
MicroRNAs and Long Non-Coding RNAs in Adrenocortical Carcinoma

 ,
,  , , , , , ,
, , , , , ,  and
and
Abstract
:1. Introduction
2. Circulating miRNA in Adrenocortical Carcinoma
3. Tumorigenesis Pathways in Which Five of the Main ACC Circulating miRNAs Are Involved
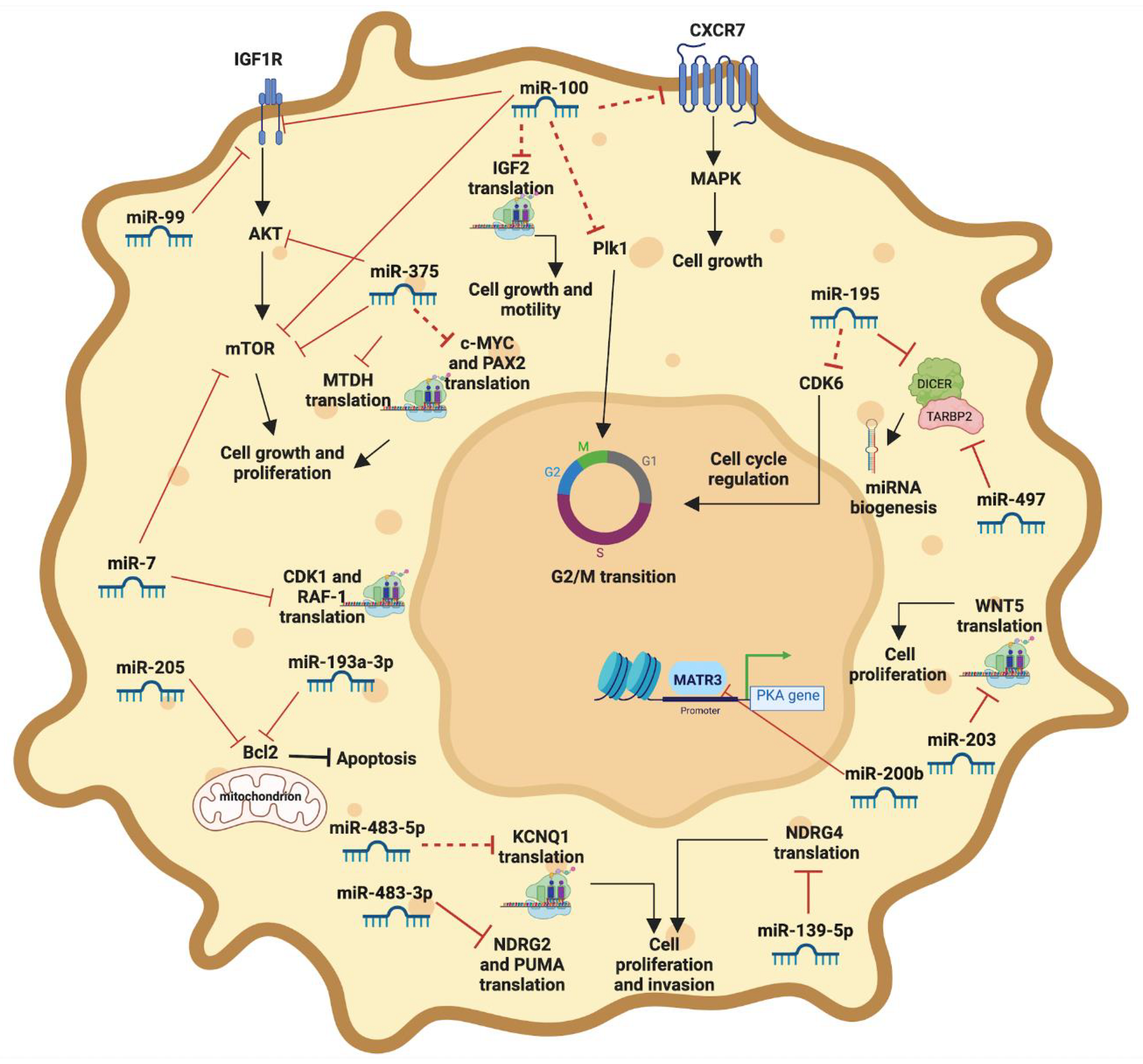
3.1. miR-100
3.2. miR-181b
3.3. miR-195
3.4. miR-483-5p
4. Circulating miRNAs Associated with Hormonal Secretion in the Adrenal
5. Tissue miRNAs in Adrenocortical Carcinoma
6. Tumorigenesis Pathways in Which Principal ACC Tissue miRNAs Are Involved
6.1. miR-184
6.2. miR-375
6.3. miR-483-3p
6.4. miR-503
7. Tissue-miRNAs Associated with Hormonal Secretion
8. Specific Role of miRs in the Pathogenesis of ACC
8.1. miR-7
8.2. miR-200b
8.3. miR-203
8.4. miR-193a-3p
8.5. miR-483-5p and miR-483-3p
8.6. miR-139-5p
8.7. miR-205
8.8. miR-375
8.9. miR-100 and miR-99
8.10. miR-195 and miR-497
9. Role of miRNA in the Prognosis and Therapy of ACC
10. Long Non-Coding RNAs in ACC
11. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fassnacht, M.; Dekkers, O.M.; Else, T.; Baudin, E.; Berruti, A.; de Krijger, R.; Haak, H.R.; Mihai, R.; Assie, G.; Terzolo, M. European Society of Endocrinology Clinical Practice Guidelines on the management of adrenocortical carcinoma in adults, in collaboration with the European Network for the Study of Adrenal Tumors. Eur. J. Endocrinol. 2018, 179, G1–G46. [Google Scholar] [CrossRef] [PubMed]
- Altieri, B.; Ronchi, C.L.; Kroiss, M.; Fassnacht, M. Next-generation therapies for adrenocortical carcinoma. Best Pract. Res. Clin. Endocrinol. Metab. 2020, 34, 101434. [Google Scholar] [CrossRef] [PubMed]
- Giordano, T.J.; Berney, D.; de Krijger, R.R.; Erickson, L.; Fassnacht, M.; Mete, O.; Papathomas, T.; Papotti, M.; Sasano, H.; Thompson, L.D.R.; et al. Data set for reporting of carcinoma of the adrenal cortex: Explanations and recommendations of the guidelines from the International Collaboration on Cancer Reporting. Hum. Pathol. 2021, 110, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Mete, O.; Erickson, L.A.; Juhlin, C.C.; de Krijger, R.R.; Sasano, H.; Volante, M.; Papotti, M.G. Overview of the 2022 WHO Classification of Adrenal Cortical Tumors. Endocr. Pathol. 2022, 33, 155–196. [Google Scholar] [CrossRef]
- Mete, O.; Gucer, H.; Kefeli, M.; Asa, S.L. Diagnostic and Prognostic Biomarkers of Adrenal Cortical Carcinoma. Am. J. Surg. Pathol. 2018, 42, 201–213. [Google Scholar] [CrossRef]
- Poli, G.; Ruggiero, C.; Cantini, G.; Canu, L.; Baroni, G.; Armignacco, R.; Jouinot, A.; Santi, R.; Ercolino, T.; Ragazzon, B.; et al. Fascin-1 Is a Novel Prognostic Biomarker Associated With Tumor Invasiveness in Adrenocortical Carcinoma. J. Clin. Endocrinol. Metab. 2019, 104, 1712–1724. [Google Scholar] [CrossRef]
- Altieri, B.; Sbiera, S.; Della Casa, S.; Weigand, I.; Wild, V.; Steinhauer, S.; Fadda, G.; Kocot, A.; Bekteshi, M.; Mambretti, E.M.; et al. Livin/BIRC7 expression as malignancy marker in adrenocortical tumors. Oncotarget 2017, 8, 9323–9338. [Google Scholar] [CrossRef] [Green Version]
- Mizdrak, M.; Ticinovic Kurir, T.; Bozic, J. The Role of Biomarkers in Adrenocortical Carcinoma: A Review of Current Evidence and Future Perspectives. Biomedicines 2021, 9, 174. [Google Scholar] [CrossRef]
- Terzolo, M.; Angeli, A.; Fassnacht, M.; Daffara, F.; Tauchmanova, L.; Conton, P.A.; Rossetto, R.; Buci, L.; Sperone, P.; Grossrubatscher, E.; et al. Adjuvant mitotane treatment for adrenocortical carcinoma. N. Engl. J. Med. 2007, 356, 2372–2380. [Google Scholar] [CrossRef] [Green Version]
- Altieri, B.; Lalli, E.; Faggiano, A. Mitotane treatment in adrenocortical carcinoma: Mechanisms of action and predictive markers of response to therapy. Minerva Endocrinol. 2021, 47, 203–214. [Google Scholar] [CrossRef]
- Basile, V.; Puglisi, S.; Calabrese, A.; Pia, A.; Perotti, P.; Berruti, A.; Reimondo, G.; Terzolo, M. Unwanted Hormonal and Metabolic Effects of Postoperative Adjuvant Mitotane Treatment for Adrenocortical Cancer. Cancers 2020, 12, 2615. [Google Scholar] [CrossRef]
- Altieri, B.; Sbiera, S.; Herterich, S.; De Francia, S.; Della Casa, S.; Calabrese, A.; Pontecorvi, A.; Quinkler, M.; Kienitz, T.; Mannelli, M.; et al. Effects of Germline CYP2W1*6 and CYP2B6*6 Single Nucleotide Polymorphisms on Mitotane Treatment in Adrenocortical Carcinoma: A Multicenter ENSAT Study. Cancers 2020, 12, 359. [Google Scholar] [CrossRef] [Green Version]
- Fassnacht, M.; Terzolo, M.; Allolio, B.; Baudin, E.; Haak, H.; Berruti, A.; Welin, S.; Schade-Brittinger, C.; Lacroix, A.; Jarzab, B.; et al. Combination chemotherapy in advanced adrenocortical carcinoma. N. Engl. J. Med. 2012, 366, 2189–2197. [Google Scholar] [CrossRef]
