
Masculinizer and Doublesex as Key Factors Regulate Sexual Dimorphism in Ostrinia furnacalis


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Strains and Rearing
2.2. Phylogenetic Analysis
2.3. Quantitative Real-Time PCR (qRT-PCR)
2.4. In Vitro Transcription of Cas9 mRNA and sgRNA
2.5. Microinjection of Embryos
2.6. Genomic DNA Extraction and Identification of Mutagenesis
2.7. Hatchability Assay
2.8. Statistical Analysis of Data
3. Results
3.1. Phylogenetic Analysis of MASC and DSX Proteins in O. furnacalis
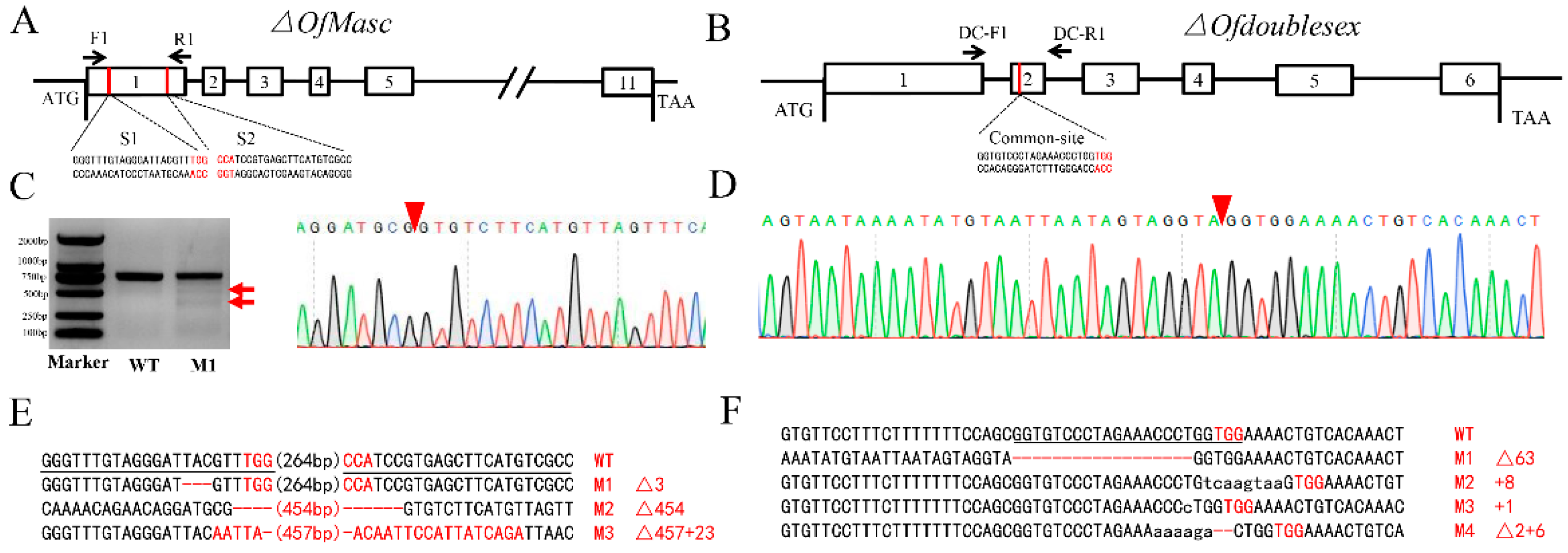
3.2. CRISPR/Cas9-Mediated Mutagenesis of OfMasc and Ofdsx
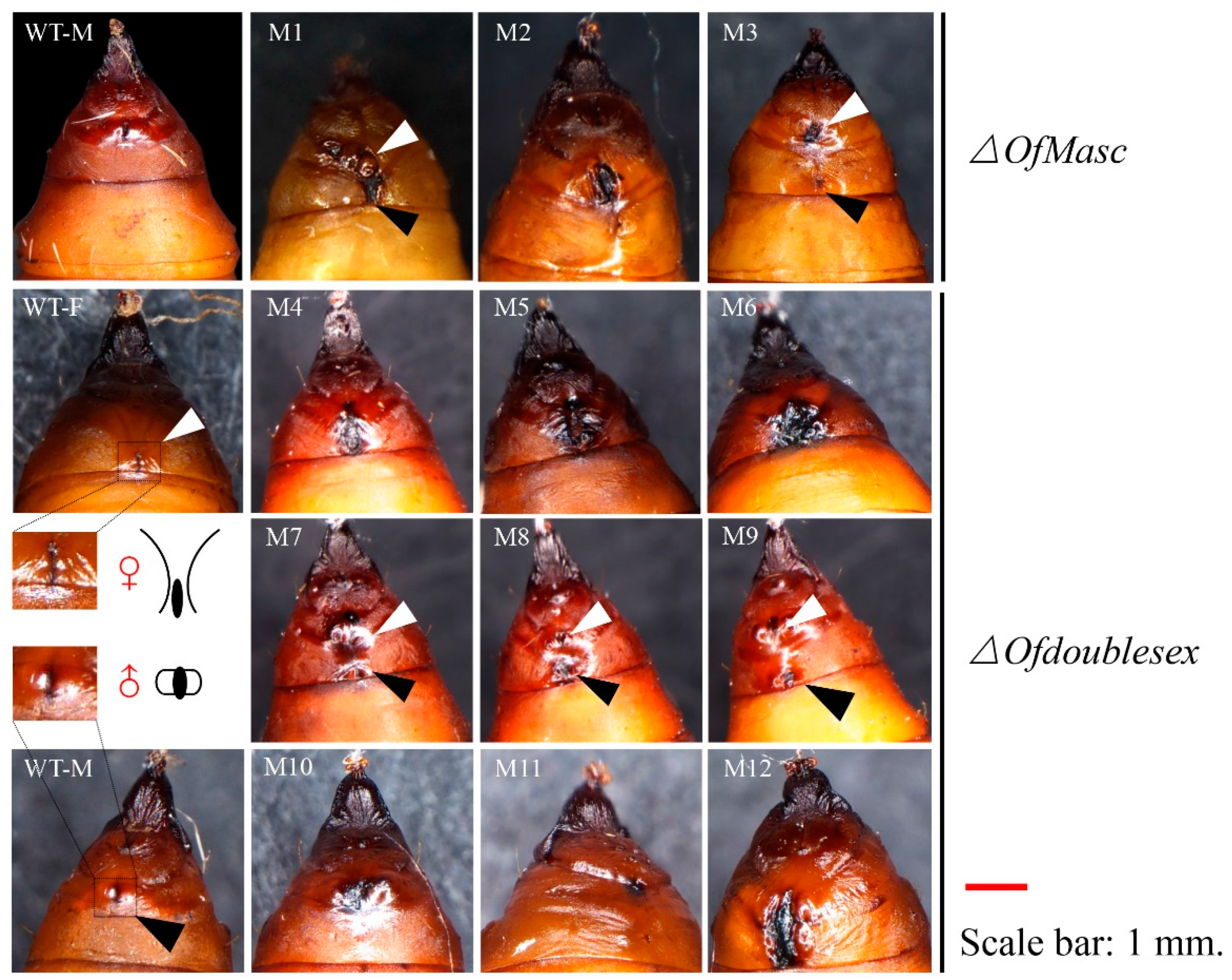
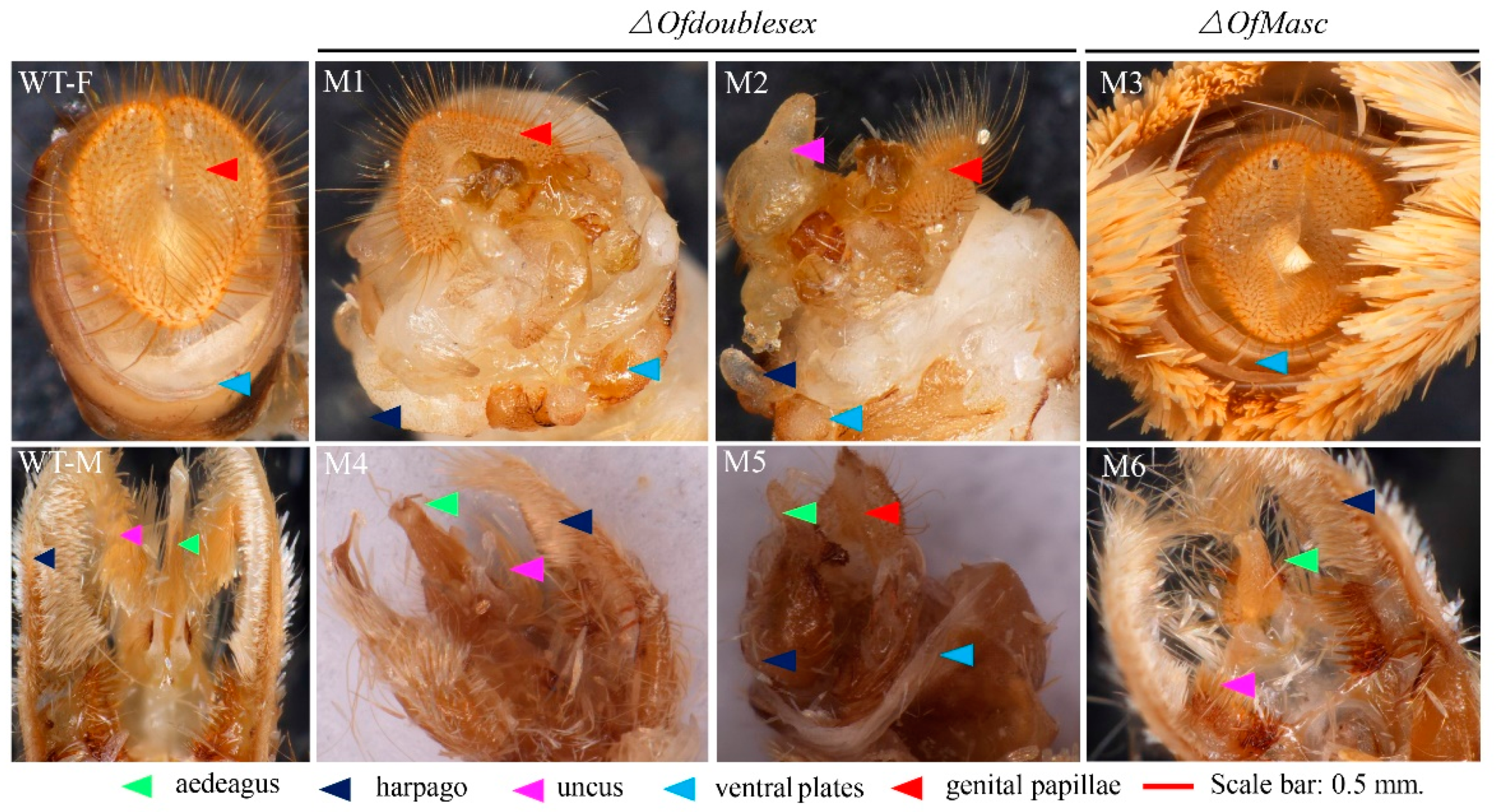
3.3. Disruption of OfMasc and Ofdsx Genes Induced Abnormal External Genitalia and Pigmentation
3.4. OfMasc and Ofdsx Mutations Induce Sterility in O. furnacalis
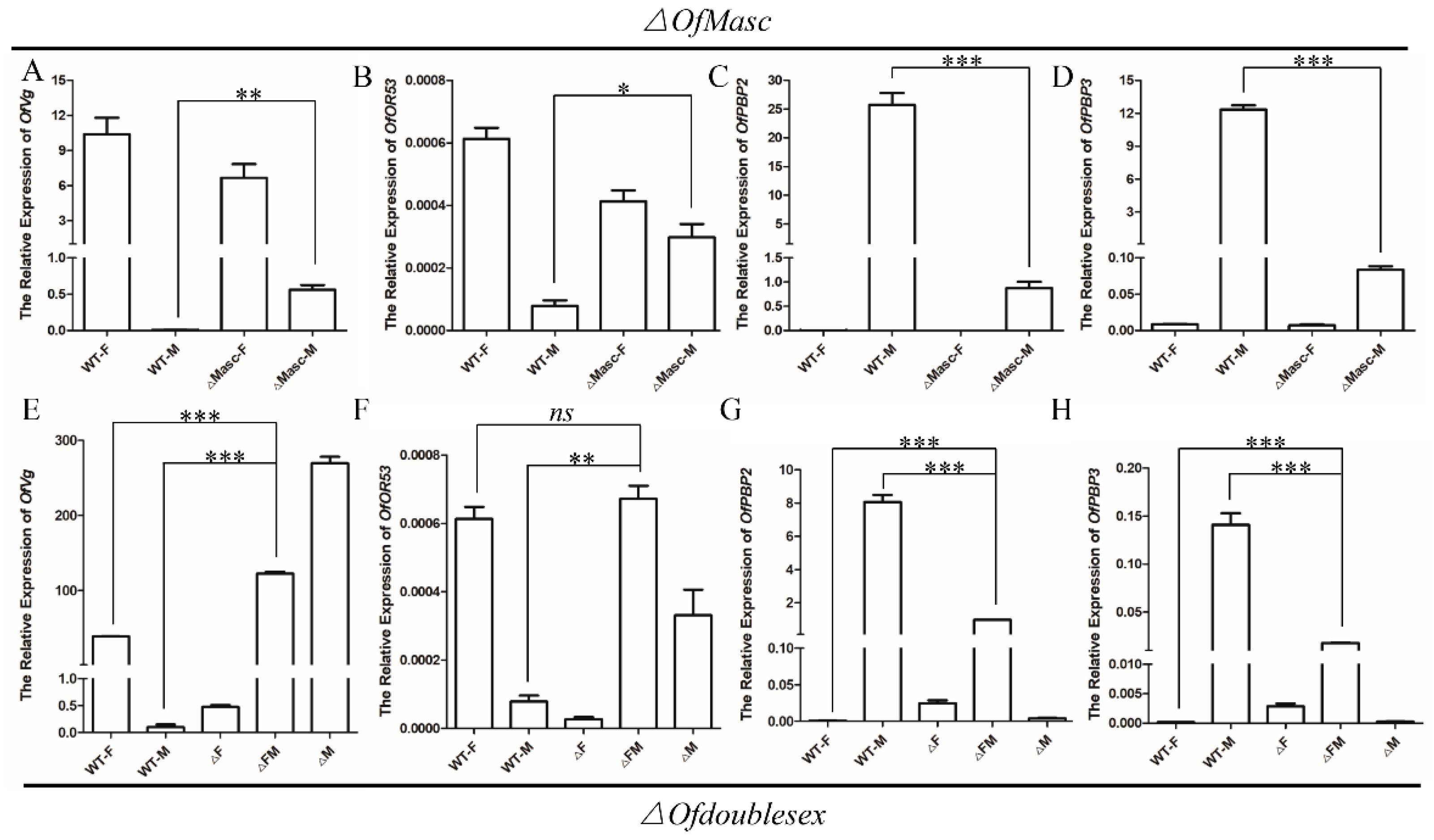
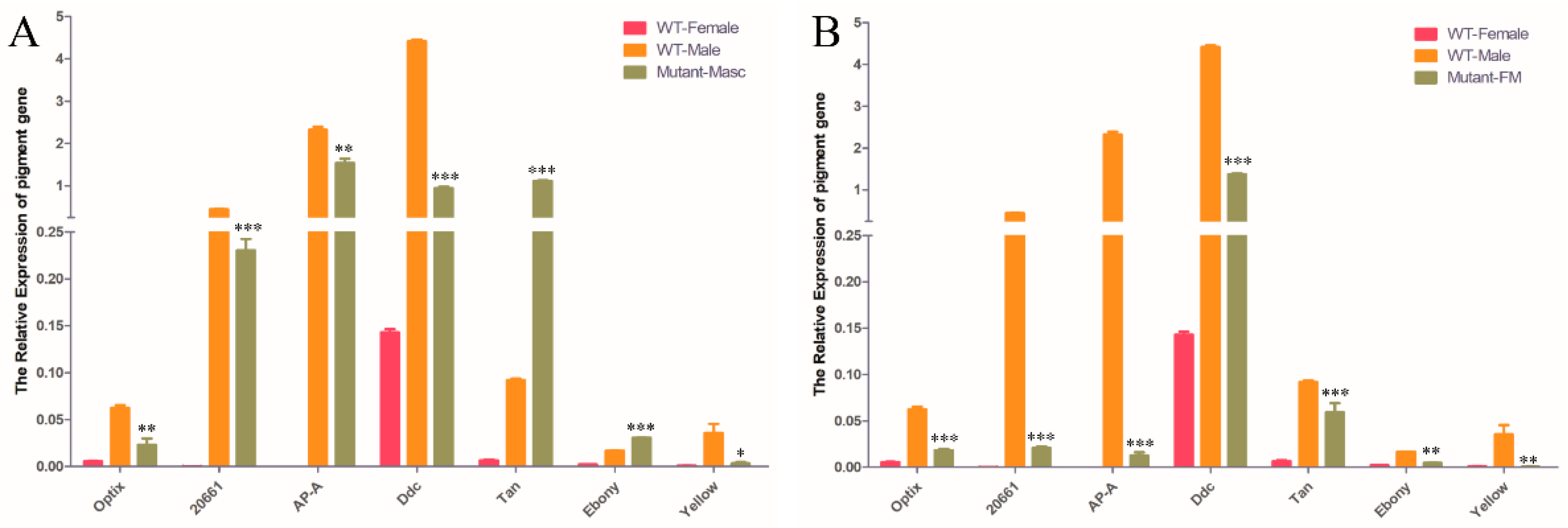
3.5. Detection of Sex-Specific Gene Expression in OfMasc and Ofdsx Mutants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hopkins, B.R.; Kopp, A. Evolution of sexual development and sexual dimorphism in insects. Curr. Opin. Genet. Dev. 2021, 69, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Parker, G.A. The sexual cascade and the rise of pre-ejaculatory (Darwinian) sexual selection, sex roles, and sexual conflict. Cold Spring Harb. Perspect. Biol. 2014, 6, a017509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mank, J.E. The transcriptional architecture of phenotypic dimorphism. Nat. Ecol. Evol. 2017, 1, 6. [Google Scholar] [CrossRef]
