
Induced Endothelial Cell-Integrated Liver Assembloids Promote Hepatic Maturation and Therapeutic Effect on Cholestatic Liver Fibrosis

 , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Virus Construction and Production
2.3. Generation of Induced Hepatic Stem Cells (iHepSCs)
2.4. Generation of Induced Endothelial Cells (iECs)
2.5. Co-Culture of iHepSCs and iECs in 2D and Generation of Vascularized Induced Hepatic-Endothelial Cell Assembloids
2.6. In Vitro Functionality Analysis of iHLOs and iHEAs
2.7. Quantitative Real-Time PCR (qRT-PCR)
2.8. Immunocytochemistry
2.9. Western Blotting
2.10. Flow Cytometric Analysis
2.11. DDC-Induced Mouse Model of Cholestatic Liver Fibrosis and Kidney Subcapsular Transplantation of iHLOs or iHEAs
2.12. Histological Analysis
2.13. Porphyrin Measurement from Liver Tissue
2.14. Total Liver Collagen Quantification
2.15. Serum Biochemistry
2.16. Teratoma Formation Assay
2.17. RNA-Sequencing Analysis
2.18. Statistical Analysis
2.19. Data Availability
3. Results
3.1. Generation of Liver Assembloids Consists of Induced Hepatic Stem Cells and Induced Endothelial Cells Generated via Direct Conversion
3.2. Enhanced Functional Maturation of iHepSCs in iHEAs than iHLOs through Interaction with Fibronectin Expressed from iECs
3.3. Global Transcriptional Profile of iHLOs and iHEAs
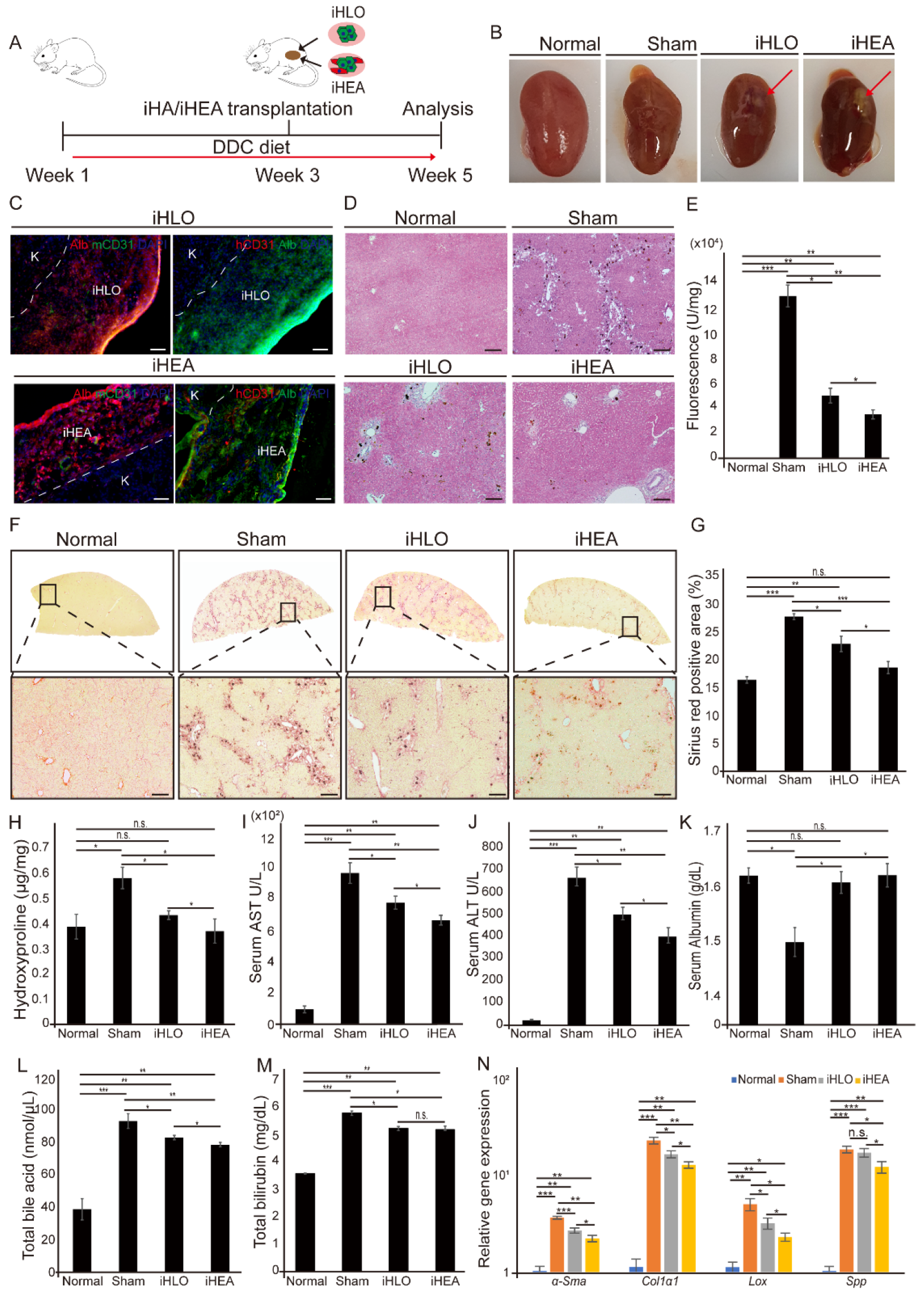
3.4. Transplantation of iHLOs and iHEAs in DDC-Induced Cholestatic Liver Fibrosis Model Ameliorates Liver Fibrosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baptista, P.M.; Penning, L.C. Transplantable Liver Organoids, Too Many Cell Types to Choose: A Need for Scientific Self-Organization. Curr. Transplant. Rep. 2020, 7, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Cordero-Espinoza, L.; Huch, M. The Balancing Act of the Liver: Tissue Regeneration Versus Fibrosis. J. Clin. Investig. 2018, 128, 85–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sepanlou, S.G.; Safiri, S.; Bisignano, C.; Ikuta, K.S.; Merat, S.; Saberifiroozi, M.; Poustchi, H.; Tsoi, D.; Colombara, D.V.; Abdoli, A.; et al. The Global, Regional, and National Burden of Cirrhosis by Cause in 195 Countries and Territories, 1990–2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet Gastroenterol. Hepatol. 2020, 5, 245–266. [Google Scholar] [CrossRef] [Green Version]
- Asrani, S.K.; Devarbhavi, H.; Eaton, J.; Kamath, P.S. Burden of liver diseases in the world. J. Hepatol. 2019, 70, 151–171. [Google Scholar] [CrossRef]
- Kim, J.Y.; An, H.J.; Kim, W.H.; Park, Y.Y.; Park, K.D.; Park, K.K. Apamin suppresses biliary fibrosis and activation of hepatic stellate cells. Int. J. Mol. Med. 2017, 39, 1188–1194. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Tang, L.; Yi, Q. Engineering the Vasculature of Stem-Cell-Derived Liver Organoids. Biomolecules 2021, 11, 966. [Google Scholar] [CrossRef]