- Fassnacht, M.; Johanssen, S.; Quinkler, M.; Bucsky, P.; Willenberg, H.S.; Beuschlein, F.; Terzolo, M.; Mueller, H.H.; Hahner, S.; Allolio, B.; et al. Limited prognostic value of the 2004 International Union Against Cancer staging classification for adrenocortical carcinoma: Proposal for a Revised TNM Classification. Cancer 2009, 115, 243–250. [Google Scholar] [CrossRef]
- Libe, R.; Borget, I.; Ronchi, C.L.; Zaggia, B.; Kroiss, M.; Kerkhofs, T.; Bertherat, J.; Volante, M.; Quinkler, M.; Chabre, O.; et al. Prognostic factors in stage III-IV adrenocortical carcinomas (ACC): An European Network for the Study of Adrenal Tumor (ENSAT) study. Ann. Oncol. 2015, 26, 2119–2125. [Google Scholar] [CrossRef]
- Bilimoria, K.Y.; Shen, W.T.; Elaraj, D.; Bentrem, D.J.; Winchester, D.J.; Kebebew, E.; Sturgeon, C. Adrenocortical carcinoma in the United States: Treatment utilization and prognostic factors. Cancer 2008, 113, 3130–3136. [Google Scholar] [CrossRef]
- Vanbrabant, T.; Fassnacht, M.; Assie, G.; Dekkers, O.M. Influence of hormonal functional status on survival in adrenocortical carcinoma: Systematic review and meta-analysis. Eur. J. Endocrinol. 2018, 179, 429–436. [Google Scholar] [CrossRef] [Green Version]
- Beuschlein, F.; Weigel, J.; Saeger, W.; Kroiss, M.; Wild, V.; Daffara, F.; Libe, R.; Ardito, A.; Al Ghuzlan, A.; Quinkler, M.; et al. Major prognostic role of Ki67 in localized adrenocortical carcinoma after complete resection. J. Clin. Endocrinol. Metab. 2015, 100, 841–849. [Google Scholar] [CrossRef]
- Lippert, J.; Appenzeller, S.; Liang, R.; Sbiera, S.; Kircher, S.; Altieri, B.; Nanda, I.; Weigand, I.; Gehrig, A.; Steinhauer, S.; et al. Targeted Molecular Analysis in Adrenocortical Carcinomas: A Strategy Toward Improved Personalized Prognostication. J. Clin. Endocrinol. Metab. 2018, 103, 4511–4523. [Google Scholar] [CrossRef]
- Elhassan, Y.S.; Altieri, B.; Berhane, S.; Cosentini, D.; Calabrese, A.; Haissaguerre, M.; Kastelan, D.; Fragoso, M.; Bertherat, J.; Al Ghuzlan, A.; et al. S-GRAS score for prognostic classification of adrenocortical carcinoma: An international, multicenter ENSAT study. Eur. J. Endocrinol. 2021, 186, 25–36. [Google Scholar] [CrossRef]
- Assie, G.; Letouze, E.; Fassnacht, M.; Jouinot, A.; Luscap, W.; Barreau, O.; Omeiri, H.; Rodriguez, S.; Perlemoine, K.; Rene-Corail, F.; et al. Integrated genomic characterization of adrenocortical carcinoma. Nat. Genet. 2014, 46, 607–612. [Google Scholar] [CrossRef]
- De Martino, M.C.; Al Ghuzlan, A.; Aubert, S.; Assie, G.; Scoazec, J.Y.; Leboulleux, S.; Do Cao, C.; Libe, R.; Nozieres, C.; Lombes, M.; et al. Molecular screening for a personalized treatment approach in advanced adrenocortical cancer. J. Clin. Endocrinol. Metab. 2013, 98, 4080–4088. [Google Scholar] [CrossRef] [Green Version]
- Decmann, A.; Perge, P.; Turai, P.I.; Patocs, A.; Igaz, P. Non-Coding RNAs in Adrenocortical Cancer: From Pathogenesis to Diagnosis. Cancers 2020, 12, 461. [Google Scholar] [CrossRef] [Green Version]
- Chehade, M.; Bullock, M.; Glover, A.; Hutvagner, G.; Sidhu, S. Key MicroRNA’s and Their Targetome in Adrenocortical Cancer. Cancers 2020, 12, 2198. [Google Scholar] [CrossRef] [PubMed]
- Alles, J.; Fehlmann, T.; Fischer, U.; Backes, C.; Galata, V.; Minet, M.; Hart, M.; Abu-Halima, M.; Grasser, F.A.; Lenhof, H.P.; et al. An estimate of the total number of true human miRNAs. Nucleic Acids Res. 2019, 47, 3353–3364. [Google Scholar] [CrossRef] [Green Version]
- Yao, Q.; Chen, Y.; Zhou, X. The roles of microRNAs in epigenetic regulation. Curr. Opin. Chem. Biol. 2019, 51, 11–17. [Google Scholar] [CrossRef]
- Stroynowska-Czerwinska, A.; Fiszer, A.; Krzyzosiak, W.J. The panorama of miRNA-mediated mechanisms in mammalian cells. Cell. Mol. Life Sci. 2014, 71, 2253–2270. [Google Scholar] [CrossRef]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—microRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef]
- Oreglia, M.; Sbiera, S.; Fassnacht, M.; Guyon, L.; Denis, J.; Cristante, J.; Chabre, O.; Cherradi, N. Early Postoperative Circulating miR-483-5p Is a Prognosis Marker for Adrenocortical Cancer. Cancers 2020, 12, 724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebert, L.F.R.; MacRae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Gomes, A.Q.; Nolasco, S.; Soares, H. Non-coding RNAs: Multi-tasking molecules in the cell. Int. J. Mol. Sci. 2013, 14, 16010–16039. [Google Scholar] [CrossRef]
- Wu, P.; Mo, Y.; Peng, M.; Tang, T.; Zhong, Y.; Deng, X.; Xiong, F.; Guo, C.; Wu, X.; Li, Y.; et al. Emerging role of tumor-related functional peptides encoded by lncRNA and circRNA. Mol. Cancer 2020, 19, 22. [Google Scholar] [CrossRef]