- Salz, H.K. Sex determination in insects: A binary decision based on alternative splicing. Curr. Opin. Genet. Dev. 2011, 21, 395–400. [Google Scholar] [CrossRef] [Green Version]
- Gempe, T.; Beye, M. Function and evolution of sex determination mechanisms, genes and pathways in insects. Bioessays 2011, 33, 52–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachtrog, D.; Mank, J.E.; Peichel, C.L.; Kirkpatrick, M.; Otto, S.P.; Ashman, T.L.; Hahn, M.W.; Kitano, J.; Mayrose, I.; Ming, R.; et al. Sex determination: Why so many ways of doing it? PLoS Biol. 2014, 12, e1001899. [Google Scholar] [CrossRef] [Green Version]
- Prakash, A.; Monteiro, A. Molecular mechanisms of secondary sexual trait development in insects. Curr. Opin. Insect Sci. 2016, 17, 40–48. [Google Scholar] [CrossRef]
- Coschigano, K.T.; Wensink, P.C. Sex-specific transcriptional regulation by the male and female doublesex proteins of Drosophila. Genes Dev. 1993, 7, 42–54. [Google Scholar] [CrossRef] [Green Version]
- Raymond, C.S.; Shamu, C.E.; Shen, M.M.; Seifert, K.J.; Hirsch, B.; Hodgkin, J.; Zarkower, D. Evidence for evolutionary conservation of sex-determining genes. Nature 1998, 391, 691–695. [Google Scholar] [CrossRef]
- Raymond, C.S.; Murphy, M.W.; O’Sullivan, M.G.; Bardwell, V.J.; Zarkower, D. Dmrt1, a gene related to worm and fly sexual regulators, is required for mammalian testis differentiation. Genes Dev. 2000, 14, 2587–2595. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, V.B.; Bachtrog, D. Evolution of sex chromosomes in insects. Annu. Rev. Genet. 