
Cyclic Hypoxia Induces Transcriptomic Changes in Mast Cells Leading to a Hyperresponsive Phenotype after FcεRI Cross-Linking



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Reagents and Antibodies
2.3. Culture of B16-F1 Melanoma Cells and Tumor Generation
2.4. Immunofluorescence and Confocal Microscopy in Tumor Biopsies
2.5. Generation of BMMCs Cultures
2.6. In Vitro Protocol for Cyclic Hypoxia
2.7. Viability Test
2.8. RNA Extraction and RT-PCR
2.9. Determination of ROS Production
2.10. Microarray Assay
2.10.1. Total RNA Isolation
2.10.2. Probe Preparation and Hybridization to Arrays
2.10.3. Data Acquisition and Analysis of Array Images
2.10.4. Microarray Data Analysis
2.10.5. Functional Enrichment Analysis
2.11. Quantitative Real-Time PCR
2.12. Degranulation Assay
2.13. Determination of Intracellular Calcium
2.14. Statistical Analysis
3. Results
3.1. MCs Can Be Found in Cyclic Hypoxia Regions within Murine B16 Melanoma Tumors
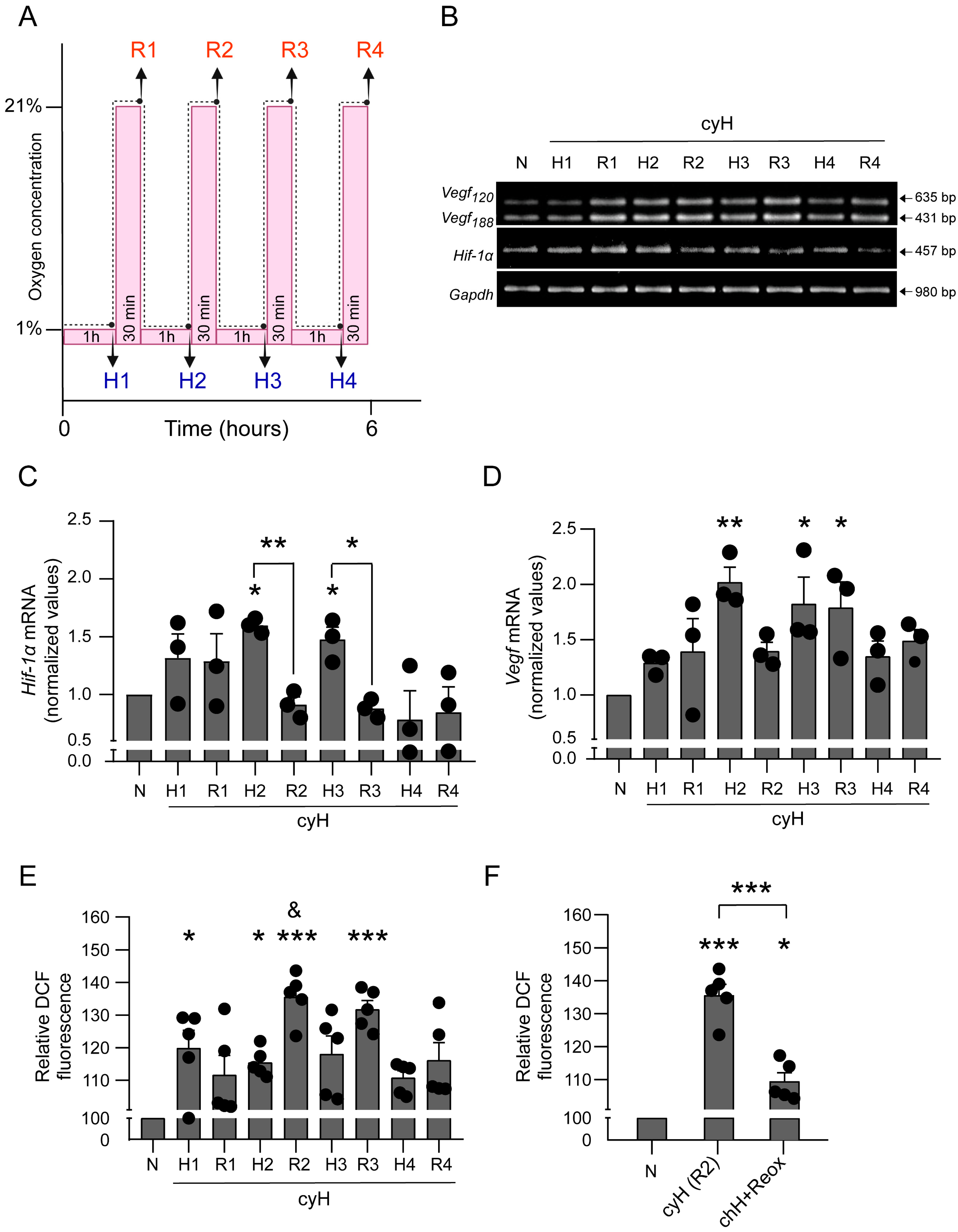
3.2. Cyclic Hypoxia Increases mRNA Levels of Hypoxia-Associated Genes and ROS Production in BMMCs
3.3. The Transcriptional Signature of MCs Exposed to Cyclic Hypoxia
3.4. Cyclic Hypoxia Induces an Increase in the Transcription of Genes Encoding Elements of the FcεRI Signaling Pathway, Causing Enhanced Responsiveness to IgE/Ag Challenge
3.5. Cyclic Hypoxia-Induced Hyperresponsive Phenotype of MCs Is Associated with an Increase in Calcium Levels and Overactivation of Phospholipase C
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krystel-Whittemore, M.; Dileepan, K.N.; Wood, J.G. Mast Cell: A Multi-Functional Master Cell. Front. Immunol. 2016, 6, 620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wernersson, S.; Pejler, G. Mast Cell Secretory Granules: Armed for Battle. Nat. Rev. Immunol. 2014, 14, 478–494. [Google Scholar] [CrossRef] [PubMed]
- Kraft, S.; Kinet, J.P. New Developments in FcεRI Regulation, Function and Inhibition. Nat. Rev. Immunol. 2007, 7, 365–378. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Miranda, E.; Ibarra-Sanchez, A.; Gonzalez-Espinosa, C. Fyn Kinase Controls FcεRI Receptor-Operated Calcium Entry Necessary for Full Degranulation in Mast Cells. Biochem. Biophys. Res. Commun. 2010, 391, 1714–1720. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, D.; Yamasaki, S.; Arase, K.; Park, S.Y.; Arase, H.; Konno, A.; Saito, T. FcεRIγ-ITAM Is Differentially Required for Mast Cell Function In Vivo. J. Immunol. 2004, 172, 2374–2381. [Google Scholar] [CrossRef] [Green Version]
- Parravicini, V.; Gadina, M.; Kovarova, M.; Odom, S.; Gonzalez-Espinosa, C.; Furumoto, Y.; Saitoh, S.; Samelson, L.E.; O’Shea, J.J.; Rivera, J. Fyn Kinase Initiates Complementary Signals Required for IgE-Dependent Mast Cell Degranulation. Nat. Immunol. 2002, 3, 741–748. [Google Scholar] [CrossRef]
- Rivera, J.; Fierro, N.A.; Olivera, A.; Suzuki, R. New Insights on Mast Cell Activation via the High Affinity Receptor for IgE. Adv. Immunol. 2008, 98, 85–120. [Google Scholar] [CrossRef] [Green Version]