- Panda, A.C. Circular RNAs Act as miRNA Sponges. Adv. Exp. Med. Biol. 2018, 1087, 67–79. [Google Scholar] [CrossRef]
- Igaz, P. Circulating microRNAs in adrenal tumors. Curr. Opin. Endocrinol. Diabetes Obes. 2019, 26, 155–159. [Google Scholar] [CrossRef]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. miRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef] [Green Version]
- Chabre, O.; Libe, R.; Assie, G.; Barreau, O.; Bertherat, J.; Bertagna, X.; Feige, J.J.; Cherradi, N. Serum miR-483-5p and miR-195 are predictive of recurrence risk in adrenocortical cancer patients. Endocr.-Relat. Cancer 2013, 20, 579–594. [Google Scholar] [CrossRef]
- Szabo, D.R.; Luconi, M.; Szabo, P.M.; Toth, M.; Szucs, N.; Horanyi, J.; Nagy, Z.; Mannelli, M.; Patocs, A.; Racz, K.; et al. Analysis of circulating microRNAs in adrenocortical tumors. Lab. Investig. 2014, 94, 331–339. [Google Scholar] [CrossRef] [Green Version]
- Salvianti, F.; Canu, L.; Poli, G.; Armignacco, R.; Scatena, C.; Cantini, G.; Di Franco, A.; Gelmini, S.; Ercolino, T.; Pazzagli, M.; et al. New insights in the clinical and translational relevance of miR483-5p in adrenocortical cancer. Oncotarget 2017, 8, 65525–65533. [Google Scholar] [CrossRef]
- Perge, P.; Butz, H.; Pezzani, R.; Bancos, I.; Nagy, Z.; Paloczi, K.; Nyiro, G.; Decmann, A.; Pap, E.; Luconi, M.; et al. Evaluation and diagnostic potential of circulating extracellular vesicle-associated microRNAs in adrenocortical tumors. Sci. Rep. 2017, 7, 5474. [Google Scholar] [CrossRef]
- Patel, D.; Boufraqech, M.; Jain, M.; Zhang, L.; He, M.; Gesuwan, K.; Gulati, N.; Nilubol, N.; Fojo, T.; Kebebew, E. MiR-34a and miR-483-5p are candidate serum biomarkers for adrenocortical tumors. Surgery 2013, 154, 1224–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decmann, A.; Perge, P.; Nyiro, G.; Darvasi, O.; Liko, I.; Borka, K.; Micsik, T.; Toth, Z.; Bancos, I.; Pezzani, R.; et al. MicroRNA Expression Profiling in Adrenal Myelolipoma. J. Clin. Endocrinol. Metab. 2018, 103, 3522–3530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decmann, A.; Bancos, I.; Khanna, A.; Thomas, M.A.; Turai, P.; Perge, P.; Pinter, J.Z.; Toth, M.; Patocs, A.; Igaz, P. Comparison of plasma and urinary microRNA-483-5p for the diagnosis of adrenocortical malignancy. J. Biotechnol. 2019, 297, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Shu, J.; Shi, G.; Yan, F.; Li, Y.; Ding, H. miR-100 Suppresses the Proliferation, Invasion, and Migration of Hepatocellular Carcinoma Cells via Targeting CXCR7. J. Immunol. Res. 2021, 2021, 9920786. [Google Scholar] [CrossRef]
- Chifu, I.; Heinze, B.; Fuss, C.T.; Lang, K.; Kroiss, M.; Kircher, S.; Ronchi, C.L.; Altieri, B.; Schirbel, A.; Fassnacht, M.; et al. Impact of the Chemokine Receptors CXCR4 and CXCR7 on Clinical Outcome in Adrenocortical Carcinoma. Front. Endocrinol. 2020, 11, 597878. [Google Scholar] [CrossRef]
- Seol, H.S.; Akiyama, Y.; Lee, S.E.; Shimada, S.; Jang, S.J. Loss of miR-100 and miR-125b results in cancer stem cell properties through IGF2 upregulation in hepatocellular carcinoma. Sci. Rep. 2020, 10, 21412. [Google Scholar] [CrossRef]
- Shi, W.; Alajez, N.M.; Bastianutto, C.; Hui, A.B.; Mocanu, J.D.; Ito, E.; Busson, P.; Lo, K.W.; Ng, R.; Waldron, J.; et al. Significance of Plk1 regulation by miR-100 in human nasopharyngeal cancer. Int. J. Cancer 2010, 126, 2036–2048. [Google Scholar] [CrossRef]
- De Martino, M.C.; van Koetsveld, P.M.; Feelders, R.A.; de Herder, W.W.; Dogan, F.; Janssen, J.; Hofste Op Bruinink, D.; Pivonello, C.; Waaijers, A.M.; Colao, A.; et al. IGF and mTOR pathway expression and in vitro effects of linsitinib and mTOR inhibitors in adrenocortical cancer. Endocrine 2019, 64, 673–684. [Google Scholar] [CrossRef] [Green Version]
- Altieri, B.; Colao, A.; Faggiano, A. The role of insulin-like growth factor system in the adrenocortical tumors. Minerva Endocrinol. 2019, 44, 43–57. [Google Scholar] [CrossRef] [Green Version]
- Altieri, B.; Tirabassi, G.; Della Casa, S.; Ronchi, C.L.; Balercia, G.; Orio, F.; Pontecorvi, A.; Colao, A.; Muscogiuri, G. Adrenocortical tumors and insulin resistance: What is the first step? Int. J. Cancer 2016, 138, 2785–2794. [Google Scholar] [CrossRef] [Green Version]
- Doghman, M.; El Wakil, A.; Cardinaud, B.; Thomas, E.; Wang, J.; Zhao, W.; Peralta-Del Valle, M.H.; Figueiredo, B.C.; Zambetti, G.P.; Lalli, E. Regulation of insulin-like growth factor-mammalian target of rapamycin signaling by microRNA in childhood adrenocortical tumors. Cancer Res. 2010, 70, 4666–4675. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Wang, W.; Hu, W.; Xu, W.; Xiao, G.; Nie, Q.; Ouyang, K.; Chen, S. MicroRNA-205 suppresses the growth of adrenocortical carcinoma SW-13 cells via targeting Bcl-2. Oncol. Rep. 2015, 34, 3104–3110. [Google Scholar] [CrossRef]