2010, 44, 91–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, S.M.; Yang, S.Y.; Van Doren, M. Germ cell sex determination: A collaboration between soma and germline. Curr. Opin. Cell Biol. 2010, 22, 722–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burtis, K.C.; Baker, B.S. Drosophila doublesex gene controls somatic sexual differentiation by producing alternatively spliced mRNAs encoding related sex-specific polypeptides. Cell 1989, 56, 997–1010. [Google Scholar] [CrossRef]
- Matson, C.K.; Zarkower, D. Sex and the singular DM domain: Insights into sexual regulation, evolution and plasticity. Nat. Rev. Genet. 2012, 13, 163–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, T.M.; Selegue, J.E.; Werner, T.; Gompel, N.; Kopp, A.; Carroll, S.B. The regulation and evolution of a genetic switch controlling sexually dimorphic traits in Drosophila. Cell 2008, 134, 610–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Futahashi, R. Color vision and color formation in dragonflies. Curr. Opin. Insect Sci. 2016, 17, 32–39. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Harano, K.I.; Nishide, Y.; Sugahara, R. The mechanism controlling phenotypic plasticity of body color in the desert locust: Some recent progress. Curr. Opin. Insect Sci. 2016, 17, 10–15. [Google Scholar] [CrossRef]
- Traut, W.; Sahara, K.; Marec, F. Sex chromosomes and sex determination in Lepidoptera. Sex. Dev. 2007, 1, 332–346. [Google Scholar] [CrossRef]
- Fujii, T.; Shimada, T. Sex determination in the silkworm, Bombyx mori: A female determinant on the W chromosome and the sex-determining gene cascade. Semin. Cell Dev. Biol. 2007, 18, 379–388. [Google Scholar] [CrossRef]