- Blank, U.; Rivera, J. The Ins and Outs of IgE-Dependent Mast-Cell Exocytosis. Trends Immunol. 2004, 25, 266–273. [Google Scholar] [CrossRef]
- Lundequist, A.; Pejler, G. Biological Implications of Preformed Mast Cell Mediators. Cell. Mol. Life Sci. 2011, 68, 965–975. [Google Scholar] [CrossRef]
- Blank, U.; Madera-Salcedo, I.K.; Danelli, L.; Claver, J.; Tiwari, N.; Sánchez-Miranda, E.; Vázquez-Victorio, G.; Ramírez-Valadez, K.A.; Macias-Silva, M.; González-Espinosa, C. Vesicular Trafficking and Signaling for Cytokine and Chemokine Secretion in Mast Cells. Front. Immunol. 2014, 5, 453. [Google Scholar] [CrossRef] [Green Version]
- Espinosa-Riquer, Z.P.; Segura-Villalobos, D.; Ramírez-Moreno, I.G.; Pérez Rodríguez, M.J.; Lamas, M.; Gonzalez-Espinosa, C. Signal Transduction Pathways Activated by Innate Immunity in Mast Cells: Translating Sensing of Changes into Specific Responses. Cells 2020, 9, 2411. [Google Scholar] [CrossRef] [PubMed]
- Valent, P.; Akin, C.; Hartmann, K.; Nilsson, G.; Reiter, A.; Hermine, O.; Sotlar, K.; Sperr, W.R.; Escribano, L.; George, T.I.; et al. Mast Cells as a Unique Hematopoietic Lineage and Cell System: From Paul Ehrlich’s Visions to Precision Medicine Concepts. Theranostics 2020, 10, 10743–10768. [Google Scholar] [CrossRef] [PubMed]
- Segura-Villalobos, D.; Ramírez-Moreno, I.G.; Martínez-Aguilar, M.; Ibarra-Sánchez, A.; Muñoz-Bello, J.O.; Anaya-Rubio, I.; Padilla, A.; Macías-Silva, M.; Lizano, M.; González-Espinosa, C. Mast Cell-Tumor Interactions: Molecular Mechanisms of Recruitment, Intratumoral Communication and Potential Therapeutic Targets for Tumor Growth. Cells 2022, 11, 349. [Google Scholar] [CrossRef]
- Varricchi, G.; Galdiero, M.R.; Loffredo, S.; Marone, G.; Iannone, R.; Marone, G.; Granata, F. Are Mast Cells MASTers in Cancer? Front. Immunol. 2017, 8, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dvorak, A.M.; Mihm, M.C., Jr.; Osage, J.E.; Dvorak, H.F. Melanoma. An Ultrastructural Study of the Host Inflammatory and Vascular Responses. J. Investig. Dermatol. 1980, 75, 388–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajabi, P.; Bagheri, A.; Hani, M. Intratumoral and Peritumoral Mast Cells in Malignant Melanoma: An Immunohistochemical Study. Adv. Biomed. Res. 2017, 6, 39. [Google Scholar] [CrossRef] [PubMed]
- Derakhshani, A.; Vahidian, F.; Alihasanzadeh, M.; Mokhtarzadeh, A.; Nezhad, P.L.; Baradaran, B. Mast Cells: A Double-Edged Sword in Cancer. Immunol. Lett. 2019, 209, 28–35. [Google Scholar] [CrossRef]
- Marichal, T.; Tsai, M.; Galli, S.J. Mast Cells: Potential Positive and Negative Roles in Tumor Biology. Cancer Immunol. Res. 2013, 1, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Albini, A.; Bruno, A.; Noonan, D.M.; Mortara, L. Contribution to Tumor Angiogenesis from Innate Immune Cells within the Tumor Microenvironment: Implications for Immunotherapy. Front. Immunol. 2018, 9, 527. [Google Scholar] [CrossRef]
- Oshi, M.; Tokumaru, Y.; Asaoka, M.; Yan, L.; Satyananda, V.; Matsuyama, R.; Matsuhashi, N.; Futamura, M.; Ishikawa, T.; Yoshida, K.; et al. M1 Macrophage and M1/M2 Ratio Defined by Transcriptomic Signatures Resemble Only Part of Their Conventional Clinical Characteristics in Breast Cancer. Sci. Rep. 2020, 10, 16554. [Google Scholar] [CrossRef]