- Crona, J.; Beuschlein, F. Adrenocortical carcinoma—Towards genomics guided clinical care. Nat. Rev. Endocrinol. 2019, 15, 548–560. [Google Scholar] [CrossRef]
- Li, Y.; Fan, S.; Xia, W.; Qiao, B.; Huang, K.; Zhou, J.; Liang, M. MiR-181b suppresses angiogenesis by directly targeting cellular communication network factor 1. Lab. Investig. 2021, 101, 1026–1035. [Google Scholar] [CrossRef]
- Xu, X.; Ge, S.; Jia, R.; Zhou, Y.; Song, X.; Zhang, H.; Fan, X. Hypoxia-induced miR-181b enhances angiogenesis of retinoblastoma cells by targeting PDCD10 and GATA6. Oncol. Rep. 2015, 33, 2789–2796. [Google Scholar] [CrossRef] [Green Version]
- Furuta, M.; Kozaki, K.; Tanimoto, K.; Tanaka, S.; Arii, S.; Shimamura, T.; Niida, A.; Miyano, S.; Inazawa, J. The tumor-suppressive miR-497-195 cluster targets multiple cell-cycle regulators in hepatocellular carcinoma. PLoS ONE 2013, 8, e60155. [Google Scholar] [CrossRef]
- Hadjadj, D.; Kim, S.J.; Denecker, T.; Driss, L.B.; Cadoret, J.C.; Maric, C.; Baldacci, G.; Fauchereau, F. A hypothesis-driven approach identifies CDK4 and CDK6 inhibitors as candidate drugs for treatments of adrenocortical carcinomas. Aging 2017, 9, 2695–2716. [Google Scholar] [CrossRef] [Green Version]
- Caramuta, S.; Lee, L.; Ozata, D.M.; Akcakaya, P.; Xie, H.; Hoog, A.; Zedenius, J.; Backdahl, M.; Larsson, C.; Lui, W.O. Clinical and functional impact of TARBP2 over-expression in adrenocortical carcinoma. Endocr.-Relat. Cancer 2013, 20, 551–564. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; He, B.; Xu, J.; Li, Y.; Guo, C.; Cai, Q.; Wang, S. miR-483-5p Targets MKNK1 to Suppress Wilms’ Tumor Cell Proliferation and Apoptosis In Vitro and In Vivo. Med. Sci. Monit. 2019, 25, 1459–1468. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, H.; Zhu, S.; Lan, X. miR-483-5p promotes esophageal cancer progression by targeting KCNQ1. Biochem. Biophys. Res. Commun. 2020, 531, 615–621. [Google Scholar] [CrossRef]
- Ozata, D.M.; Caramuta, S.; Velazquez-Fernandez, D.; Akcakaya, P.; Xie, H.; Hoog, A.; Zedenius, J.; Backdahl, M.; Larsson, C.; Lui, W.O. The role of microRNA deregulation in the pathogenesis of adrenocortical carcinoma. Endocr.-Relat. Cancer 2011, 18, 643–655. [Google Scholar] [CrossRef] [Green Version]
- Perge, P.; Decmann, A.; Pezzani, R.; Bancos, I.; Fassina, A.; Luconi, M.; Canu, L.; Toth, M.; Boscaro, M.; Patocs, A.; et al. Analysis of circulating extracellular vesicle-associated microRNAs in cortisol-producing adrenocortical tumors. Endocrine 2018, 59, 280–287. [Google Scholar] [CrossRef] [Green Version]
- Igaz, I.; Nyiro, G.; Nagy, Z.; Butz, H.; Nagy, Z.; Perge, P.; Sahin, P.; Toth, M.; Racz, K.; Igaz, P.; et al. Analysis of Circulating MicroRNAs In Vivo following Administration of Dexamethasone and Adrenocorticotropin. Int. J. Endocrinol. 2015, 2015, 589230. [Google Scholar] [CrossRef] [Green Version]
- Vetrivel, S.; Zhang, R.; Engel, M.; Altieri, B.; Braun, L.; Osswald, A.; Bidlingmaier, M.; Fassnacht, M.; Beuschlein, F.; Reincke, M.; et al. Circulating microRNA Expression in Cushing’s Syndrome. Front. Endocrinol. 2021, 12, 620012. [Google Scholar] [CrossRef]
- Pivonello, C.; Patalano, R.; Negri, M.; Montò, T.; Amatrudo, F.; Di Paola, N.; Iacono, M.; Simeoli, C.; Ferrigno, R.; Solari, D.; et al. Circulating myomiRNAs as biomarkers of Cushing’s syndrome. Endocr. Abstr. 2021, 73, YI1. [Google Scholar] [CrossRef]
- Decmann, A.; Nyiro, G.; Darvasi, O.; Turai, P.; Bancos, I.; Kaur, R.J.; Pezzani, R.; Iacobone, M.; Kraljevic, I.; Kastelan, D.; et al. Circulating miRNA Expression Profiling in Primary Aldosteronism. Front. Endocrinol. 2019, 10, 739. [Google Scholar] [CrossRef]
- Belaya, Z.; Khandaeva, P.; Nonn, L.; Nikitin, A.; Solodovnikov, A.; Sitkin, I.; Grigoriev, A.; Pikunov, M.; Lapshina, A.; Rozhinskaya, L.; et al. Circulating Plasma microRNA to Differentiate Cushing’s Disease From Ectopic ACTH Syndrome. Front. Endocrinol. 2020, 11, 331. [Google Scholar] [CrossRef]
- Tombol, Z.; Szabo, P.M.; Molnar, V.; Wiener, Z.; Tolgyesi, G.; Horanyi, J.; Riesz, P.; Reismann, P.; Patocs, A.; Liko, I.; et al. Integrative molecular bioinformatics study of human adrenocortical tumors: MicroRNA, tissue-specific target prediction, and pathway analysis. Endocr.-Relat. Cancer 2009, 16, 895–906. [Google Scholar] [CrossRef] [Green Version]
- Soon, P.S.; Tacon, L.J.; Gill, A.J.; Bambach, C.P.; Sywak, M.S.; Campbell, P.R.; Yeh, M.W.; Wong, S.G.; Clifton-Bligh, R.J.; Robinson, B.G.; et al. miR-195 and miR-483-5p Identified as Predictors of Poor Prognosis in Adrenocortical Cancer. Clin. Cancer Res. 2009, 15, 7684–7692. [Google Scholar] [CrossRef] [Green Version]