- Mavila, N.; Trecartin, A.; Spurrier, R.; Xiao, Y.; Hou, X.; James, D.; Fu, X.; Truong, B.; Wang, C.; Lipshutz, G.S.; et al. Functional Human and Murine Tissue-Engineered Liver Is Generated from Adult Stem/Progenitor Cells. Stem Cells Transl. Med. 2017, 6, 238–248. [Google Scholar] [CrossRef] [Green Version]
- Hughes, R.D.; Mitry, R.R.; Dhawan, A. Current status of hepatocyte transplantation. Transplantation 2012, 93, 342–347. [Google Scholar] [CrossRef]
- Nussler, A.; Konig, S.; Ott, M.; Sokal, E.; Christ, B.; Thasler, W.; Brulport, M.; Gabelein, G.; Schormann, W.; Schulze, M.; et al. Present status and perspectives of cell-based therapies for liver diseases. J. Hepatol. 2006, 45, 144–159. [Google Scholar] [CrossRef]
- Soto-Gutiérrez, A.; Kobayashi, N.; Rivas-Carrillo, J.D.; Navarro-Alvarez, N.; Zhao, D.; Okitsu, T.; Noguchi, H.; Basma, H.; Tabata, Y.; Chen, Y.; et al. Reversal of mouse hepatic failure using an implanted liver-assist device containing ES cell-derived hepatocytes. Nat. Biotechnol. 2006, 24, 1412–1419. [Google Scholar] [CrossRef] [Green Version]
- Woo, D.H.; Kim, S.K.; Lim, H.J.; Heo, J.; Park, H.S.; Kang, G.Y.; Kim, S.E.; You, H.J.; Hoeppner, D.J.; Kim, Y.; et al. Direct and indirect contribution of human embryonic stem cell-derived hepatocyte-like cells to liver repair in mice. Gastroenterology 2012, 142, 602–611. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.M.; Liao, Y.W.; Chiang, C.H.; Chen, Y.J.; Lai, Y.H.; Chang, Y.L.; Chen, H.L.; Jeng, S.Y.; Hsieh, J.H.; Peng, C.H.; et al. Improvement of carbon tetrachloride-induced acute hepatic failure by transplantation of induced pluripotent stem cells without reprogramming factor c-Myc. Int. J. Mol. Sci. 2012, 13, 3598–3617. [Google Scholar] [CrossRef] [PubMed]
- Park, M.R.; Wong, M.S.; Araúzo-Bravo, M.J.; Lee, H.; Nam, D.; Park, S.Y.; Seo, H.D.; Lee, S.M.; Zeilhofer, H.F.; Zaehres, H.; et al. Oct4 and Hnf4α-induced hepatic stem cells ameliorate chronic liver injury in liver fibrosis model. PLoS ONE 2019, 14, e0221085. [Google Scholar] [CrossRef] [Green Version]
- Miura, K.; Okada, Y.; Aoi, T.; Okada, A.; Takahashi, K.; Okita, K.; Nakagawa, M.; Koyanagi, M.; Tanabe, K.; Ohnuki, M.; et al. Variation in the safety of induced pluripotent stem cell lines. Nat. Biotechnol. 2009, 27, 743–745. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Yang, Y.; Liu, J.; Qian, L. Direct cell reprogramming: Approaches, mechanisms and progress. Nat. Rev. Mol. Cell Biol. 2021, 22, 410–424. [Google Scholar] [CrossRef]