- Varricchi, G.; de Paulis, A.; Marone, G.; Galli, S.J. Future Needs in Mast Cell Biology. Int. J. Mol. Sci. 2019, 20, 4397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Presta, I.; Donato, A.; Zaffino, P.; Spadea, M.F.; Mancuso, T.; Malara, N.; Chiefari, E.; Donato, G. Does a Polarization State Exist for Mast Cells in Cancer? Med. Hypotheses 2019, 131, 109281. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhao, Q.; Zhang, Y.; Liu, Z.; Zheng, Z.; Liu, S.; Meng, L.; Xin, Y.; Jiang, X. Targeting Hypoxia in the Tumor Microenvironment: A Potential Strategy to Improve Cancer Immunotherapy. J. Exp. Clin. Cancer Res. 2021, 40, 24. [Google Scholar] [CrossRef] [PubMed]
- Saxena, K.; Jolly, M.K. Acute vs. Chronic vs. Cyclic Hypoxia: Their Differential Dynamics, Molecular Mechanisms, and Effects on Tumor Progression. Biomolecules 2019, 9, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boareto, M.; Jolly, M.K.; Ben-Jacob, E.; Onuchic, J.N. Jagged Mediates Differences in Normal and Tumor Angiogenesis by Affecting Tip-Stalk Fate Decision. Proc. Natl. Acad. Sci. USA 2015, 112, E3836–E3844. [Google Scholar] [CrossRef] [Green Version]
- Bader, S.B.; Dewhirst, M.W.; Hammond, E.M. Cyclic Hypoxia: An Update on Its Characteristics, Methods to Measure It and Biological Implications in Cancer. Cancers 2020, 13, 23. [Google Scholar] [CrossRef]
- Jiménez-Andrade, G.Y.; Ibarra-Sánchez, A.; González, D.; Lamas, M.; González-Espinosa, C. Immunoglobulin E Induces VEGF Production in Mast Cells and Potentiates Their Pro-Tumorigenic Actions through a Fyn Kinase-Dependent Mechanism. J. Hematol. Oncol. 2013, 6, 56. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Moreno, I.G.; Ibarra-Sánchez, A.; Castillo-Arellano, J.I.; Blank, U.; González-Espinosa, C. Mast Cells Localize in Hypoxic Zones of Tumors and Secrete CCL-2 under Hypoxia through Activation of L-Type Calcium Channels. J. Immunol. 2020, 204, 1056–1068. [Google Scholar] [CrossRef]
- Gulliksson, M.; Carvalho, R.F.S.; Ullerås, E.; Nilsson, G. Mast Cell Survival and Mediator Secretion in Response to Hypoxia. PLoS ONE 2010, 5, e12360. [Google Scholar] [CrossRef] [Green Version]
- García-Román, J.; Ibarra-Sánchez, A.; Lamas, M.; Espinosa, C.G. VEGF Secretion during Hypoxia Depends on Free Radicals-Induced Fyn Kinase Activity in Mast Cells. Biochem. Biophys. Res. Commun. 2010, 401, 262–267. [Google Scholar] [CrossRef]
- Burry, R.W. Controls for Immunocytochemistry: An Update. J. Histochem. Cytochem. 