- Koperski, L.; Kotlarek, M.; Swierniak, M.; Kolanowska, M.; Kubiak, A.; Gornicka, B.; Jazdzewski, K.; Wojcicka, A. Next-generation sequencing reveals microRNA markers of adrenocortical tumors malignancy. Oncotarget 2017, 8, 49191–49200. [Google Scholar] [CrossRef]
- Patterson, E.E.; Holloway, A.K.; Weng, J.; Fojo, T.; Kebebew, E. MicroRNA profiling of adrenocortical tumors reveals miR-483 as a marker of malignancy. Cancer 2011, 117, 1630–1639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duregon, E.; Rapa, I.; Votta, A.; Giorcelli, J.; Daffara, F.; Terzolo, M.; Scagliotti, G.V.; Volante, M.; Papotti, M. MicroRNA expression patterns in adrenocortical carcinoma variants and clinical pathologic correlations. Hum. Pathol. 2014, 45, 1555–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, S.; Cherniack, A.D.; Dewal, N.; Moffitt, R.A.; Danilova, L.; Murray, B.A.; Lerario, A.M.; Else, T.; Knijnenburg, T.A.; Ciriello, G.; et al. Comprehensive Pan-Genomic Characterization of Adrenocortical Carcinoma. Cancer Cell 2016, 29, 723–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz, K.J.; Helwig, J.; Bertram, S.; Sheu, S.Y.; Suttorp, A.C.; Seggewiss, J.; Willscher, E.; Walz, M.K.; Worm, K.; Schmid, K.W. Differential expression of microRNA-675, microRNA-139-3p and microRNA-335 in benign and malignant adrenocortical tumours. J. Clin. Pathol. 2011, 64, 529–535. [Google Scholar] [CrossRef]
- Feinmesser, M.; Benbassat, C.; Meiri, E.; Benjamin, H.; Lebanony, D.; Lebenthal, Y.; de Vries, L.; Drozd, T.; Spector, Y. Specific MicroRNAs Differentiate Adrenocortical Adenomas from Carcinomas and Correlate With Weiss Histopathologic System. Appl. Immunohistochem. Mol. Morphol. 2015, 23, 522–531. [Google Scholar] [CrossRef]
- Xie, Q.; Zhao, S.; Kang, R.; Wang, X. lncRNA SNHG11 facilitates prostate cancer progression through the upregulation of IGF1R expression and by sponging miR184. Int. J. Mol. Med. 2021, 48, 182. [Google Scholar] [CrossRef]
- Zhen, Y.; Liu, Z.; Yang, H.; Yu, X.; Wu, Q.; Hua, S.; Long, X.; Jiang, Q.; Song, Y.; Cheng, C.; et al. Tumor suppressor PDCD4 modulates miR-184-mediated direct suppression of C-MYC and BCL2 blocking cell growth and survival in nasopharyngeal carcinoma. Cell Death Dis. 2013, 4, e872. [Google Scholar] [CrossRef]
- Yang, S.; Yang, R.; Lin, R.; Si, L. MicroRNA-375 inhibits the growth, drug sensitivity and metastasis of human ovarian cancer cells by targeting PAX2. J. Buon 2019, 24, 2341–2346. [Google Scholar]
- He, X.X.; Chang, Y.; Meng, F.Y.; Wang, M.Y.; Xie, Q.H.; Tang, F.; Li, P.Y.; Song, Y.H.; Lin, J.S. MicroRNA-375 targets AEG-1 in hepatocellular carcinoma and suppresses liver cancer cell growth in vitro and in vivo. Oncogene 2012, 31, 3357–3369. [Google Scholar] [CrossRef] [Green Version]
- Fu, C.; Dong, W.; Wang, Z.; Li, H.; Qin, Q.; Li, B. The expression of miR-21 and miR-375 predict prognosis of esophageal cancer. Biochem. Biophys. Res. Commun. 2014, 446, 1197–1203. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, Q.; Li, M.; Jiang, S.; Wang, X. MicroRNA-375 inhibits colorectal cancer growth by targeting PIK3CA. Biochem. Biophys. Res. Commun. 2014, 444, 199–204. [Google Scholar] [CrossRef]
- Ganesan, P.; Piha-Paul, S.; Naing, A.; Falchook, G.; Wheler, J.; Janku, F.; Zinner, R.; Laday, S.; Kies, M.; Tsimberidou, A.M. Phase I clinical trial of lenalidomide in combination with temsirolimus in patients with advanced cancer. Investig. New Drugs 2013, 31, 1505–1513. [Google Scholar] [CrossRef]
- Wagle, N.; Grabiner, B.C.; Van Allen, E.M.; Hodis, E.; Jacobus, S.; Supko, J.G.; Stewart, M.; Choueiri, T.K.; Gandhi, L.; Cleary, J.M.; et al. Activating mTOR mutations in a patient with an extraordinary response on a phase I trial of everolimus and pazopanib. Cancer Discov. 2014, 4, 546–553. [Google Scholar] [CrossRef] [Green Version]
- Naing, A.; Kurzrock, R.; Burger, A.; Gupta, S.; Lei, X.; Busaidy, N.; Hong, D.; Chen, H.X.; Doyle, L.A.; Heilbrun, L.K.; et al. Phase I trial of cixutumumab combined with temsirolimus in patients with advanced cancer. Clin. Cancer Res. 2011, 17, 6052–6060. [Google Scholar] [CrossRef] [Green Version]
- De Martino, M.C.; Feelders, R.A.; Pivonello, C.; Simeoli, C.; Papa, F.; Colao, A.; Pivonello, R.; Hofland, L.J. The role of mTOR pathway as target for treatment in adrenocortical cancer. Endocr. Connect. 2019, 8, R144–R156. [Google Scholar] [CrossRef] [Green Version]
- De Martino, M.C.; van Koetsveld, P.M.; Feelders, R.A.; Sprij-Mooij, D.; Waaijers, M.; Lamberts, S.W.; de Herder, W.W.; Colao, A.; Pivonello, R.; Hofland, L.J. The role of mTOR inhibitors in the inhibition of growth and cortisol secretion in human adrenocortical carcinoma cells. Endocr.-Relat. Cancer 2012, 19, 351–364. [Google Scholar] [CrossRef] [Green Version]