- Du, Y.; Wang, J.; Jia, J.; Song, N.; Xiang, C.; Xu, J.; Hou, Z.; Su, X.; Liu, B.; Jiang, T.; et al. Human hepatocytes with drug metabolic function induced from fibroblasts by lineage reprogramming. Cell Stem Cell 2014, 14, 394–403. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.; He, Z.; Ji, S.; Sun, H.; Xiang, D.; Liu, C.; Hu, Y.; Wang, X.; Hui, L. Induction of functional hepatocyte-like cells from mouse fibroblasts by defined factors. Nature 2011, 475, 386–389. [Google Scholar] [CrossRef]
- Lee, C.A.; Sinha, S.; Fitzpatrick, E.; Dhawan, A. Hepatocyte transplantation and advancements in alternative cell sources for liver-based regenerative medicine. J. Mol. Med. 2018, 96, 469–481. [Google Scholar] [CrossRef] [Green Version]
- Hughes, R.D.; Mitry, R.R.; Dhawan, A. Hepatocyte transplantation for metabolic liver disease: UK experience. J. R. Soc. Med. 2005, 98, 341–345. [Google Scholar] [CrossRef]
- Ott, M.; Castell, J.V. Hepatocyte transplantation, a step forward? J. Hepatol. 2019, 70, 1049–1050. [Google Scholar] [CrossRef] [Green Version]
- Takebe, T.; Sekine, K.; Enomura, M.; Koike, H.; Kimura, M.; Ogaeri, T.; Zhang, R.-R.; Ueno, Y.; Zheng, Y.-W.; Koike, N.; et al. Vascularized and functional human liver from an iPSC-derived organ bud transplant. Nature 2013, 499, 481. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.-Z.; Zheng, Y.-W.; Ogawa, M.; Miyagi, E.; Taniguchi, H. Human liver organoids generated with single donor-derived multiple cells rescue mice from acute liver failure. Stem Cell Res. Ther. 2018, 9, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, N. Assembloids. Nat. Methods 2021, 18, 27. [Google Scholar] [CrossRef]
- Wang, L.; Sievert, D.; Clark, A.E.; Lee, S.; Federman, H.; Gastfriend, B.D.; Shusta, E.V.; Palecek, S.P.; Carlin, A.F.; Gleeson, J.G. A human three-dimensional neural-perivascular ‘assembloid’ promotes astrocytic development and enables modeling of SARS-CoV-2 neuropathology. Nat. Med. 2021, 27, 1600–1606. [Google Scholar] [CrossRef]
- Warlich, E.; Kuehle, J.; Cantz, T.; Brugman, M.H.; Maetzig, T.; Galla, M.; Filipczyk, A.A.; Halle, S.; Klump, H.; Schöler, H.R.; et al. Lentiviral vector design and imaging approaches to visualize the early stages of cellular reprogramming. Mol. Ther. 2011, 19, 782–789. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B.; Zaehres, H.; Araúzo-Bravo, M.J.; Schöler, H.R. Generation of induced pluripotent stem cells from neural stem cells. Nat. Protoc. 2009, 4, 1464–1470. [Google Scholar] [CrossRef]
- Morita, R.; Suzuki, M.; Kasahara, H.; Shimizu, N.; Shichita, T.; Sekiya, T.; Kimura, A.; Sasaki, K.-I.; Yasukawa, H.; Yoshimura, A. ETS transcription factor ETV2 directly converts human fibroblasts into functional endothelial cells. Proc. Natl. Acad. Sci. USA 2015, 112, 160–165. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Park, C.; Han, J.W.; Kim, J.Y.; Cho, K.; Kim, E.J.; Kim, S.; Lee, S.J.; Oh, S.Y.; Tanaka, Y.; et al. Direct Reprogramming of Human Dermal Fibroblasts Into Endothelial Cells Using ER71/ETV2. Circ. Res. 2017, 120, 848–861. [Google Scholar] [CrossRef] [Green Version]
