
Ferroptosis-Related Genes Are Potential Therapeutic Targets and the Model of These Genes Influences Overall Survival of NSCLC Patients


Abstract
:1. Introduction
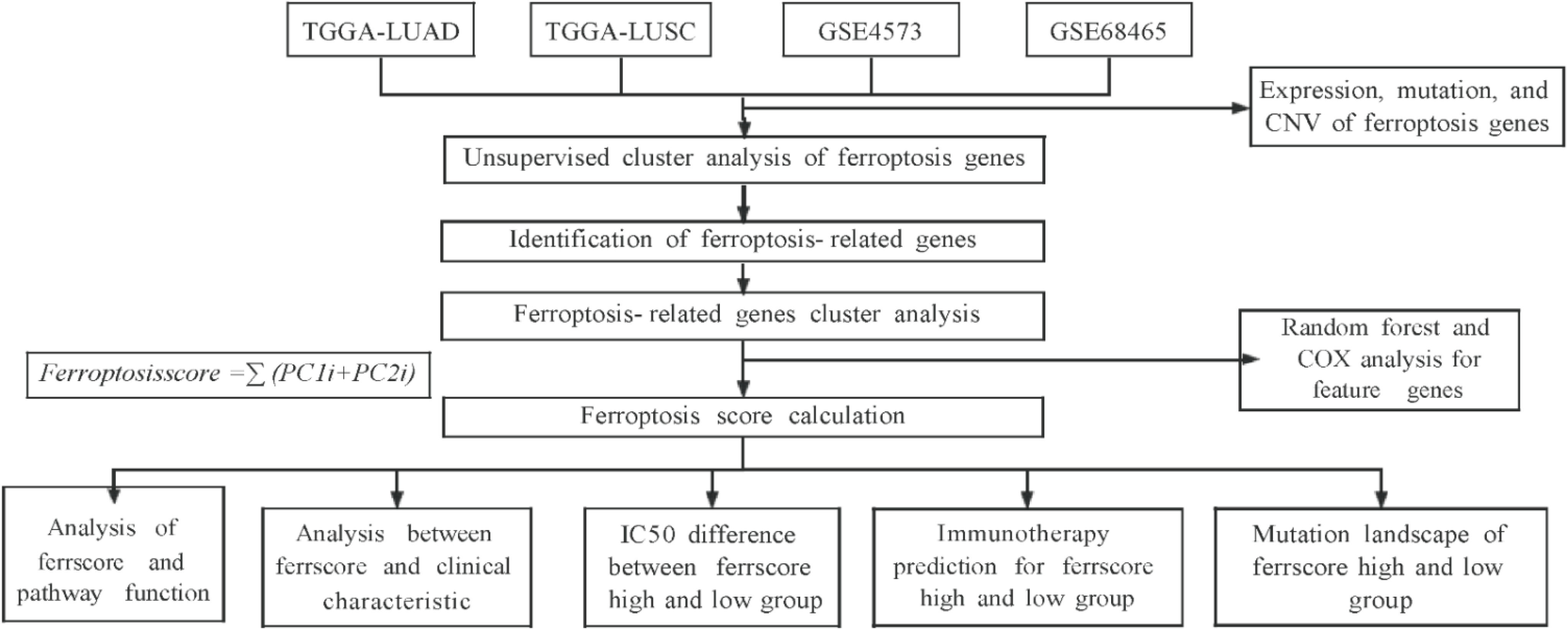
2. Methods
2.1. Data Collection
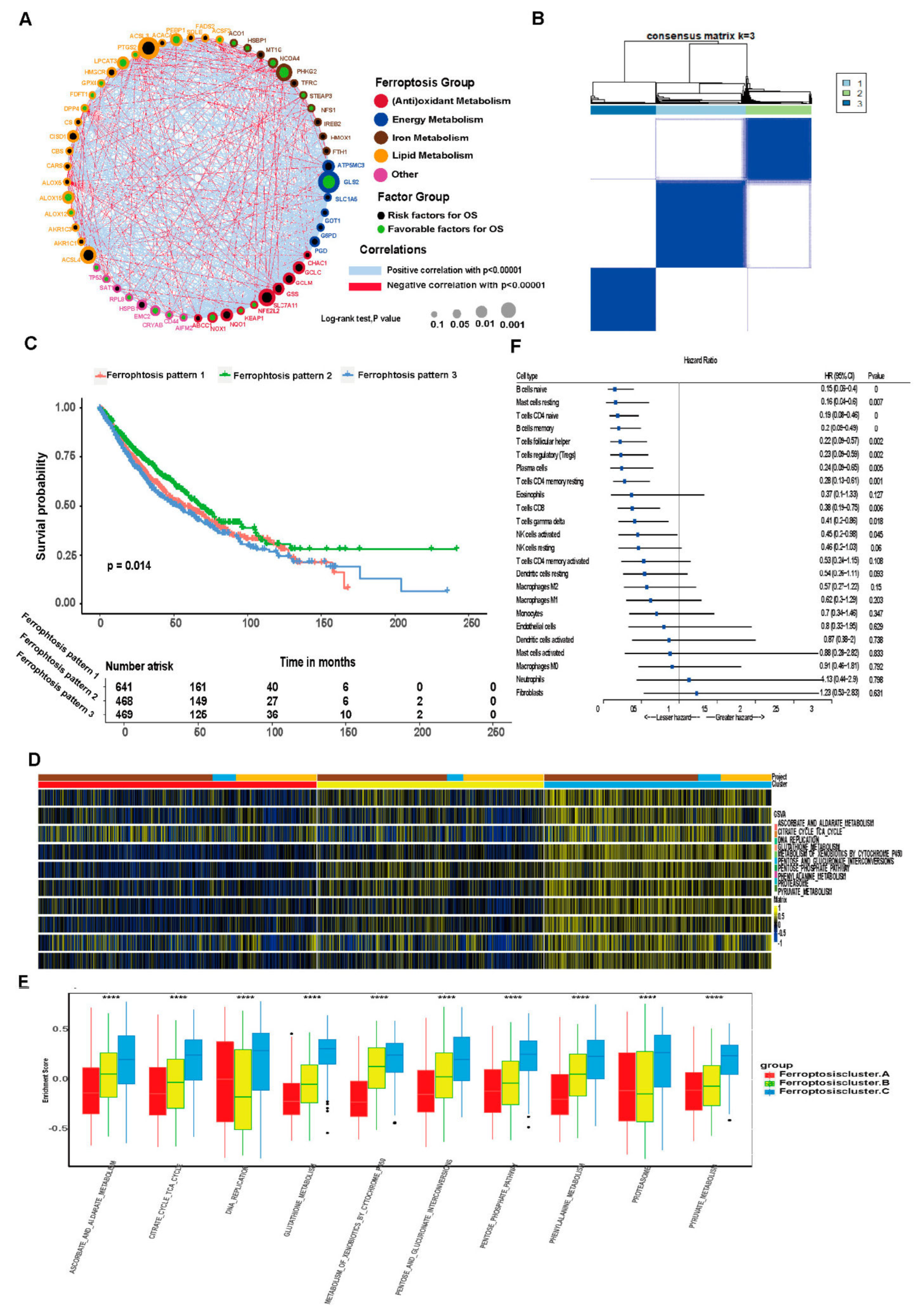
2.2. Gene Set Variation Analysis (GSVA) and Single-Sample Gene Set Enrichment Analysis (ssGSEA)
2.3. Model Establishment and Ferroptosis Score Calculation
2.4. Correlation between Ferroptosis Score and Other Biological Processes
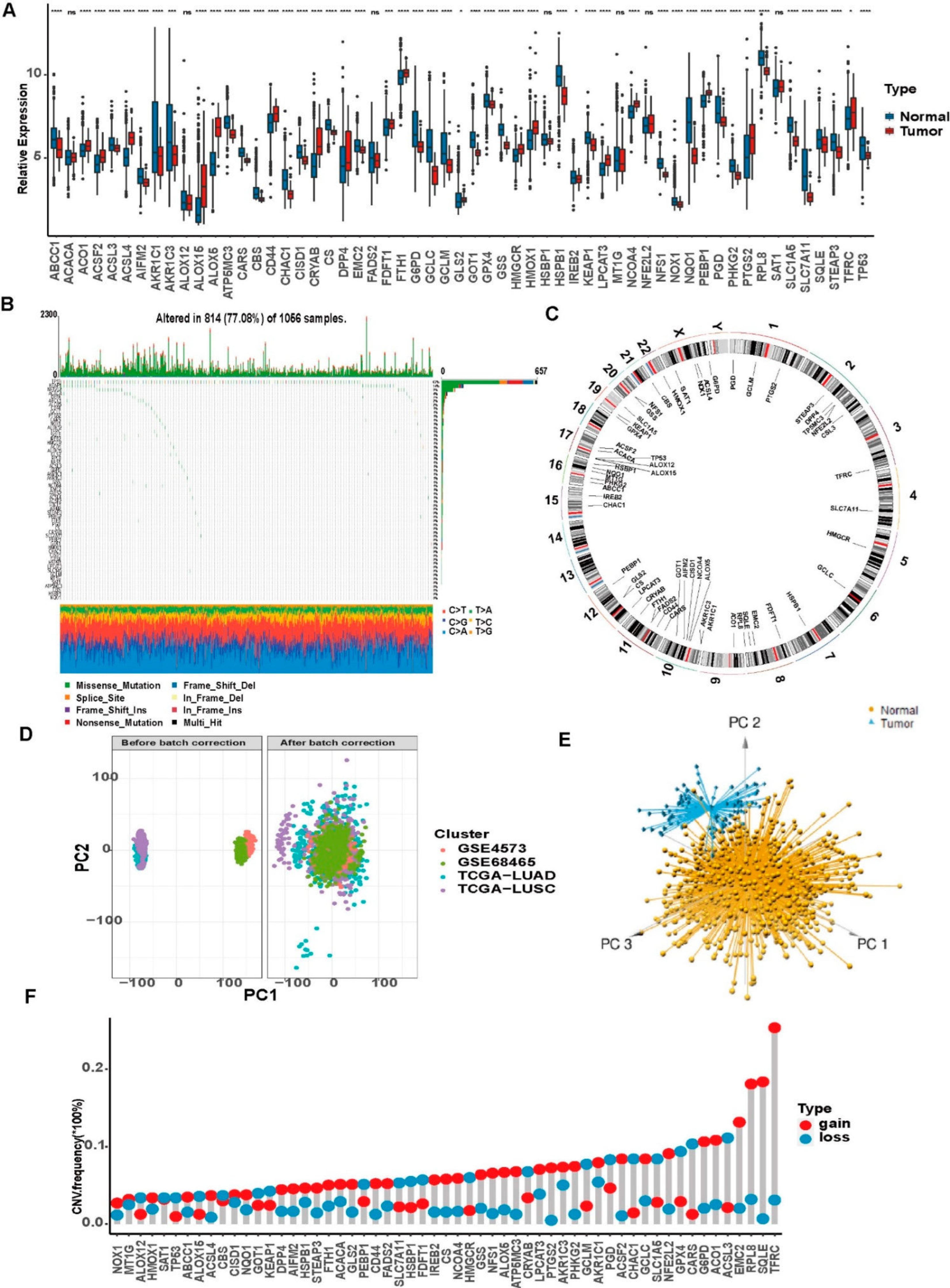
2.5. CNV Analysis
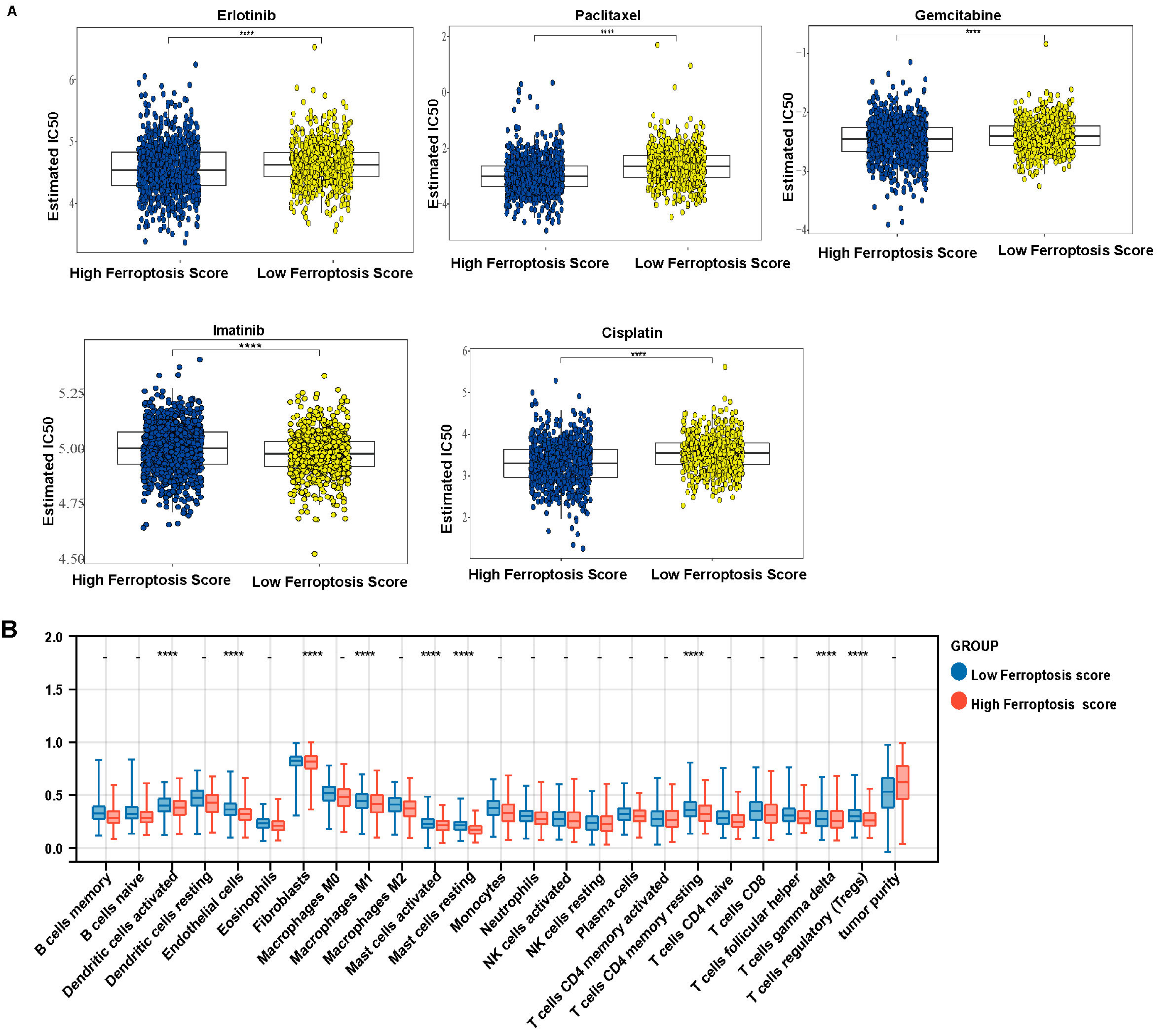
2.6. Tumor Half Maximal Inhibitory Concentration 50 (IC50)
2.7. Statistical Analysis
3. Results
3.1. Patient Characteristics
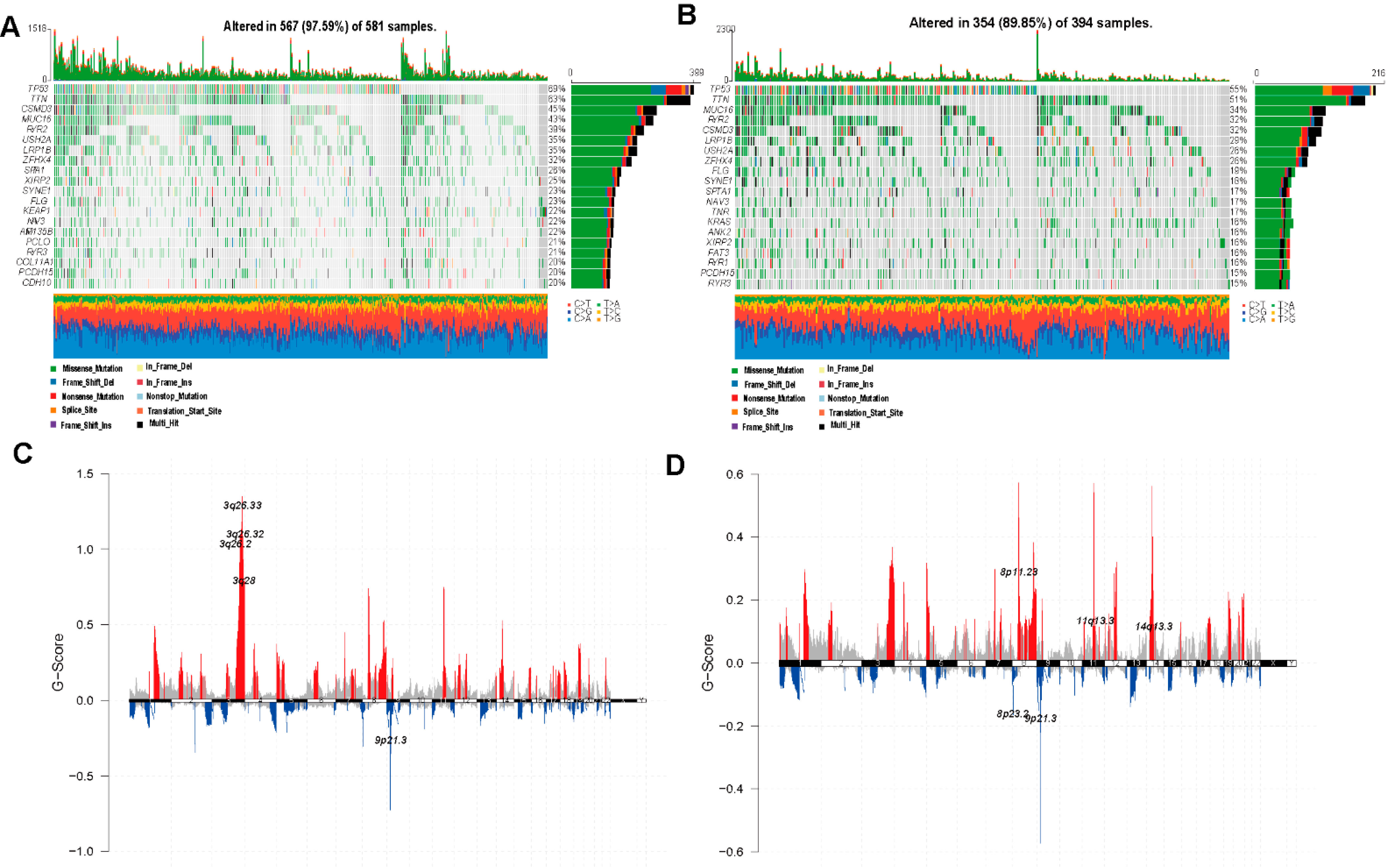
3.2. Genetic Variations of Ferroptosis-Related Genes in LUAD and LUSCC Patients
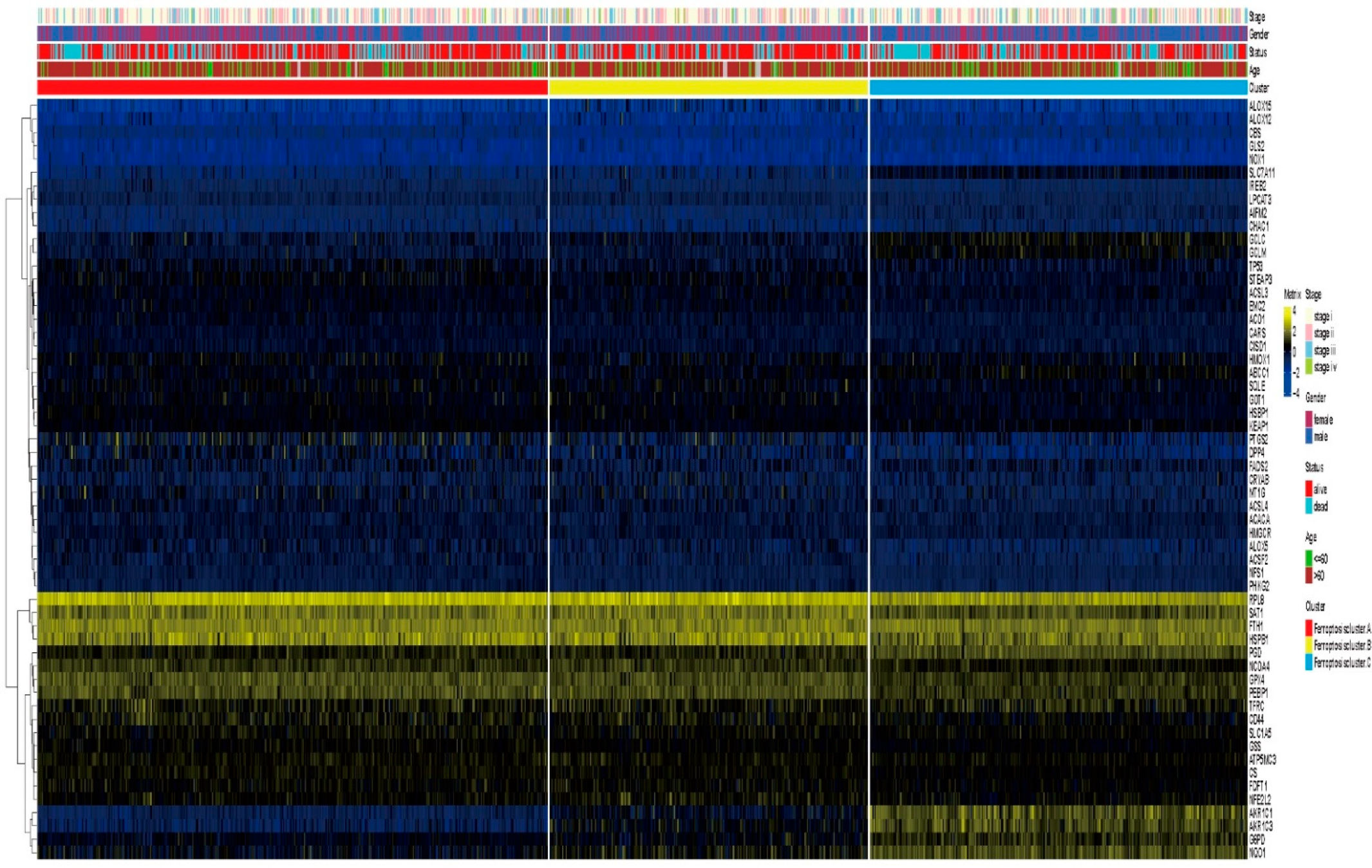
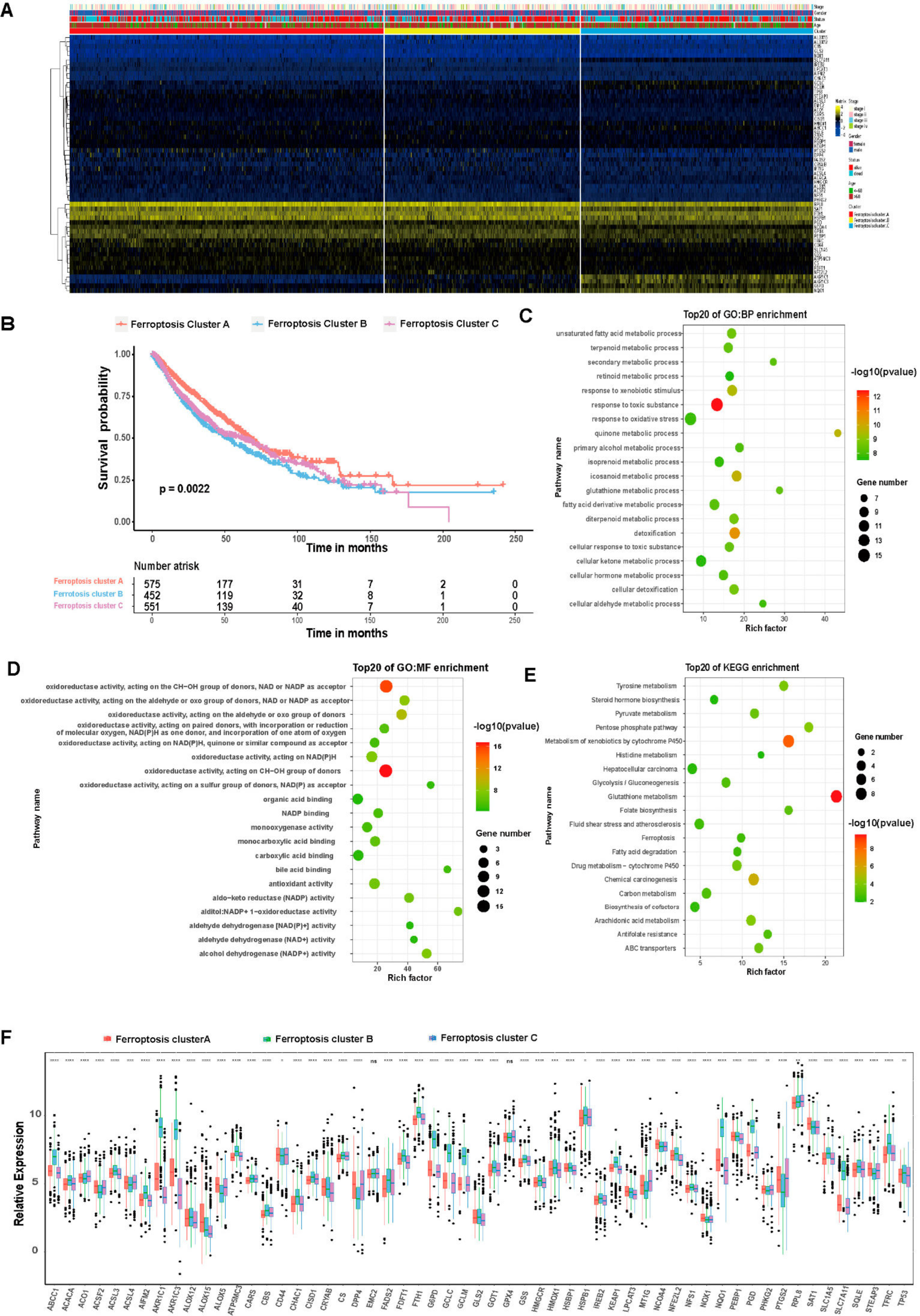
3.3. Unsupervised Clustering of Ferroptosis-Related Genes in LUAD and LUSCC Patients
3.4. Differential Gene Expression in Ferroptosis Patterns
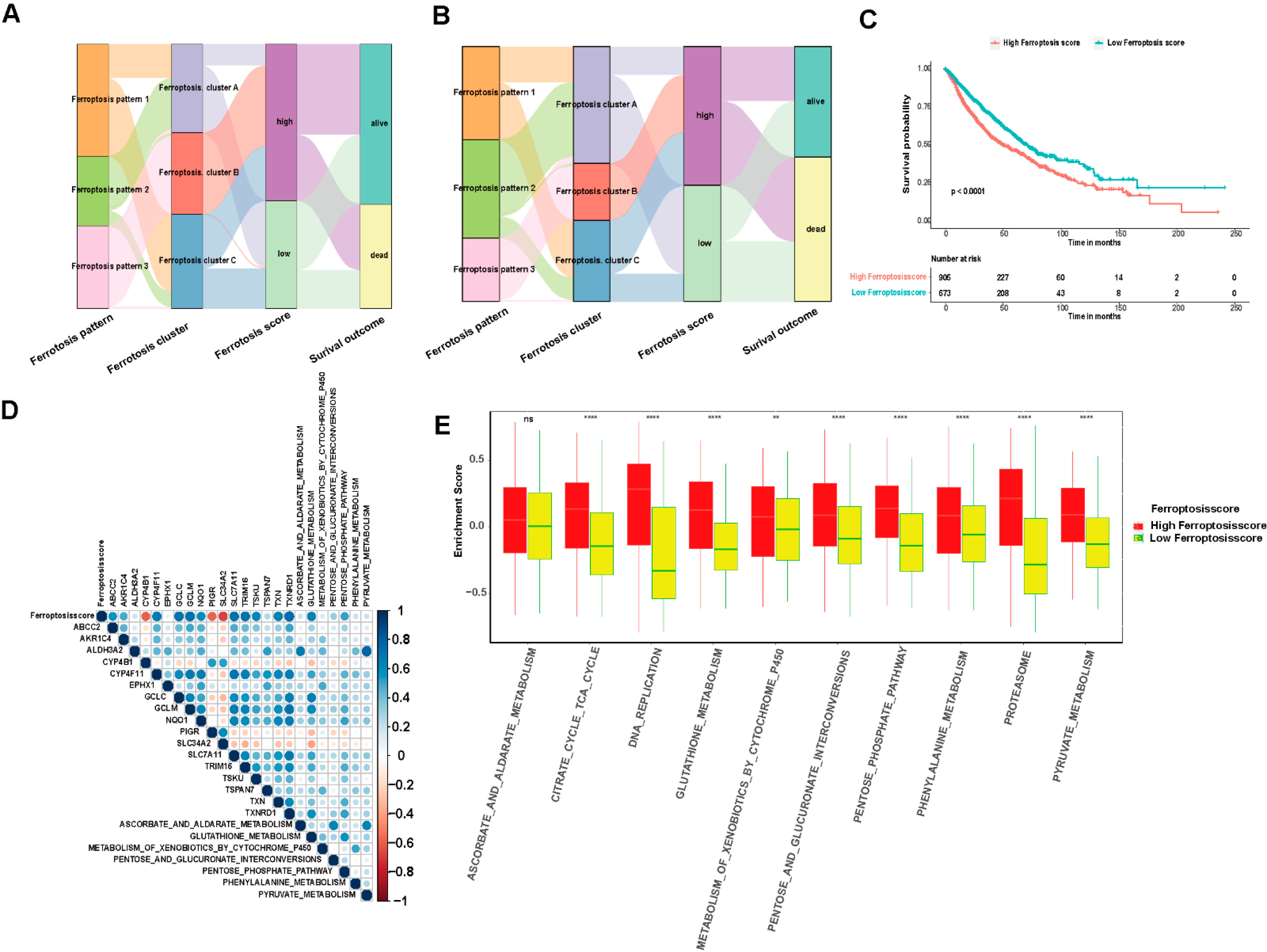
3.5. Model Establishment and Ferroptosis Score Analysis
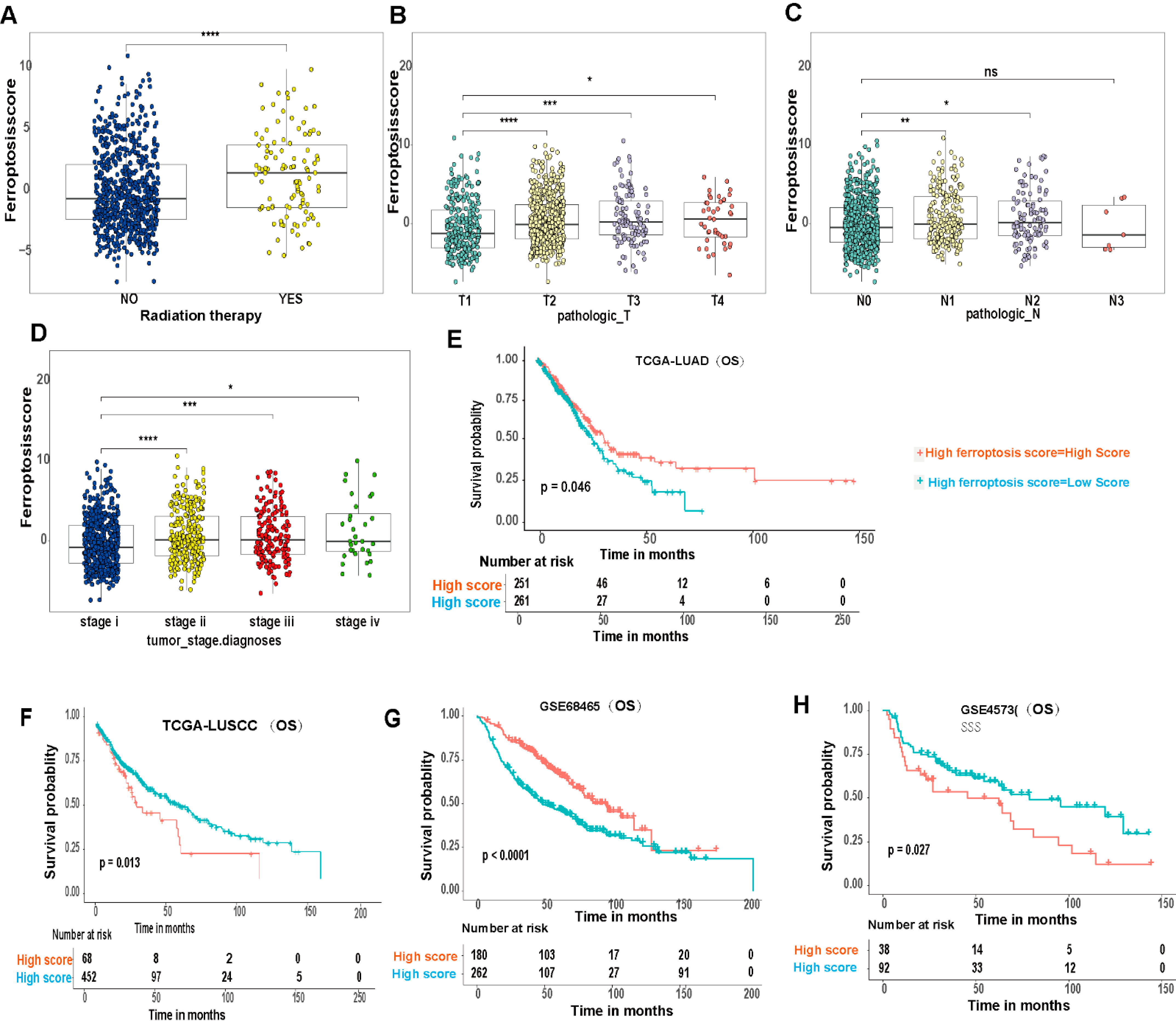
3.6. Ferroptosis Score Is Linked to Therapeutic Responses in LUAD and LUSCC Patients
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ahmad, S.; Qureshi, A.N.; Kazmi, A.; Rasool, A.; Gul, M.; Ashfaq, M.; Batool, L.; Rehman, R.A.; Ahmad, J.; Muniba, M. First cancer statistics report from Hazara division. J. Ayub Med. Coll. Abbottabad 2013, 25, 71–73. [Google Scholar] [PubMed]
- Bo, X.; Wang, J.; Wang, C.; Nan, L.; Gao, Z.; Xin, Y.; Li, M.; Shen, S.; Liu, H.; Ni, X.; et al. High infiltration of mast cells is associated with improved response to adjuvant chemotherapy in gallbladder cancer. Cancer Sci. 2020, 111, 817–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castelli, V.; Giordano, A.; Benedetti, E.; Giansanti, F.; Quintiliani, M.; Cimini, A.; d’Angelo, M. The great escape: The power of cancer stem cells to evade programmed cell death. Cancers 2021, 13, 328. [Google Scholar] [CrossRef]