- De Martino, M.C.; van Koetsveld, P.M.; Pivonello, R.; Hofland, L.J. Role of the mTOR pathway in normal and tumoral adrenal cells. Neuroendocrinology 2010, 92 (Suppl. 1), 28–34. [Google Scholar] [CrossRef] [Green Version]
- Provvisiero, D.P.; Negri, M.; de Angelis, C.; Di Gennaro, G.; Patalano, R.; Simeoli, C.; Papa, F.; Ferrigno, R.; Auriemma, R.S.; De Martino, M.C.; et al. Vitamin D reverts resistance to the mTOR inhibitor everolimus in hepatocellular carcinoma through the activation of a miR-375/oncogenes circuit. Sci. Rep. 2019, 9, 11695. [Google Scholar] [CrossRef] [Green Version]
- Pennanen, M.; Hagstrom, J.; Heiskanen, I.; Sane, T.; Mustonen, H.; Arola, J.; Haglund, C. C-myc expression in adrenocortical tumours. J. Clin. Pathol. 2018, 71, 129–134. [Google Scholar] [CrossRef]
- Cerquetti, L.; Sampaoli, C.; De Salvo, M.; Bucci, B.; Argese, N.; Chimento, A.; Vottari, S.; Marchese, R.; Pezzi, V.; Toscano, V.; et al. C-MYC modulation induces responsiveness to paclitaxel in adrenocortical cancer cell lines. Int. J. Oncol. 2015, 46, 2231–2240. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Cao, Y.; Su, T.; Jiang, Y.; Jiang, L.; Zhou, W.; Zhang, C.; Wang, W.; Ning, G. Downregulation of miR-375 in aldosterone-producing adenomas promotes tumour cell growth via MTDH. Clin. Endocrinol. 2015, 83, 581–589. [Google Scholar] [CrossRef]
- Lupini, L.; Pepe, F.; Ferracin, M.; Braconi, C.; Callegari, E.; Pagotto, S.; Spizzo, R.; Zagatti, B.; Lanuti, P.; Fornari, F.; et al. Over-expression of the miR-483-3p overcomes the miR-145/TP53 pro-apoptotic loop in hepatocellular carcinoma. Oncotarget 2016, 7, 31361–31371. [Google Scholar] [CrossRef] [Green Version]
- Veronese, A.; Visone, R.; Consiglio, J.; Acunzo, M.; Lupini, L.; Kim, T.; Ferracin, M.; Lovat, F.; Miotto, E.; Balatti, V.; et al. Mutated beta-catenin evades a microRNA-dependent regulatory loop. Proc. Natl. Acad. Sci. USA 2011, 108, 4840–4845. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Liu, L.; Deng, X.; Li, D.; Cai, H.; Ma, Y.; Jia, C.; Wu, B.; Fan, Y.; Lv, Z. MicroRNA 483-3p targets Pard3 to potentiate TGF-beta1-induced cell migration, invasion, and epithelial-mesenchymal transition in anaplastic thyroid cancer cells. Oncogene 2019, 38, 699–715. [Google Scholar] [CrossRef]
- Hikisz, P.; Kilianska, Z.M. PUMA, a critical mediator of cell death—oOne decade on from its discovery. Cell. Mol. Biol. Lett. 2012, 17, 646–669. [Google Scholar] [CrossRef]
- Wang, T.; Ge, G.; Ding, Y.; Zhou, X.; Huang, Z.; Zhu, W.; Shu, Y.; Liu, P. MiR-503 regulates cisplatin resistance of human gastric cancer cell lines by targeting IGF1R and BCL2. Chin. Med. J. 2014, 127, 2357–2362. [Google Scholar]
- Yang, Y.; Liu, L.; Zhang, Y.; Guan, H.; Wu, J.; Zhu, X.; Yuan, J.; Li, M. MiR-503 targets PI3K p85 and IKK-beta and suppresses progression of non-small cell lung cancer. Int. J. Cancer 2014, 135, 1531–1542. [Google Scholar] [CrossRef]
- Sun, Y.; Li, L.; Xing, S.; Pan, Y.; Shi, Y.; Zhang, L.; Shen, Q. miR-503-3p induces apoptosis of lung cancer cells by regulating p21 and CDK4 expression. Cancer Biomark. 2017, 20, 597–608. [Google Scholar] [CrossRef]
- Tombol, Z.; Turai, P.I.; Decmann, A.; Igaz, P. MicroRNAs and Adrenocortical Tumors: Where do we Stand on Primary Aldosteronism? Horm. Metab. Res. 2020, 52, 394–403. [Google Scholar] [CrossRef]
- Lenzini, L.; Caroccia, B.; Campos, A.G.; Fassina, A.; Belloni, A.S.; Seccia, T.M.; Kuppusamy, M.; Ferraro, S.; Skander, G.; Bader, M.; et al. Lower expression of the TWIK-related acid-sensitive K+ channel 2 (TASK-2) gene is a hallmark of aldosterone-producing adenoma causing human primary aldosteronism. J. Clin. Endocrinol. Metab. 2014, 99, E674–E682. [Google Scholar] [CrossRef] [Green Version]
- Robertson, S.; Diver, L.A.; Alvarez-Madrazo, S.; Livie, C.; Ejaz, A.; Fraser, R.; Connell, J.M.; MacKenzie, S.M.; Davies, E. Regulation of Corticosteroidogenic Genes by MicroRNAs. Int. J. Endocrinol. 2017, 2017, 2021903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Zou, X.; Liu, Q.; Xie, T.; Huang, R.; Kang, H.; Lai, C.; Zhu, J. MiR-193a-3p functions as a tumour suppressor in human aldosterone-producing adrenocortical adenoma by down-regulating CYP11B2. Int. J. Exp. Pathol. 2018, 99, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S.; MacKenzie, S.M.; Alvarez-Madrazo, S.; Diver, L.A.; Lin, J.; Stewart, P.M.; Fraser, R.; Connell, J.M.; Davies, E. MicroRNA-24 is a novel regulator of aldosterone and cortisol production in the human adrenal cortex. Hypertension 2013, 62, 572–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nusrin, S.; Tong, S.K.; Chaturvedi, G.; Wu, R.S.; Giesy, J.P.; Kong, R.Y. Regulation of CYP11B1 and CYP11B2 steroidogenic genes by hypoxia-inducible miR-10b in H295R cells. Mar. Pollut. Bull. 2014, 85, 344–351. [Google Scholar] [CrossRef]