- Singhal, N.; Araúzo-Bravo, M.; Sinn, M.; Zaehres, H. Baf Complex Enhances Reprogramming of Adult Human Fibroblasts. J. Stem Cell Res. Ther. 2016, 6, 0336. [Google Scholar] [CrossRef] [Green Version]
- Haridass, D.; Yuan, Q.; Becker, P.D.; Cantz, T.; Iken, M.; Rothe, M.; Narain, N.; Bock, M.; Nörder, M.; Legrand, N.; et al. Repopulation efficiencies of adult hepatocytes, fetal liver progenitor cells, and embryonic stem cell-derived hepatic cells in albumin-promoter-enhancer urokinase-type plasminogen activator mice. Am. J. Pathol. 2009, 175, 1483–1492. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Hillebrand, A.; Wang, D.Q.; Liu, M. Isolation and primary culture of rat hepatic cells. J. Vis. Exp. 2012, 64, e3917. [Google Scholar] [CrossRef] [PubMed]
- Chiba, T.; Seki, A.; Aoki, R.; Ichikawa, H.; Negishi, M.; Miyagi, S.; Oguro, H.; Saraya, A.; Kamiya, A.; Nakauchi, H.; et al. Bmi1 promotes hepatic stem cell expansion and tumorigenicity in both Ink4a/Arf-dependent and -independent manners in mice. Hepatology 2010, 52, 1111–1123. [Google Scholar] [CrossRef]
- Dai, Z.; Song, G.; Balakrishnan, A.; Yang, T.; Yuan, Q.; Möbus, S.; Weiss, A.-C.; Bentler, M.; Zhu, J.; Jiang, X.; et al. Growth differentiation factor 11 attenuates liver fibrosis via expansion of liver progenitor cells. Gut 2020, 69, 1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takebe, T.; Zhang, R.-R.; Koike, H.; Kimura, M.; Yoshizawa, E.; Enomura, M.; Koike, N.; Sekine, K.; Taniguchi, H. Generation of a vascularized and functional human liver from an iPSC-derived organ bud transplant. Nat. Protoc. 2014, 9, 396. [Google Scholar] [CrossRef] [PubMed]
- Saggi, H.; Maitra, D.; Jiang, A.; Zhang, R.; Wang, P.; Cornuet, P.; Singh, S.; Locker, J.; Ma, X.; Dailey, H.; et al. Loss of hepatocyte β-catenin protects mice from experimental porphyria-associated liver injury. J. Hepatol. 2019, 70, 108–117. [Google Scholar] [CrossRef]
- Lee, H.; Lee, H.Y.; Lee, B.E.; Gerovska, D.; Park, S.Y.; Zaehres, H.; Araúzo-Bravo, M.J.; Kim, J.I.; Ha, Y.; Schöler, H.R.; et al. Sequentially induced motor neurons from human fibroblasts facilitate locomotor recovery in a rodent spinal cord injury model. Elife 2020, 9, e52069. [Google Scholar] [CrossRef]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Koivumäki, J.T.; Naumenko, N.; Tuomainen, T.; Takalo, J.; Oksanen, M.; Puttonen, K.A.; Lehtonen, Š.; Kuusisto, J.; Laakso, M.; Koistinaho, J.; et al. Structural Immaturity of Human iPSC-Derived Cardiomyocytes: In Silico Investigation of Effects on Function and Disease Modeling. Front. Physiol. 