2011, 59, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Pike, J.A.; Styles, I.B.; Rappoport, J.Z.; Heath, J.K. Quantifying Receptor Trafficking and Colocalization with Confocal Microscopy. Methods 2017, 115, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Tellier, C.; Desmet, D.; Petit, L.; Finet, L.; Graux, C.; Raes, M.; Feron, O.; Michiels, C. Cycling Hypoxia Induces a Specific Amplified Inflammatory Phenotype in Endothelial Cells and Enhances Tumor-Promoting Inflammation in Vivo. Neoplasia 2015, 17, 66–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Espinosa, C.; Odom, S.; Olivera, A.; Hobson, J.P.; Martinez, M.E.C.; Oliveira-dos-Santos, A.; Barra, L.; Spiegel, S.; Penninger, J.M.; Rivera, J. Preferential Signaling and Induction of Allergy-Promoting Lymphokines Upon Weak Stimulation of the High Affinity IgE Receptor on Mast Cells. J. Exp. Med. 2003, 197, 1453–1465. [Google Scholar] [CrossRef] [PubMed]
- Paivandy, A.; Eriksson, J.; Melo, F.R.; Sellin, M.E.; Pejler, G. Lysosomotropic Challenge of Mast Cells Causes Intra-Granular Reactive Oxygen Species Production. Cell Death Discov. 2019, 5, 95. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Cruz, M.; Coral-Vázquez, R.M.; Hernández-Stengele, G.; Sánchez, R.; Salazar, E.; Sanchez-Muñoz, F.; Encarnación-Guevara, S.; Ramírez-Salcedo, J. Identification of Putative Ortholog Gene Blocks Involved in Gestant and Lactating Mammary Gland Development: A Rodent Cross-Species Microarray Transcriptomics Approach. Int. J. Genom. 2013, 2013, 624681. [Google Scholar] [CrossRef] [Green Version]
- Luna-Moreno, D.; Vázquez-Martínez, O.; Báez-Ruiz, A.; Ramírez, J.; Díaz-Muñoz, M. Food Restricted Schedules Promote Differential Lipoperoxidative Activity in Rat Hepatic Subcellular Fractions. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 146, 632–643. [Google Scholar] [CrossRef]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A Web Server for Functional Enrichment Analysis and Functional Annotation of Gene Lists (2021 Update). Nucleic Acids Res. 2022. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Lee, H.-S.; Choi, E.-J.; Choi, H.; Lee, K.-S.; Kim, H.-R.; Na, B.-R.; Kwon, M.-S.; Jeong, G.-S.; Choi, H.G.; Choi, E.Y.; et al. Oral Administration of 4-Hydroxy-3-Methoxycinnamaldehyde Attenuates Atopic Dermatitis by Inhibiting T Cell and Keratinocyte Activation. PLoS ONE 2015, 10, e0144521. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, S.; Arudchandran, R.; Manetz, T.S.; Zhang, W.; Sommers, C.L.; Love, P.E.; Rivera, J.; Samelson, L.E. LAT is Essential for Fc(Epsilon)RI-Mediated Mast Cell Activation. Immunity 2000, 12, 525–535. [Google Scholar] [CrossRef] [Green Version]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A New Generation of Ca2+ Indicators with Greatly Improved Fluorescence Properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar] [CrossRef]
- Matsumoto, S.; Yasui, H.; Mitchell, J.B.; Krishna, M.C. Imaging Cycling Tumor Hypoxia. Cancer Res. 2010, 70, 10019–10023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michiels, C.; Tellier, C.; Feron, O. Cycling Hypoxia: A Key Feature of the Tumor Microenvironment. Biochim. Biophys. Acta 2016, 1866, 76–86. [Google Scholar] [CrossRef]
- Boidot, R.; Branders, S.; Helleputte, T.; Rubio, L.I.; Dupont, P.; Feron, O. A Generic Cycling Hypoxia-Derived Prognostic Gene Signature: Application to Breast Cancer Profiling. Oncotarget 2014, 5, 6947–6963. [Google Scholar] [CrossRef]