- Catarata, M.J.; Ribeiro, R.; Oliveira, M.J.; Robalo Cordeiro, C.; Medeiros, R. Renin-angiotensin system in lung tumor and microenvironment interactions. Cancers 2020, 12, 1457. [Google Scholar] [CrossRef]
- Cheng, X.; Yang, S.; Xu, C.; Li, L.; Zhang, Y.; Guo, Y.; Zhang, C.; Li, P.; Long, M.; He, J. Proanthocyanidins protect against beta-hydroxybutyrate-induced oxidative damage in bovine endometrial cells. Molecules 2019, 24, 400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cronin, K.A.; Lake, A.J.; Scott, S.; Sherman, R.L.; Noone, A.M.; Howlader, N.; Henley, S.J.; Anderson, R.N.; Firth, A.U.; Ma, J.; et al. Annual report to the nation on the status of cancer, part I: National cancer statistics. Cancer 2018, 124, 2785–2800. [Google Scholar] [CrossRef] [Green Version]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Ou, Y.; Liu, H.; Zhan, L.; Zhu, X.; Cheng, M.; Li, Q.; Yin, D.; Liao, L. A ferroptosis-related prognostic signature based on antitumor immunity and tumor protein p53 mutation exploration for guiding treatment in patients with head and neck squamous cell carcinoma. Front. Genet. 2021, 12, 732211. [Google Scholar] [CrossRef]
- Gammella, E.; Recalcati, S.; Cairo, G. Dual role of ROS as signal and stress agents: Iron tips the balance in favor of toxic effects. Oxidative Med. Cell. Longev. 2016, 2016, 8629024. [Google Scholar] [CrossRef] [Green Version]
- Gaut, D.; Sim, M.S.; Yue, Y.; Wolf, B.R.; Abarca, P.A.; Carroll, J.M.; Goldman, J.W.; Garon, E.B. Clinical implications of the T790M mutation in disease characteristics and treatment response in patients with epidermal growth factor receptor (EGFR)-mutated non-small-cell lung cancer (NSCLC). Clin. Lung Cancer 2018, 19, e19–e28. [Google Scholar] [CrossRef]
- Ghoochani, A.; Hsu, E.C.; Aslan, M.; Rice, M.A.; Nguyen, H.M.; Brooks, J.D.; Corey, E.; Paulmurugan, R.; Stoyanova, T. Ferroptosis inducers are a novel therapeutic approach for advanced prostate cancer. Cancer Res. 2021, 81, 1583–1594. [Google Scholar] [CrossRef] [PubMed]