2018, 9, 80. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, J.; Udono, M.; Miura, S.; Sekiya, S.; Suzuki, A. Cell Aggregation Culture Induces Functional Differentiation of Induced Hepatocyte-like Cells through Activation of Hippo Signaling. Cell Rep. 2018, 25, 183–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Park, M.R.; Choi, C.; Kim, J.B.; Cha, C. Synergistic control of mechanics and microarchitecture of 3D bioactive hydrogel platform to promote the regenerative potential of engineered hepatic tissue. Biomaterials 2021, 270, 120688. [Google Scholar] [CrossRef]
- Seo, W.; Jeong, W.I. Hepatic non-parenchymal cells: Master regulators of alcoholic liver disease? World J. Gastroenterol. 2016, 22, 1348–1356. [Google Scholar] [CrossRef] [PubMed]
- Asai, A.; Aihara, E.; Watson, C.; Mourya, R.; Mizuochi, T.; Shivakumar, P.; Phelan, K.; Mayhew, C.; Helmrath, M.; Takebe, T.; et al. Paracrine signals regulate human liver organoid maturation from induced pluripotent stem cells. Development 2017, 144, 1056–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohno, M.; Motojima, K.; Okano, T.; Taniguchi, A. Maturation of the extracellular matrix and cell adhesion molecules in layered co-cultures of HepG2 and endothelial cells. J. Biochem. 2009, 145, 591–597. [Google Scholar] [CrossRef]
- Ohno, M.; Motojima, K.; Okano, T.; Taniguchi, A. Up-regulation of drug-metabolizing enzyme genes in layered co-culture of a human liver cell line and endothelial cells. Tissue Eng. Part A 2008, 14, 1861–1869. [Google Scholar] [CrossRef]
- Pettinato, G.; Lehoux, S.; Ramanathan, R.; Salem, M.M.; He, L.-X.; Muse, O.; Flaumenhaft, R.; Thompson, M.T.; Rouse, E.A.; Cummings, R.D.; et al. Generation of fully functional hepatocyte-like organoids from human induced pluripotent stem cells mixed with Endothelial Cells. Sci. Rep. 2019, 9, 8920. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, A.; Alvarez, A.M.; Pagan, R.; Roncero, C.; Vilaró, S.; Benito, M.; Fabregat, I. Fibronectin regulates morphology, cell organization and gene expression of rat fetal hepatocytes in primary culture. J. Hepatol. 2000, 32, 242–250. [Google Scholar] [CrossRef]
- Nishida, Y.; Taniguchi, A. Induction of albumin expression in HepG2 cells using immobilized simplified recombinant fibronectin protein. In Vitro Cell Dev. Biol. Anim. 2013, 49, 400–407. [Google Scholar] [CrossRef]