- Nagaraju, J.; Gopinath, G.; Sharma, V.; Shukla, J.N. Lepidopteran sex determination: A cascade of surprises. Sex. Dev. 2014, 8, 104–112. [Google Scholar] [CrossRef]
- Kiuchi, T.; Koga, H.; Kawamoto, M.; Shoji, K.; Sakai, H.; Arai, Y.; Ishihara, G.; Kawaoka, S.; Sugano, S.; Shimada, T.; et al. A single female-specific piRNA is the primary determiner of sex in the silkworm. Nature 2014, 509, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Katsuma, S.; Sugano, Y.; Kiuchi, T.; Shimada, T. Two conserved cysteine residues are required for the masculinizing activity of the silkworm Masc protein. J. Biol. Chem. 2015, 290, 26114–26124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiuchi, T.; Sugano, Y.; Shimada, T.; Katsuma, S. Two CCCH-type zinc finger domains in the Masc protein are dispensable for masculinization and dosage compensation in Bombyx mori. Insect Biochem. Mol. Biol. 2019, 104, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Chen, S.; Zeng, B.; James, A.A.; Tan, A.; Huang, Y. Bombyx mori P-element Somatic Inhibitor (BmPSI) is a key auxiliary factor for silkworm male sex determination. PLoS Genet. 2017, 13, e1006576. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Wang, Y.; Li, Z.; Ling, L.; Zeng, B.; James, A.A.; Tan, A.; Huang, Y. Transcription activator-like effector nuclease (TALEN)-mediated female-specific sterility in the silkworm, Bombyx mori. Insect Mol. Biol. 2014, 23, 800–807. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Yan, S.; Huang, Y.; Tan, A.; Stanley, D.W.; Song, Q. Genetic transformation mediated by piggyBac in the Asian corn borer, Ostrinia furnacalis (Lepidoptera: Crambidae). Arch. Insect Biochem. Physiol. 2012, 80, 140–150. [Google Scholar] [CrossRef]