- Hangauer, M.J.; Viswanathan, V.S.; Ryan, M.J.; Bole, D.; Eaton, J.K.; Matov, A.; Galeas, J.; Dhruv, H.D.; Berens, M.E.; Schreiber, S.L.; et al. Drug-tolerant persister cancer cells are vulnerable to GPX4 inhibition. Nature 2017, 551, 247–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Wang, Y.; Li, C.; Zou, Z.; Liang, B. Development of an immune-related risk signature in patients with bladder urothelial carcinoma. BioMed Res. Int. 2020, 2020, 5848493. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.Y.; Wang, D.S.; Lin, H.C.; Chen, X.X.; Yang, H.; Zheng, Y.; Li, Y.H. A novel ferroptosis-related gene signature for overall survival prediction in patients with hepatocellular carcinoma. Int. J. Biol. Sci. 2020, 16, 2430–2441. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, M.; Xu, S.; Zhang, J.; Zou, J.; Yang, C.; Zhang, Y.; Gong, C.; Kai, Y.; Li, Y. HOXC8 promotes proliferation and migration through transcriptional up-regulation of TGFbeta1 in non-small cell lung cancer. Oncogenesis 2018, 7, 1. [Google Scholar] [CrossRef]
- Liu, W.; Li, C.; Wu, Y.; Xu, W.; Chen, S.; Zhang, H.; Huang, H.; Zhao, S.; Wang, J. Integrating m6A regulators-mediated methylation modification models and tumor immune microenvironment characterization in caucasian and chinese low-grade gliomas. Front. Cell Dev. Biol. 2021, 9, 725764. [Google Scholar] [CrossRef]
- Mariathasan, S.; Turley, S.J.; Nickles, D.; Castiglioni, A.; Yuen, K.; Wang, Y.; Kadel, E.E., III; Koeppen, H.; Astarita, J.L.; Cubas, R.; et al. TGFβ attenuates tumour response to PD-L1 blockade by contributing to exclusion of T cells. Nature 2018, 554, 544–548. [Google Scholar] [CrossRef]
- Mou, Y.; Wang, J.; Wu, J.; He, D.; Zhang, C.; Duan, C.; Li, B. Ferroptosis, a new form of cell death: Opportunities and challenges in cancer. J. Hematol. Oncol. 2019, 12, 34. [Google Scholar] [CrossRef] [Green Version]
- Nicot, C. RNA-Seq reveal the circular RNAs landscape of lung cancer. Mol. Cancer 2019, 18, 183. [Google Scholar] [CrossRef] [Green Version]