- Chen, W.-L.; Wang, C.-C.; Lin, Y.-J.; Wu, C.-P.; Hsieh, C.-H. Cycling Hypoxia Induces Chemoresistance through the Activation of Reactive Oxygen Species-Mediated B-Cell Lymphoma Extra-Long Pathway in Glioblastoma Multiforme. J. Transl. Med. 2015, 13, 389. [Google Scholar] [CrossRef] [Green Version]
- Martinive, P.; Defresne, F.; Quaghebeur, E.; Daneau, G.; Crokart, N.; Grégoire, V.; Gallez, B.; Dessy, C.; Feron, O. Impact of Cyclic Hypoxia on HIF-1alpha Regulation in Endothelial Cells—New Insights for Antitumor Treatments. FEBS J. 2009, 276, 509–518. [Google Scholar] [CrossRef] [Green Version]
- Snezhkina, A.V.; Kudryavtseva, A.V.; Kardymon, O.L.; Savvateeva, M.V.; Melnikova, N.V.; Krasnov, G.S.; Dmitriev, A.A. ROS Generation and Antioxidant Defense Systems in Normal and Malignant Cells. Oxid. Med. Cell. Longev. 2019, 2019, 6175804. [Google Scholar] [CrossRef]
- Cheung, E.C.; Vousden, K.H. The Role of ROS in Tumour Development and Progression. Nat. Rev. Cancer 2022, 22, 280–297. [Google Scholar] [CrossRef]
- Wardman, P. Fluorescent and Luminescent Probes for Measurement of Oxidative and Nitrosative Species in Cells and Tissues: Progress, Pitfalls, and Prospects. Free Radic. Biol. Med. 2007, 43, 995–1022. [Google Scholar] [CrossRef] [PubMed]
- Natoli, G.; Ostuni, R. Adaptation and Memory in Immune Responses. Nat. Immunol. 2019, 20, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Imbratta, C.; Hussein, H.; Andris, F.; Verdeil, G. C-MAF, a Swiss Army Knife for Tolerance in Lymphocytes. Front. Immunol. 2020, 11, 206. [Google Scholar] [CrossRef] [PubMed]
- Chinenov, Y.; Coppo, M.; Gupte, R.; Sacta, M.A.; Rogatsky, I. Glucocorticoid Receptor Coordinates Transcription Factor-Dominated Regulatory Network in Macrophages. BMC Genom. 2014, 15, 656. [Google Scholar] [CrossRef] [Green Version]
- Rottem, M.; Barbieri, S.; Kinet, J.P.; Metcalfe, D.D. Kinetics of the Appearance of Fc Epsilon RI-Bearing Cells in Interleukin-3-Dependent Mouse Bone Marrow Cultures: Correlation with Histamine Content and Mast Cell Maturation. Blood 1992, 79, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Nagata, Y.; Suzuki, R. FcεRI: A Master Regulator of Mast Cell Functions. Cells 2022, 11, 622. [Google Scholar] [CrossRef]
- Dolmetsch, R.E.; Lewis, R.S.; Goodnow, C.C.; Healy, J.I. Differential Activation of Transcription Factors Induced by Ca2+ Response Amplitude and Duration. Nature 1997, 386, 855–858. [Google Scholar] [CrossRef]
- Ribatti, D.; Annese, T.; Tamma, R. Controversial Role of Mast Cells in Breast Cancer Tumor Progression and Angiogenesis. Clin. Breast Cancer 2021, 21, 486–491. [Google Scholar] [CrossRef]
- Cheng, S.; Li, Z.; Gao, R.; Xing, B.; Gao, Y.; Yang, Y.; Qin, S.; Zhang, L.; Ouyang, H.; Du, P.; et al. A Pan-Cancer Single-Cell Transcriptional Atlas of Tumor Infiltrating Myeloid Cells. Cell 2021, 184, 792–809.e23. [Google Scholar] [CrossRef]
- Tóth-Jakatics, R.; Jimi, S.; Takebayashi, S.; Kawamoto, N. Cutaneous Malignant Melanoma: Correlation between Neovascularization and Peritumor Accumulation of Mast Cells Overexpressing Vascular Endothelial Growth Factor. Hum. Pathol. 2000, 31, 955–960. [Google Scholar] [CrossRef]