- Liu, Q.; Hallerman, E.; Peng, Y.; Li, Y. Development of Bt rice and Bt maize in China and their efficacy in target pest control. Int. J. Mol. Sci. 2016, 17, 1561. [Google Scholar] [CrossRef] [Green Version]
- Kageyama, D.; Nishimura, G.; Hoshizaki, S.; Ishikawa, Y. Feminizing Wolbachia in an insect, Ostrinia furnacalis (Lepidoptera: Crambidae). Heredity 2002, 88, 444–449. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Y.; Zheng, Z.Z.; Song, H.S.; Xu, Y.Z. Conserved RNA cis-elements regulate alternative splicing of Lepidopteran doublesex. Insect Biochem. Mol. Biol. 2014, 44, 1–11. [Google Scholar] [CrossRef]
- Fukui, T.; Kawamoto, M.; Shoji, K.; Kiuchi, T.; Sugano, S.; Shimada, T.; Suzuki, Y.; Katsuma, S. The endosymbiotic bacterium Wolbachia selectively kills male hosts by targeting the Masculinizing gene. PLoS Pathog. 2015, 11, e1005048. [Google Scholar] [CrossRef]
- Fukui, T.; Kiuchi, T.; Shoji, K.; Kawamoto, M.; Shimada, T.; Katsuma, S. In vivo masculinizing function of the Ostrinia furnacalis Masculinizer gene. Biochem. Biophys. Res. Commun. 2018, 503, 1768–1772. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Ozaki, K.; Ishikawa, Y.; Matsuo, T. Identification of candidate odorant receptors in Asian corn borer Ostrinia furnacalis. PLoS ONE 2015, 10, e0121261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Li, Z.; Xu, J.; Zeng, B.; Ling, L.; You, L.; Chen, Y.; Huang, Y.; Tan, A. The CRISPR/Cas system mediates efficient genome engineering in Bombyx mori. Cell Res. 2013, 23, 1414–1416. [Google Scholar] [CrossRef] [Green Version]