- Juhlin, C.C.; Bertherat, J.; Giordano, T.J.; Hammer, G.D.; Sasano, H.; Mete, O. What Did We Learn from the Molecular Biology of Adrenal Cortical Neoplasia? From Histopathology to Translational Genomics. Endocr. Pathol. 2021, 32, 102–133. [Google Scholar] [CrossRef]
- Faillot, S.; Foulonneau, T.; Neou, M.; Espiard, S.; Garinet, S.; Vaczlavik, A.; Jouinot, A.; Rondof, W.; Septier, A.; Drougat, L.; et al. Genomic classification of benign adrenocortical lesions. Endocr.-Relat. Cancer 2021, 28, 79–95. [Google Scholar] [CrossRef]
- Glover, A.R.; Zhao, J.T.; Gill, A.J.; Weiss, J.; Mugridge, N.; Kim, E.; Feeney, A.L.; Ip, J.C.; Reid, G.; Clarke, S.; et al. MicroRNA-7 as a tumor suppressor and novel therapeutic for adrenocortical carcinoma. Oncotarget 2015, 6, 36675–36688. [Google Scholar] [CrossRef] [Green Version]
- Bimpaki, E.I.; Iliopoulos, D.; Moraitis, A.; Stratakis, C.A. MicroRNA signature in massive macronodular adrenocortical disease and implications for adrenocortical tumourigenesis. Clin. Endocrinol. 2010, 72, 744–751. [Google Scholar] [CrossRef]
- Peng, K.Y.; Chang, H.M.; Lin, Y.F.; Chan, C.K.; Chang, C.H.; Chueh, S.J.; Yang, S.Y.; Huang, K.H.; Lin, Y.H.; Wu, V.C.; et al. miRNA-203 Modulates Aldosterone Levels and Cell Proliferation by Targeting Wnt5a in Aldosterone-Producing Adenomas. J. Clin. Endocrinol. Metab. 2018, 103, 3737–3747. [Google Scholar] [CrossRef] [Green Version]
- Agosta, C.; Laugier, J.; Guyon, L.; Denis, J.; Bertherat, J.; Libe, R.; Boisson, B.; Sturm, N.; Feige, J.J.; Chabre, O.; et al. MiR-483-5p and miR-139-5p promote aggressiveness by targeting N-myc downstream-regulated gene family members in adrenocortical cancer. Int. J. Cancer. 2018, 143, 944–957. [Google Scholar] [CrossRef] [Green Version]
- Kwok, G.T.Y.; Zhao, J.T.; Glover, A.R.; Gill, A.J.; Clifton-Bligh, R.; Robinson, B.G.; Ip, J.C.Y.; Sidhu, S.B. microRNA-431 as a Chemosensitizer and Potentiator of Drug Activity in Adrenocortical Carcinoma. Oncologist 2019, 24, e241–e250. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Li, M.Y.; Liu, Y.; Vlantis, A.C.; Chan, J.Y.; Xue, L.; Hu, B.G.; Yang, S.; Chen, M.X.; Zhou, S.; et al. The role of microRNA in cisplatin resistance or sensitivity. Expert Opin. Ther. Targets 2020, 24, 885–897. [Google Scholar] [CrossRef]
- Nagy, Z.; Baghy, K.; Hunyadi-Gulyas, E.; Micsik, T.; Nyiro, G.; Racz, G.; Butz, H.; Perge, P.; Kovalszky, I.; Medzihradszky, K.F.; et al. Evaluation of 9-cis retinoic acid and mitotane as antitumoral agents in an adrenocortical xenograft model. Am. J. Cancer Res. 2015, 5, 3645–3658. [Google Scholar]
- Jung, S.; Nagy, Z.; Fassnacht, M.; Zambetti, G.; Weiss, M.; Reincke, M.; Igaz, P.; Beuschlein, F.; Hantel, C. Preclinical progress and first translational steps for a liposomal chemotherapy protocol against adrenocortical carcinoma. Endocr.-Relat. Cancer 2016, 23, 825–837. [Google Scholar] [CrossRef] [Green Version]
- Glover, A.R.; Zhao, J.T.; Ip, J.C.; Lee, J.C.; Robinson, B.G.; Gill, A.J.; Soon, P.S.; Sidhu, S.B. Long noncoding RNA profiles of adrenocortical cancer can be used to predict recurrence. Endocr.-Relat. Cancer 2015, 22, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Sonkoly, E.; Bata-Csorgo, Z.; Pivarcsi, A.; Polyanka, H.; Kenderessy-Szabo, A.; Molnar, G.; Szentpali, K.; Bari, L.; Megyeri, K.; Mandi, Y.; et al. Identification and characterization of a novel, psoriasis susceptibility-related noncoding RNA gene, PRINS. J. Biol. Chem. 2005, 280, 24159–24167. [Google Scholar] [CrossRef] [Green Version]
- Szegedi, K.; Sonkoly, E.; Nagy, N.; Nemeth, I.B.; Bata-Csorgo, Z.; Kemeny, L.; Dobozy, A.; Szell, M. The anti-apoptotic protein G1P3 is overexpressed in psoriasis and regulated by the non-coding RNA, PRINS. Exp. Dermatol. 2010, 19, 269–278. [Google Scholar] [CrossRef]
- Shiyanbola, O.; Hardin, H.; Hu, R.; Eickhoff, J.C.; Lloyd, R.V. Long Noncoding RNA Expression in Adrenal Cortical Neoplasms. Endocr. Pathol. 2020, 31, 385–391. [Google Scholar] [CrossRef]
- Liu, B.; Zhan, X.; Liu, C. Long Noncoding RNA MALAT1 Interacts with miR-124-3p to Modulate Osteosarcoma Progression by Targeting SphK1. J. Oncol. 2021, 2021, 8390165. [Google Scholar] [CrossRef]
- Kim, J.; Piao, H.L.; Kim, B.J.; Yao, F.; Han, Z.; Wang, Y.; Xiao, Z.; Siverly, A.N.; Lawhon, S.E.; Ton, B.N.; et al. Long noncoding RNA MALAT1 suppresses breast cancer metastasis. Nat. Genet. 2018, 50, 1705–1715. [Google Scholar] [CrossRef]