- Hui, L.; Chen, Y. Tumor Microenvironment: Sanctuary of the Devil. Cancer Lett. 2015, 368, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Dunn, G.P.; Bruce, A.T.; Ikeda, H.; Old, L.J.; Schreiber, R.D. Cancer Immunoediting: From Immunosurveillance to Tumor Escape. Nat. Immunol. 2002, 3, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Lugano, R.; Ramachandran, M.; Dimberg, A. Tumor Angiogenesis: Causes, Consequences, Challenges and Opportunities. Cell. Mol. Life Sci. 2020, 77, 1745–1770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varia, M.A.; Calkins-Adams, D.P.; Rinker, L.H.; Kennedy, A.S.; Novotny, D.B.; Fowler, W.C.; Raleigh, J.A. Pimonidazole: A Novel Hypoxia Marker for Complementary Study of Tumor Hypoxia and Cell Proliferation in Cervical Carcinoma. Gynecol. Oncol. 1998, 71, 270–277. [Google Scholar] [CrossRef]
- Schlüter, A.; Weller, P.; Kanaan, O.; Nel, I.; Heusgen, L.; Höing, B.; Haßkamp, P.; Zander, S.; Mandapathil, M.; Dominas, N.; et al. CD31 and VEGF Are Prognostic Biomarkers in Early-Stage, but Not in Late-Stage, Laryngeal Squamous Cell Carcinoma. BMC Cancer 2018, 18, 272. [Google Scholar] [CrossRef] [Green Version]
- Koyasu, S.; Tsuji, Y.; Harada, H.; Nakamoto, Y.; Nobashi, T.; Kimura, H.; Sano, K.; Koizumi, K.; Hamaji, M.; Togashi, K. Evaluation of Tumor-Associated Stroma and Its Relationship with Tumor Hypoxia Using Dynamic Contrast-Enhanced CT andF Misonidazole PET in Murine Tumor Models. Radiology 2016, 278, 734–741. [Google Scholar] [CrossRef] [Green Version]
- Carmona-Fontaine, C.; Deforet, M.; Akkari, L.; Thompson, C.B.; Joyce, J.A.; Xavier, J.B. Metabolic Origins of Spatial Organization in the Tumor Microenvironment. Proc. Natl. Acad. Sci. USA 2017, 114, 2934–2939. [Google Scholar] [CrossRef] [Green Version]
- Baudelet, C.; Cron, G.O.; Ansiaux, R.; Crokart, N.; DeWever, J.; Feron, O.; Gallez, B. The Role of Vessel Maturation and Vessel Functionality in Spontaneous Fluctuations of T2*-Weighted GRE Signal within Tumors. NMR Biomed. 2006, 19, 69–76. [Google Scholar] [CrossRef]
- Morgan, M.J.; Liu, Z. Crosstalk of Reactive Oxygen Species and NF-ΚB Signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Hwang, H.J.; Lynn, S.G.; Vengellur, A.; Saini, Y.; Grier, E.A.; Ferguson-Miller, S.M.; LaPres, J.J. Hypoxia Inducible Factors Modulate Mitochondrial Oxygen Consumption and Transcriptional Regulation of Nuclear-Encoded Electron Transport Chain Genes. Biochemistry 2015, 54, 3739–3748. [Google Scholar] [CrossRef] [Green Version]
- Klimova, T.; Chandel, N.S. Mitochondrial Complex III Regulates Hypoxic Activation of HIF. Cell Death Differ. 2008, 15, 660–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichiule, P.; Chavez, J.C.; LaManna, J.C. Hypoxic Regulation of Angiopoietin-2 Expression in Endothelial Cells. J. Biol. Chem. 2004, 279, 12171–12180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, K.; Kataoka, N. Regulation of Gene Expression under Hypoxic Conditions. Int. J. Mol. Sci. 2019, 20, 3278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowler, E.; Porazinski, S.; Uzor, S.; Thibault, P.; Durand, M.; Lapointe, E.; Rouschop, K.M.A.; Hancock, J.; Wilson, I.; Ladomery, M. Hypoxia Leads to Significant Changes in Alternative Splicing and Elevated Expression of CLK Splice Factor Kinases in PC3 Prostate Cancer Cells. BMC Cancer 2018, 18, 355. [Google Scholar] [CrossRef] [PubMed]