- Pasipoularides, A. Genomic translational research: Paving the way to individualized cardiac functional analyses and personalized cardiology. Int. J. Cardiol. 2017, 230, 384–401. [Google Scholar] [CrossRef] [Green Version]
- Qi, L.; Jiang, J.; Zhang, J.; Zhang, L.; Wang, T. Curcumin protects human trophoblast HTR8/SVneo cells from H2O2-induced oxidative stress by activating Nrf2 signaling pathway. Antioxidants 2020, 9, 121. [Google Scholar] [CrossRef] [Green Version]
- Rau, K.M.; Chen, H.K.; Shiu, L.Y.; Chao, T.L.; Lo, Y.P.; Wang, C.C.; Lin, M.C.; Huang, C.C. Discordance of mutation statuses of epidermal growth factor receptor and K-ras between primary adenocarcinoma of lung and brain metastasis. Int. J. Mol. Sci. 2016, 17, 524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seiler, A.; Schneider, M.; Förster, H.; Roth, S.; Wirth, E.K.; Culmsee, C.; Plesnila, N.; Kremmer, E.; Rådmark, O.; Wurst, W.; et al. Glutathione peroxidase 4 senses and translates oxidative stress into 12/15-lipoxygenase dependent- and AIF-mediated cell death. Cell Metab. 2008, 8, 237–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Şenbabaoğlu, Y.; Gejman, R.S.; Winer, A.G.; Liu, M.; Van Allen, E.M.; de Velasco, G.; Miao, D.; Ostrovnaya, I.; Drill, E.; Luna, A.; et al. Tumor immune microenvironment characterization in clear cell renal cell carcinoma identifies prognostic and immunotherapeutically relevant messenger RNA signatures. Genome Biol. 2016, 17, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Z.; Song, J.; Yung, B.C.; Zhou, Z.; Wu, A.; Chen, X. Emerging strategies of cancer therapy based on ferroptosis. Adv. Mater. 2018, 30, e1704007. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Taylor, W.R.; Fedorka, S.R.; Gad, I.; Shah, R.; Alqahtani, H.D.; Koranne, R.; Kuganesan, N.; Dlamini, S.; Rogers, T.; Al-Hamashi, A.; et al. Small-molecule ferroptotic agents with potential to selectively target cancer stem cells. Sci. Rep. 2019, 9, 5926. [Google Scholar] [CrossRef] [Green Version]