- Di Dalmazi, G.; Altieri, B.; Scholz, C.; Sbiera, S.; Luconi, M.; Waldman, J.; Kastelan, D.; Ceccato, F.; Chiodini, I.; Arnaldi, G.; et al. RNA Sequencing and Somatic Mutation Status of Adrenocortical Tumors: Novel Pathogenetic Insights. J. Clin. Endocrinol. Metab. 2020, 105, e4459–e4473. [Google Scholar] [CrossRef]
- Zhang, J.; Yin, M.; Peng, G.; Zhao, Y. CRNDE: An important oncogenic long non-coding RNA in human cancers. Cell Prolif. 2018, 51, e12440. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Tang, Y.; Xing, W.; Dong, W.; Wang, Z. LncRNA, CRNDE promotes osteosarcoma cell proliferation, invasion and migration by regulating Notch1 signaling and epithelial-mesenchymal transition. Exp. Mol. Pathol. 2018, 104, 19–25. [Google Scholar] [CrossRef]
- Du, D.X.; Lian, D.B.; Amin, B.H.; Yan, W. Long non-coding RNA CRNDE is a novel tumor promoter by modulating PI3K/AKT signal pathways in human gastric cancer. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 5392–5398. [Google Scholar] [CrossRef]
- Ronchi, C.L.; Sbiera, S.; Altieri, B.; Steinhauer, S.; Wild, V.; Bekteshi, M.; Kroiss, M.; Fassnacht, M.; Allolio, B. Notch1 pathway in adrenocortical carcinomas: Correlations with clinical outcome. Endocr.-Relat. Cancer. 2015, 22, 531–543. [Google Scholar] [CrossRef] [Green Version]
- Buishand, F.O.; Liu-Chittenden, Y.; Fan, Y.; Tirosh, A.; Gara, S.K.; Patel, D.; Meerzaman, D.; Kebebew, E. Adrenocortical tumors have a distinct, long, non-coding RNA expression profile and LINC00271 is downregulated in malignancy. Surgery 2020, 167, 224–232. [Google Scholar] [CrossRef]
- Li, W.; Liu, R.; Wei, D.; Zhang, W.; Zhang, H.; Huang, W.; Hao, L. Circular RNA circ-CCAC1 Facilitates Adrenocortical Carcinoma Cell Proliferation, Migration, and Invasion through Regulating the miR-514a-5p/C22orf46 Axis. BioMed Res. Int. 2020, 2020, 3501451. [Google Scholar] [CrossRef]


{kind=link}
| Tumor Entity | miRNA | Expression |
|---|---|---|
| Adrenocortical carcinoma | miR-483-5p | Overexpression vs. A |
| miR-101 | Overexpression vs. A | |
| miR-100 | Overexpression vs. A | |
| miR-181b | Overexpression vs. A | |
| miR-184 | Overexpression vs. A | |
| miR-210 | Overexpression vs. A | |
| miR-195 | Underexpression vs. A | |
| miR-335 | Underexpression vs. A | |
| Primary hypercortisolism | miR-22-3p | Overexpression vs. NA |
| miR-27a-3p | Overexpression vs. NA | |
| miR-320b | Overexpression vs. NA | |
| miR182-5p * | Overexpression vs. healthy | |
| miR133a-3p * | Overexpression vs. healthy | |
| miR200b-3p ** | Overexpression vs. healthy | |
| miR-449 *** | Overexpression vs. CPA | |
| miR-142 **** | Overexpression related to ARMC5 wild type | |
| Primary hyperaldosteronism *** | miR-30d-5p | Overexpression in hyperplasia vs. APA |
| miR-7-5p | Overexpression in hyperplasia vs. APA | |
| miR-30e-5p | Overexpression in hyperplasia vs. APA |
| Tumor Entity | miRNA | Expression |
|---|---|---|
| Adrenocortical carcinoma | miR-483-5p | Overexpression A |
| miR-184 | Overexpression N, A | |
| miR-503 | Overexpression N, A | |
| miR-508-3 | Overexpression N A | |
| miR-210 | Overexpression A, N | |
| miR-483-3p | Overexpression A | |
| miR-335 | Underexpression N, A | |
| miR195 | Underexpression A, N | |
| miR-511 | Underexpression A, N | |
| miR-214 | Underexpression A, N | |
| miR-497 | Underexpression A, N | |
| miR-7 | Underexpression N, A | |
| Primary hypercortisolism | miR-210 | Underexpression vs. ACC |
| miR-375 | Underexpression vs. N | |
| Primary hyperaldosteronism * | miR-23 * | Overexpression related to higher aldosterone secretion |
| miR-34a * | Overexpression related to higher aldosterone secretion | |
| miR-320a-3p | Overexpression vs. N | |
| miR-193a-3p | Underexpression vs. N | |
| miR-125a-5p | Underexpression vs. N | |
| miR-495-3p | Underexpression vs. N |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Detomas, M.; Pivonello, C.; Pellegrini, B.; Landwehr, L.-S.; Sbiera, S.; Pivonello, R.; Ronchi, C.L.; Colao, A.; Altieri, B.; De Martino, M.C. MicroRNAs and Long Non-Coding RNAs in Adrenocortical Carcinoma. Cells 2022, 11, 2234. https://doi.org/10.3390/cells11142234
Detomas M, Pivonello C, Pellegrini B, Landwehr L-S, Sbiera S, Pivonello R, Ronchi CL, Colao A, Altieri B, De Martino MC. MicroRNAs and Long Non-Coding RNAs in Adrenocortical Carcinoma. Cells. 2022; 11(14):2234. https://doi.org/10.3390/cells11142234
Chicago/Turabian StyleDetomas, Mario, Claudia Pivonello, Bianca Pellegrini, Laura-Sophie Landwehr, Silviu Sbiera, Rosario Pivonello, Cristina L. Ronchi, Annamaria Colao, Barbara Altieri, and Maria Cristina De Martino. 2022. "MicroRNAs and Long Non-Coding RNAs in Adrenocortical Carcinoma" Cells 11, no. 14: 2234. https://doi.org/10.3390/cells11142234