- Wang, Y.; Kim, M.H.; Shirahama, H.; Lee, J.H.; Ng, S.S.; Glenn, J.S.; Cho, N.J. ECM proteins in a microporous scaffold influence hepatocyte morphology, function, and gene expression. Sci. Rep. 2016, 6, 37427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, Y.; You, D.; Kang, H.G.; Yu, J.; Kim, S.W.; Nam, S.J.; Lee, J.E.; Kim, S. Berberine Suppresses Fibronectin Expression through Inhibition of c-Jun Phosphorylation in Breast Cancer Cells. J. Breast Cancer 2018, 21, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Borkham-Kamphorst, E.; Zimmermann, H.W.; Gassler, N.; Bissels, U.; Bosio, A.; Tacke, F.; Weiskirchen, R.; Kanse, S.M. Factor VII activating protease (FSAP) exerts anti-inflammatory and anti-fibrotic effects in liver fibrosis in mice and men. J. Hepatol. 2013, 58, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Palacian, A.; Kanse, S.M.; Weiskirchen, R. Factor VII activating protease (FSAP): A novel protective factor in liver fibrosis. Proteom. Clin. Appl. 2014, 8, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Liedtke, C.; Luedde, T.; Sauerbruch, T.; Scholten, D.; Streetz, K.; Tacke, F.; Tolba, R.; Trautwein, C.; Trebicka, J.; Weiskirchen, R. Experimental liver fibrosis research: Update on animal models, legal issues and translational aspects. Fibrogenesis Tissue Repair 2013, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, G.; Qu, J.; Yuan, Z.; Pan, R.; Li, K. Calcipotriol Inhibits NLRP3 Signal Through YAP1 Activation to Alleviate Cholestatic Liver Injury and Fibrosis. Front. Pharmacol. 2020, 11, 200. [Google Scholar] [CrossRef]
- Pose, E.; Sancho-Bru, P.; Coll, M. 3,5-Diethoxycarbonyl-1,4-Dihydrocollidine Diet: A Rodent Model in Cholestasis Research. Methods Mol. Biol. 2019, 1981, 249–257. [Google Scholar]
- Naganuma, H.; Nishinakamura, R. From organoids to transplantable artificial kidneys. Transpl. Int. 2019, 32, 563–570. [Google Scholar] [CrossRef] [Green Version]
- van den Berg, C.W.; Ritsma, L.; Avramut, M.C.; Wiersma, L.E.; van den Berg, B.M.; Leuning, D.G.; Lievers, E.; Koning, M.; Vanslambrouck, J.M.; Koster, A.J.; et al. Renal Subcapsular Transplantation of PSC-Derived Kidney Organoids Induces Neo-vasculogenesis and Significant Glomerular and Tubular Maturation In Vivo. Stem Cell Rep. 2018, 10, 751–765. [Google Scholar] [CrossRef] [Green Version]
- Greaves, P. Chapter 9—Liver and Pancreas. In Histopathology of Preclinical Toxicity Studies, 4th ed.; Greaves, P., Ed.; Academic Press: Boston, MA, USA, 2012. [Google Scholar]
- Fragoulis, A.; Schenkel, J.; Herzog, M.; Schellenberg, T.; Jahr, H.; Pufe, T.; Trautwein, C.; Kensler, T.W.; Streetz, K.L.; Wruck, C.J. Nrf2 Ameliorates DDC-Induced Sclerosing Cholangitis and Biliary Fibrosis and Improves the Regenerative Capacity of the Liver. Toxicol. Sci. 2019, 169, 485–498. [Google Scholar] [CrossRef]