- Tu, G.; Peng, W.; Cai, Q.; Zhao, Z.; Peng, X.; He, B.; Zhang, P.; Shi, S.; Tao, Y.; Wang, X. Construction and validation of a 15-gene ferroptosis signature in lung adenocarcinoma. PeerJ 2021, 9, e11687. [Google Scholar] [CrossRef]
- Viswanathan, V.S.; Ryan, M.J.; Dhruv, H.D.; Gill, S.; Eichhoff, O.M.; Seashore-Ludlow, B.; Kaffenberger, S.D.; Eaton, J.K.; Shimada, K.; Aguirre, A.J.; et al. Dependency of a therapy-resistant state of cancer cells on a lipid peroxidase pathway. Nature 2017, 547, 453–457. [Google Scholar] [CrossRef]
- Wan, R.J.; Peng, W.; Xia, Q.X.; Zhou, H.H.; Mao, X.Y. Ferroptosis-related gene signature predicts prognosis and immunotherapy in glioma. CNS Neurosci. Ther. 2021, 27, 973–986. [Google Scholar] [CrossRef]
- Wang, W.; Gao, Z.; Wang, L.; Li, J.; Yu, J.; Han, S.; Meng, X. Application and Prospects of Molecular Imaging in Immunotherapy. Cancer Manag. Res. 2020, 12, 9389–9403. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.Y.; Chan, C.H.; Dubey, N.K.; Wei, H.J.; Lu, J.H.; Chang, C.C.; Cheng, H.C.; Ou, K.L.; Deng, W.P. Highly expressed FOXF1 inhibit non-small-cell lung cancer growth via inducing tumor suppressor and G1-phase cell-cycle arrest. Int. J. Mol. Sci. 2020, 21, 3227. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yu, C.; Luo, M.; Cen, C.; Qiu, J.; Zhang, S.; Hu, K. Ferroptosis in cancer treatment: Another way to Rome. Front. Oncol. 2020, 10, 571127. [Google Scholar] [CrossRef] [PubMed]
- Wylie, B.; Macri, C.; Mintern, J.D.; Waithman, J. Dendritic cells and cancer: From biology to therapeutic intervention. Cancers 2019, 11, 521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; He, Y.; Lin, L.; Chen, P.; Chen, M.; Zhang, S. The emerging role of ferroptosis in intestinal disease. Cell Death Dis. 2021, 12, 289. [Google Scholar] [CrossRef]
- Yang, W.S.; SriRamaratnam, R.; Welsch, M.E.; Shimada, K.; Skouta, R.; Viswanathan, V.S.; Cheah, J.H.; Clemons, P.A.; Shamji, A.F.; Clish, C.B.; et al. Regulation of ferroptotic cancer cell death by GPX4. Cell 2014, 156, 317–331. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Yin, H.; Shi, L.; Qian, X. A novel microRNA signature for pathological grading in lung adenocarcinoma based on TCGA and GEO data. Int. J. Mol. Med. 2020, 45, 1397–1408. [Google Scholar] [CrossRef] [Green Version]