- Komi, D.E.A.; Redegeld, F.A. Role of Mast Cells in Shaping the Tumor Microenvironment. Clin. Rev. Allergy Immunol. 2020, 58, 313–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Lepus, C.M.; Raghu, H.; Reber, L.L.; Tsai, M.M.; Wong, H.H.; von Kaeppler, E.; Lingampalli, N.; Bloom, M.S.; Hu, N.; et al. IgE-Mediated Mast Cell Activation Promotes Inflammation and Cartilage Destruction in Osteoarthritis. Elife 2019, 8, e39905. [Google Scholar] [CrossRef]
- Li, M.; Yin, H.; Yan, Z.; Li, H.; Wu, J.; Wang, Y.; Wei, F.; Tian, G.; Ning, C.; Li, H.; et al. The Immune Microenvironment in Cartilage Injury and Repair. Acta Biomater. 2022, 140, 23–42. [Google Scholar] [CrossRef]
- Kaesler, S.; Röcken, M.; Biedermann, T.; Kaesler, S.; Wölbing, F.; Kempf, W.E.; Skabytska, Y.; Köberle, M.; Volz, T.; Sinnberg, T.; et al. Targeting Tumor-Resident Mast Cells for Effective Anti-Melanoma Immune Responses. JCI Insight 2019, 4, e125057. [Google Scholar] [CrossRef]
- Grützkau, A.; Smorodchenko, A.; Lippert, U.; Kirchhof, L.; Artuc, M.; Henz, B.M. LAMP-1 and LAMP-2, but Not LAMP-3, Are Reliable Markers for Activation-Induced Secretion of Human Mast Cells. Cytom. A 2004, 61, 62–68. [Google Scholar] [CrossRef]
- Raposo, G.; Tenza, D.; Mecheri, S.; Peronet, R.; Bonnerot, C.; Desaymard, C. Accumulation of Major Histocompatibility Complex Class II Molecules in Mast Cell Secretory Granules and Their Release upon Degranulation. Mol. Biol. Cell 1997, 8, 2631–2645. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Xiao, Y.; Li, Q.; Yao, J.; Yuan, X.; Zhang, Y.; Yin, X.; Saito, Y.; Fan, H.; Li, P.; et al. The Allergy Mediator Histamine Confers Resistance to Immunotherapy in Cancer Patients via Activation of the Macrophage Histamine Receptor H1. Cancer Cell 2022, 40, 36–52.e9. [Google Scholar] [CrossRef] [PubMed]
- Akula, S.; Paivandy, A.; Fu, Z.; Thorpe, M.; Pejler, G.; Hellman, L. How Relevant Are Bone Marrow-Derived Mast Cells (BMMCs) as Models for Tissue Mast Cells? A Comparative Transcriptome Analysis of BMMCs and Peritoneal Mast Cells. Cells 2020, 9, 2118. [Google Scholar] [CrossRef] [PubMed]
- Narravula, S.; Colgan, S.P. Hypoxia-Inducible Factor 1-Mediated Inhibition of Peroxisome Proliferator-Activated Receptor Alpha Expression during Hypoxia. J. Immunol. 2001, 166, 7543–7548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuura, T.; Takahashi, K.; Nakayama, K.; Kobayashi, T.; Choi-Miura, N.-H.; Tomita, M.; Kanayama, N. Increased Expression of Vascular Endothelial Growth Factor in Placentas of P57(Kip2) Null Embryos. FEBS Lett. 2002, 532, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, S.; Jana, M.; Zhou, Y.; Fung, Y.K.; Ghosh, S.; Pahan, K. Antineuroinflammatory Effect of NF-KappaB Essential Modifier-Binding Domain Peptides in the Adoptive Transfer Model of Experimental Allergic Encephalomyelitis. J. Immunol. 