- Yap, K.K.; Gerrand, Y.-W.; Dingle, A.M.; Yeoh, G.C.; Morrison, W.A.; Mitchell, G.M. Liver sinusoidal endothelial cells promote the differentiation and survival of mouse vascularised hepatobiliary organoids. Biomaterials 2020, 251, 120091. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Boyer, J.L. The maintenance and generation of membrane polarity in hepatocytes. Hepatology 2004, 39, 892–899. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.S.; Nolan, D.J.; Butler, J.M.; James, D.; Babazadeh, A.O.; Rosenwaks, Z.; Mittal, V.; Kobayashi, H.; Shido, K.; Lyden, D.; et al. Inductive angiocrine signals from sinusoidal endothelium are required for liver regeneration. Nature 2010, 468, 310–315. [Google Scholar] [CrossRef] [Green Version]
- Moriya, K.; Sakai, K.; Yan, M.H.; Sakai, T. Fibronectin is essential for survival but is dispensable for proliferation of hepatocytes in acute liver injury in mice. Hepatology 2012, 56, 311–321. [Google Scholar] [CrossRef] [Green Version]
- Jaffe, E.A.; Mosher, D.F. Synthesis of fibronectin by cultured human endothelial cells. Ann. N. Y. Acad. Sci. 1978, 312, 122–131. [Google Scholar] [CrossRef]
- DeLaForest, A.; Nagaoka, M.; Si-Tayeb, K.; Noto, F.K.; Konopka, G.; Battle, M.A.; Duncan, S.A. HNF4A is essential for specification of hepatic progenitors from human pluripotent stem cells. Development 2011, 138, 4143–4153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.H.; Sauer, B.; Gonzalez, F.J. Laron dwarfism and non-insulin-dependent diabetes mellitus in the Hnf-1alpha knockout mouse. Mol. Cell Biol. 1998, 18, 3059–3068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, D.Q.; Bussen, M.; Sehayek, E.; Ananthanarayanan, M.; Shneider, B.L.; Suchy, F.J.; Shefer, S.; Bollileni, J.S.; Gonzalez, F.J.; Breslow, J.L.; et al. Hepatocyte nuclear factor-1alpha is an essential regulator of bile acid and plasma cholesterol metabolism. Nat. Genet. 2001, 27, 375–382. [Google Scholar] [CrossRef]
- Lau, H.H.; Ng, N.H.J.; Loo, L.S.W.; Jasmen, J.B.; Teo, A.K.K. The molecular functions of hepatocyte nuclear factors—In and beyond the liver. J. Hepatol. 2018, 68, 1033–1048. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Vries, R.G.; Snippert, H.J.; van de Wetering, M.; Barker, N.; Stange, D.E.; van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J.; et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef]
- Corrò, C.; Novellasdemunt, L.; Li, V.S.W. A brief history of organoids. Am. J. Physiol. Cell Physiol. 2020, 319, C151–C165. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, G.; Rezakhani, S.; Yildiz, E.; Nuciforo, S.; Heim, M.H.; Lutolf, M.P.; Schoonjans, K. Mechano-modulatory synthetic niches for liver organoid derivation. Nat. Commun. 2020, 11, 3416. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.S.; Tang, C.; Rao, M.S.; Weissman, I.L.; Wu, J.C. Tumorigenicity as a clinical hurdle for pluripotent stem cell therapies. Nat. Med. 2013, 19, 998–1004. [Google Scholar] [CrossRef] [Green Version]