- Ye, Z.; Liu, W.; Zhuo, Q.; Hu, Q.; Liu, M.; Sun, Q.; Zhang, Z.; Fan, G.; Xu, W.; Ji, S.; et al. Ferroptosis: Final destination for cancer? Cell Prolif. 2020, 53, e12761. [Google Scholar] [CrossRef]
- Yegya-Raman, N.; Aisner, J.; Kim, S.; Sayan, M.; Li, D.; Langenfeld, J.; Patel, M.; Malhotra, J.; Jabbour, S.K. Synchronous oligometastatic non-small cell lung cancer managed with curative-intent chemoradiation therapy: Long-term outcomes from a single institution. Adv. Radiat. Oncol. 2019, 4, 541–550. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Wu, Q.; Li, B.; Wang, D.; Wang, L.; Zhou, Y.L. m(6)A regulator-mediated methylation modification patterns and tumor microenvironment infiltration characterization in gastric cancer. Mol. Cancer 2020, 19, 53. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, X.; Yu, M.; Zhang, W.; Zhang, D.; Zeng, S.; Wang, X.; Hu, X. A novel ferroptosis-related gene model for overall survival predictions of bladder urothelial carcinoma patients. Front. Oncol. 2021, 11, 698856. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.J.; Wu, Z.; Ge, L.; Yang, F.; Hong, K.; Zhang, S.; Ma, L. Ferroptosis-Related Gene-Based Prognostic Model and Immune Infiltration in Clear Cell Renal Cell Carcinoma. Front. Genet. 2021, 12, 650416. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; You, J.; Yao, X.; Lu, Q.; Guo, W.; Shen, Y. Superparamagnetic iron oxide nanoparticles promote ferroptosis of ischemic cardiomyocytes. J. Cell. Mol. Med. 2020, 24, 11030–11033. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Bao, J. FerrDb: A manually curated resource for regulators and markers of ferroptosis and ferroptosis-disease associations. Database 2020, 2020, baaa021. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Training Cohort and Validation Cohort | ||||
|---|---|---|---|---|---|
| TCGA-LUAD (878 Cases) | TCGA-LUSCC (765 Cases) | GSE4573 (130 Cases) | GSE68465 (462 Cases) | ||