- Harvey-Samuel, T.; Norman, V.C.; Carter, R.; Lovett, E.; Alphey, L. Identification and characterisation of a Masculinizer homolog in the diamondback moth Plutella xylostella. Insect Mol. Biol. 2019, 29, 231–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, L.; Bi, H.L.; Wang, Y.H.; Li, X.W.; Chen, X.E.; Li, Z.Q. CRISPR/Cas9-based mutation reveals Argonaute 1 is essential for pigmentation in Ostrinia furnacalis. Insect Sci. 2019, 26, 1020–1028. [Google Scholar] [CrossRef]
- Kondo, S.; Ueda, R. Highly improved gene targeting by germline-specific Cas9 expression in Drosophila. Genetics 2013, 195, 715–721. [Google Scholar] [CrossRef] [Green Version]
- Bi, H.L.; Xu, J.; Tan, A.J.; Huang, Y.P. CRISPR/Cas9-mediated targeted gene mutagenesis in Spodoptera litura. Insect Sci. 2016, 23, 469–477. [Google Scholar] [CrossRef]
- Xu, J.; Zhan, S.; Chen, S.; Zeng, B.; Li, Z.; James, A.A.; Tan, A.; Huang, Y. Sexually dimorphic traits in the silkworm, Bombyx mori, are regulated by doublesex. Insect Biochem. Mol. Biol. 2017, 80, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, X.; Liu, Z.; Xu, J.; Li, X.; Bi, H.; Andongma, A.A.; Niu, C.; Huang, Y. Mutation of doublesex induces sex-specific sterility of the diamondback moth Plutella xylostella. Insect Biochem. Mol. Biol. 2019, 112, 103180. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, Q.; Liu, H.; Bi, H.; Wang, Y.; Chen, X.; Wu, N.; Xu, J.; Zhang, Z.; Huang, Y.; et al. Mutation of doublesex in Hyphantria cunea results in sex-specific sterility. Pest Manag. Sci. 2019, 76, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kiuchi, T.; Kawamoto, M.; Shimada, T.; Katsuma, S. Identification and functional analysis of a Masculinizer orthologue in Trilocha varians (Lepidoptera: Bombycidae). Insect Mol. Biol. 2015, 24, 561–569. [Google Scholar] [CrossRef]