- Martin, R.M.; Fowler, J.L.; Cromer, M.K.; Lesch, B.J.; Ponce, E.; Uchida, N.; Nishimura, T.; Porteus, M.H.; Loh, K.M. Improving the safety of human pluripotent stem cell therapies using genome-edited orthogonal safeguards. Nat. Commun. 2020, 11, 2713. [Google Scholar] [CrossRef]
- Xie, B.; Sun, D.; Du, Y.; Jia, J.; Sun, S.; Xu, J.; Liu, Y.; Xiang, C.; Chen, S.; Xie, H.; et al. A two-step lineage reprogramming strategy to generate functionally competent human hepatocytes from fibroblasts. Cell Res. 2019, 29, 696–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.B.; Lee, H.; Araúzo-Bravo, M.J.; Hwang, K.; Nam, D.; Park, M.R.; Zaehres, H.; Park, K.I.; Lee, S.J. Oct4-induced oligodendrocyte progenitor cells enhance functional recovery in spinal cord injury model. EMBO J. 2015, 34, 2971–2983. [Google Scholar] [CrossRef] [Green Version]
- Quadrato, G.; Nguyen, T.; Macosko, E.Z.; Sherwood, J.L.; Min Yang, S.; Berger, D.R.; Maria, N.; Scholvin, J.; Goldman, M.; Kinney, J.P.; et al. Cell diversity and network dynamics in photosensitive human brain organoids. Nature 2017, 545, 48–53. [Google Scholar] [CrossRef] [Green Version]
- Ormel, P.R.; Vieira de Sá, R.; van Bodegraven, E.J.; Karst, H.; Harschnitz, O.; Sneeboer, M.A.M.; Johansen, L.E.; van Dijk, R.E.; Scheefhals, N.; Berdenis van Berlekom, A.; et al. Microglia innately develop within cerebral organoids. Nat. Commun. 2018, 9, 4167. [Google Scholar] [CrossRef]
- Thompson, W.L.; Takebe, T. Chapter 3—Generation of Multi-Cellular Human Liver Organoids from Pluripotent Stem Cells. In Methods in Cell Biology; Spence, J.R., Ed.; Academic Press: Cambridge, MA, USA, 2020; Volume 159, pp. 47–68. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nam, D.; Park, M.R.; Lee, H.; Bae, S.C.; Gerovska, D.; Araúzo-Bravo, M.J.; Zaehres, H.; Schöler, H.R.; Kim, J.B. Induced Endothelial Cell-Integrated Liver Assembloids Promote Hepatic Maturation and Therapeutic Effect on Cholestatic Liver Fibrosis. Cells 2022, 11, 2242. https://doi.org/10.3390/cells11142242
Nam D, Park MR, Lee H, Bae SC, Gerovska D, Araúzo-Bravo MJ, Zaehres H, Schöler HR, Kim JB. Induced Endothelial Cell-Integrated Liver Assembloids Promote Hepatic Maturation and Therapeutic Effect on Cholestatic Liver Fibrosis. Cells. 2022; 11(14):2242. https://doi.org/10.3390/cells11142242
Chicago/Turabian StyleNam, Donggyu, Myung Rae Park, Hyunah Lee, Sung Chul Bae, Daniela Gerovska, Marcos J. Araúzo-Bravo, Holm Zaehres, Hans R. Schöler, and Jeong Beom Kim. 2022. "Induced Endothelial Cell-Integrated Liver Assembloids Promote Hepatic Maturation and Therapeutic Effect on Cholestatic Liver Fibrosis" Cells 11, no. 14: 2242. https://doi.org/10.3390/cells11142242