| RNA-seq cases | 585 (66.62%) | 550 (71.89%) | 130 (100%) | 462 (100%) | |
| Ferroptosis gene number | 60 | 60 | 57 (missing “AKR1C2”,”FANC D2”,”ZEB1”) | 57 (missing “AKR1C2”,”FANC D2”,”ZEB1”) | |
| SNV Cases number | 567 (64.59%) | 492 (64.31%) | 0 | 0 | |
| CNV Cases number | 555 (63.21) | 524 (68.50%) | 0 | 0 | |
| Cases with phenotypes | 877 (99.89%) | 765 (100%) | 130 (100%) | 462 (100%) | |
| Cases with survival information | 738 (84.05%) | 757 (98.95%) | 130 (100%) | 462 (100%) | |
| Gender | Female | 409 (46.58%) | 207 (20.06%) | 48 (36.92%) | 220 (47.62%) |
| Male | 469 (53.43%) | 558 (72.94%) | 82 (63.08%) | 242 (52.38%) | |
| Age | <65 | 330 (37.59) | 258 (33.73%) | 31 (23.85%) | 214 (46.32%) |
| ≥65 | 402 (45.79%) | 498 (65.10%) | 99 (76.15%) | 229 (49.59%) | |
| Unknown | 146 (16.63%) | 9 (1.18%) | 0 | 19 (4.11%) | |
| T classification | T1 | 253 (28.82%) | 167 (21.83%) | 33 (25.38%) | 150 (32.47%) |
| T2 | 401 (45.67) | 466 (60.92%) | 76 (58.46) | 251 (54.33%) | |
| T3 | 70 (7.97%) | 95 (12.42%) | 15 (11.54%) | 28 (6.06%) | |
| T4 | 21 (2.39%) | 37 (4.84%) | 6 (4.62%) | 12 (2.6%) | |
| Unknown | 132 (15.03%) | 0 | 0 | 21 (4.55%) | |
| N classification | N0 | 486 (55.35%) | 485 (63.40%) | 83 (63.85%) | 299 (64.72) |
| N1 | 135 (15.38%) | 199 (26.01%) | 32 (24.62%) | 88 (19.05%) | |
| N2 | 106 (12.07%) | 64 (8.37%) | 15 (11.54%) | 52 (11.26%) | |
| N3 | 2 (0.22%) | 10 (1.31%) | 0 | 0 | |
| Unknown | 148 (16.86%) | 0 | 0 | 23 (4.98%) | |
| M classification | M0 | 483 (55.01%) | 625 (81.70%) | 129 (99.23%) | NA |
| M1 | 37 (4.21%) | 12 (1.57%) | 0 | NA | |
| Other | 357 (40.66%) | 129 (16.86%) | 1 (0.77%) | NA | |
| Tumor stage | Stage I | 409 (46.58%) | 380 (49.67%) | 73 (56.15%) | NA |
| Stage II | 76 (8.66%) | 235 (30.72%) | 34 (26.15%) | NA | |
| Stage III | 118 (13.44%) | 131 (17.12%) | 23 (17.69%) | NA | |
| Stage IV | 38 (4.33%) | 12 (1.57%) | 0 | NA | |
| Unknown | 10 (1.14%) | 7 (0.92%) | 0 | NA | |
| Tobacco smoking | Ever | 622 (70.84%) | 730 (95.42%) | 120 (92.31) | 301 (65.15%) |
| History | Never | 108 (12.3%) | 22 (2.88%) | 4 (3.08%) | 49 (10.61%) |