- Wang, Y.H.; Chen, X.E.; Yang, Y.; Xu, J.; Fang, G.Q.; Niu, C.Y.; Huang, Y.P.; Zhan, S. The Masc gene product controls masculinization in the black cutworm, Agrotis ipsilon. Insect Sci. 2019, 26, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.M.; Carroll, S.B. Genetic and molecular insights into the development and evolution of sexual dimorphism. Nat. Rev. Genet. 2009, 10, 797–804. [Google Scholar] [CrossRef]
- Tan, A.; Fu, G.; Jin, L.; Guo, Q.; Li, Z.; Niu, B.; Meng, Z.; Morrison, N.I.; Alphey, L.; Huang, Y. Transgene-based, female-specific lethality system for genetic sexing of the silkworm, Bombyx mori. Proc. Natl. Acad. Sci. USA 2013, 110, 6766–6770. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, T.N.; Fujii, T.; Kayukawa, T.; Sakamoto, H.; Ishikawa, Y. Expression of a doublesex homologue is altered in sexual mosaics of Ostrinia scapulalis moths infected with Wolbachia. Insect Biochem. Mol. Biol. 2010, 40, 847–854. [Google Scholar] [CrossRef]
- Chen, X.; Cao, Y.; Zhan, S.; Tan, A.; Palli, S.R.; Huang, Y. Disruption of sex-specific doublesex exons results in male- and female-specific defects in the black cutworm, Agrotis ipsilon. Pest Manag. Sci. 2019, 75, 1697–1706. [Google Scholar] [CrossRef]
- Kunte, K.; Zhang, W.; Tenger-Trolander, A.; Palmer, D.H.; Martin, A.; Reed, R.D.; Mullen, S.P.; Kronforst, M.R. doublesex is a mimicry supergene. Nature 2014, 507, 229–232. [Google Scholar] [CrossRef]
