholar] [CrossRef] [Green Version]
- Kim, A.; Shim, S.; Kim, M.-J.; Myung, J.K.; Park, S. Mesenchymal stem cell-mediated Notch2 activation overcomes radiation-induced injury of the hematopoietic system. Sci. Rep. 2018, 8, 9277. [Google Scholar] [CrossRef]
- Williams, N.; Bertoncello, I.; Jackson, H.; Arnold, J.; Kavnoudias, H. The role of interleukin 6 in megakaryocyte formation, megakaryocyte development and platelet production. Ciba Found Symp. 1992, 167, 160–170; discussion 170–173. [Google Scholar] [CrossRef] [PubMed]
- Lotem, J.; Shabo, Y.; Sachs, L. Regulation of Megakaryocyte Development by InterIeukin-6. Blood 1989, 74, 1545–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorronsoro, A.; Lang, V.; Ferrin, I.; Fernández-Rueda, J.; Zabaleta, L.; Pérez-Ruiz, E.; Sepúlveda, P.; Trigueros, C. Intracellular role of IL-6 in mesenchymal stromal cell immunosuppression and proliferation. Sci. Rep. 2020, 10, 21853. [Google Scholar] [CrossRef]
- Yang, S.J.; Wang, X.Q.; Jia, Y.H.; Wang, R.; Cao, K.; Zhang, X.; Tan, Y. Human umbilical cord mesenchymal stem cell transplantation restores hematopoiesis in acute radia-tion disease. Am. J. Transl. Res. 2021, 13, 8670. [Google Scholar] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Species | Brand | Dilution |
|---|---|---|---|
| CD44 | Rat | AbCam, Cambridge, UK (Ab40983) | 1:100 |
| CD90 | Rat | AbCam, Cambridge, UK (Ab3105) | 1:100 |
| Fibronectin | Rabbit | Dako, California, USA (0245) | 1:200 |
| Nestin | Rabbit | AbCam, Cambridge, UK (Ab105389) | 1:200 |
| Human Nucleus | Mouse | AbCam, Cambridge, UK (Ab191181) | 1:500 |
| Vimentin | Goat | Santa Cruz Biotechnology, Texas, USA (sc7557) | 1:100 |
| Antibody | Species | Brand | Dilution |
|---|---|---|---|
| Anti-Goat IgG (H+L) (Alexa Fluor® 647 conjugate) | Goat | Thermo Fischer Scientific, California, USA (A21236) | 1:2000 |
| Anti-Goat IgG (FITC conjugate) | Goat | Santa Cruz Biotechnology, TX, USA (sc2079) | 1:1000 |
| Anti-Goat IgG (HRP conjugate) | Goat | Dako, California, USA (p0449) | 1:100 |
| Anti-Rabbit IgG (H+L) (Alexa Fluor® 633 conjugate) | Rabbit | Thermo Fischer Scientific, California, USA (A21070) | 1:2000 |
| Anti-Rabbit IgG (FITC conjugate) | Rabbit | Santa Cruz Biotechnology, TX, USA (sc2012) | 1:1000 |
| Anti-Mouse/Anti-Rabbit Envision+ Dual Link (HRP conjugate) | Mouse/Rabbit | Dako, California, USA (k4063) | - |
| Anti-Rat IgG (FITC conjugate) | Rat | Santa Cruz Biotechnology, Texas, USA (sc2011) | 1:1000 |
| Anti-Rat IgG (HRP conjugate) | Rat | Dako, California, USA (P0450) | 1:100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzaga, V.F.; Wenceslau, C.V.; Vieira, D.P.; Policiquio, B.d.O.; Khalil, C.; Araldi, R.P.; Kerkis, I. Therapeutic Potential of Human Immature Dental Pulp Stem Cells Observed in Mouse Model for Acquired Aplastic Anemia. Cells 2022, 11, 2252. https://doi.org/10.3390/cells11142252
Gonzaga VF, Wenceslau CV, Vieira DP, Policiquio BdO, Khalil C, Araldi RP, Kerkis I. Therapeutic Potential of Human Immature Dental Pulp Stem Cells Observed in Mouse Model for Acquired Aplastic Anemia. Cells. 2022; 11(14):2252. https://doi.org/10.3390/cells11142252
Chicago/Turabian StyleGonzaga, Vivian Fonseca, Cristiane Valverde Wenceslau, Daniel Perez Vieira, Bruna de Oliveira Policiquio, Charbel Khalil, Rodrigo Pinheiro Araldi, and Irina Kerkis. 2022. "Therapeutic Potential of Human Immature Dental Pulp Stem Cells Observed in Mouse Model for Acquired Aplastic Anemia" Cells 11, no. 14: 2252. https://doi.org/10.3390/cells11142252