2004, 173, 1344–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.S.; Lee, H.-M.; Yoo, H.; Park, Y.K.; Jo, E.-K. Intracellular Signaling Pathways That Regulate Macrophage Chemokine Expression in Response to Mycobacterium Abscessus. J. Bacteriol. Virol. 2012, 42, 121. [Google Scholar] [CrossRef]
- Adebanjo, O.A.; Moonga, B.S.; Yamate, T.; Sun, L.; Minkin, C.; Abe, E.; Zaidi, M. Mode of Action of Interleukin-6 on Mature Osteoclasts. Novel Interactions with Extracellular Ca2+ Sensing in the Regulation of Osteoclastic Bone Resorption. J. Cell Biol. 1998, 142, 1347–1356. [Google Scholar] [CrossRef] [Green Version]
- Hassuneh, M.R.; Nagarkatti, P.S.; Nagarkatti, M. Evidence for the Participation of Interleukin-2 (IL-2) and IL-4 in the Regulation of Autonomous Growth and Tumorigenesis of Transformed Cells of Lymphoid Origin. Blood 1997, 89, 610–620. [Google Scholar] [CrossRef]
- Biedermann, T.; Mailhammer, R.; Mai, A.; Sander, C.; Ogilvie, A.; Brombacher, F.; Maier, K.; Levine, A.D.; Röcken, M. Reversal of Established Delayed Type Hypersensitivity Reactions Following Therapy with IL-4 or Antigen-Specific Th2 Cells. Eur. J. Immunol. 2001, 31, 1582–1591. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Annotation Term | Genes | p-Value | ||
|---|---|---|---|---|
| Transcription regulation (KW-0805) | Dnmt1 Dnmt3a | Nr3c1 Nr2e3 | Srf5 | |
| Transcription (KW-0804) | Brf1 Maf1 | Maf Mafb | Ccar1 Taf3 | |
| DNA binding (KW-0238) | Aebp2 Ash2l Pou3f3 | Pou4f1 Tead1 | Ang2 Sox4 | |
| Oxidative phosphorylation (mmu00190) | Atp5g2 Atp5j2 Atp6v1e1 Atp5l Atp4b | Ndufs4 Ndufb5 Ndufb2 Ndufb4 | Ndufa3 Ndufa5 Ndufc1 Cox10 | |
| Galactose metabolism (mmu00052) | B4galt1 B4galt2 | Akr1b7 Galt | Gale | |
| cAMP signaling pathway (mmu04024) | Atp1a2 Adcy4 | Edn2 | Ppp1cb | |
| Chemical carcinogenesis—Reactive oxygen species (mmu05208) | Cyp1b1 Gstm6 | Pik3cb | Sod1 | |
| Inflammatory response (KW-0395) | Bcl6 Ccl21a | Cxcl13 | Cx3cl1 | |
| Fc epsilon RI signaling pathway (mmu04664) | Fcer1g Alox5ap Mapk8 | Map2k6 Pik3cb Pla2g4a | Raf1 Vav3 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Segura-Villalobos, D.; Lamas, M.; González-Espinosa, C. Cyclic Hypoxia Induces Transcriptomic Changes in Mast Cells Leading to a Hyperresponsive Phenotype after FcεRI Cross-Linking. Cells 2022, 11, 2239. https://doi.org/10.3390/cells11142239
Segura-Villalobos D, Lamas M, González-Espinosa C. Cyclic Hypoxia Induces Transcriptomic Changes in Mast Cells Leading to a Hyperresponsive Phenotype after FcεRI Cross-Linking. Cells. 2022; 11(14):2239. https://doi.org/10.3390/cells11142239
Chicago/Turabian StyleSegura-Villalobos, Deisy, Monica Lamas, and Claudia González-Espinosa. 2022. "Cyclic Hypoxia Induces Transcriptomic Changes in Mast Cells Leading to a Hyperresponsive Phenotype after FcεRI Cross-Linking" Cells 11, no. 14: 2239. https://doi.org/10.3390/cells11142239