| Unknown | 148 (16.86%) | 13 (1.70%) | 6 (4.62%) | 112 (24.24%) | |
| Number pack years | <30 | 174 (19.82%) | 122 (15.95%) | NA | NA |
| Smoked | >=30 | 327 (37.24%) | 499 (65.23%) | NA | NA |
| Unknown | 377 (42.94%) | 144 (18.82%) | NA | NA | |
| Radiation therapy | Yes | 87 (9.9%) | 70 (9.15%) | NA | NA |
| No | 556 (63.33%) | 534 (69.80%) | NA | NA | |
| Unknown | 235 (26.77%) | 161 (21.05%) | NA | NA | |
| Chemotherapy | Yes | NA | NA | NA | NA |
| No | NA | NA | NA | NA | |
| Unknown | NA | NA | NA | NA | |
| Additional pharmaceutical | Yes | 97 (11.05%) | 58 (7.58%) | NA | NA |
| Therapy | No | 117 (13.33%) | 152 (19.87) | NA | NA |
| Unknown | 664 (75.63%) | 555 (72.55%) | NA | NA | |
| Additional radiation | Yes | 97 (11.05%) | 55 (7.19%) | NA | NA |
| Therapy | No | 120 (13.67%) | 151 (19.74%) | NA | NA |
| Unknown | 661 (75.28%) | 559 (73.07%) | NA | NA | |
| Vital status | Alive | 477 (54.33%) | 405 (52.94%) | 67 (51.54%) | 207 (44.81%) |
| Dead | 274 (31.21%) | 360 (47.06) | 62 (47.69%) | 236 (51.08%) | |
| Unknown | 127 (14.46%) | 0 | 1 (0.77%) | 0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, N.; Wu, Y.; Wu, Y.; Wang, L.; Chen, J.; Wang, X.; Chard Dunmall, L.S.; Cheng, Z.; Wang, Y. Ferroptosis-Related Genes Are Potential Therapeutic Targets and the Model of These Genes Influences Overall Survival of NSCLC Patients. Cells 2022, 11, 2207. https://doi.org/10.3390/cells11142207
Zhang N, Wu Y, Wu Y, Wang L, Chen J, Wang X, Chard Dunmall LS, Cheng Z, Wang Y. Ferroptosis-Related Genes Are Potential Therapeutic Targets and the Model of These Genes Influences Overall Survival of NSCLC Patients. Cells. 2022; 11(14):2207. https://doi.org/10.3390/cells11142207
Chicago/Turabian StyleZhang, Na, Yangyang Wu, Yifan Wu, Lihong Wang, Jingfei Chen, Xiaosa Wang, Louisa S. Chard Dunmall, Zhenguo Cheng, and Yaohe Wang. 2022. "Ferroptosis-Related Genes Are Potential Therapeutic Targets and the Model of These Genes Influences Overall Survival of NSCLC Patients" Cells 11, no. 14: 2207. https://doi.org/10.3390/cells11142207