- Nishikawa, H.; Iijima, T.; Kajitani, R.; Yamaguchi, J.; Ando, T.; Suzuki, Y.; Sugano, S.; Fujiyama, A.; Kosugi, S.; Hirakawa, H.; et al. A genetic mechanism for female-limited Batesian mimicry in Papilio butterfly. Nat. Genet. 2015, 47, 405–409. [Google Scholar] [CrossRef]
- Clarke, C.A.; Sheppard, P.M. Super-genes and mimicry. Heredity 1960, 14, 175–185. [Google Scholar] [CrossRef]
- Loehlin, D.W.; Carroll, S.B. Evolutionary biology: Sex, lies and butterflies. Nature 2014, 507, 172–173. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Weight (g) or Volume (mL) |
|---|---|
| Wheat germ powder | 150 |
| Yeast extract powder | 40 |
| Agar strip | 14 |
| Sucrose | 5 |
| Vitamin C | 4 |
| Sorbic acid | 4 |
| Methyl p-hydroxybenzoate | 4 |
| Linoleic acid | 0.5 |
| purified water | 900 |
| Primer Name | Primer Sequence (5′-3′) | Primer Purpose |
|---|---|---|
| Masc-sgF1 | TAATACGACTCACTATAGGGTTTGTAGGGATTACGTTGTTTTAGAGCTAGAAATAGCAA | Preparation of sgRNA templates |
| Masc-sgF2 | TAATACGACTCACTATAGGCGACATGAAGCTCACGGAGTTTTAGAGCTAGAAATAGCAA | |
| Dsx-sgF1 | TAATACGACTCACTATAGGTGTCCCTAGAAACCCTGGGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAG | |
| sgRNA-R | AAAAGCACCGACTCGGTGCCACTTTTTCAAGTTGATAACGGACTAGCCTTATTTTAACTTGCTATTTCTAGCTCTAAAAC | |
| Masc-F1 | ACATAGTGAACAAAATGGCCGCCAC | Identification of somatic mutations |
| Masc-R1 | TTGAGGTGGTGGTGCTGAAACAGAA | |
| Dsx-DC-F1 | AAACGCTTTTATTTAGAGGTTAAGAGGG | |
| Dsx-DC-R1 | GCTGAAATGATGATGATGATCCAAA | |
| Dsx-RTPCR-F | AAGTTCCACTATTCCTGGGAG | qRT-PCR for genes |
| Dsx-RTPCR-R | AGCACATCGAGTACGAGGAG | |
| Actin-qF | CCGTCCTCCTGACCGAGGCTC | |
| Actin-qR | GGTGTGGGAGACACCATCTCCG | |
| Vg-qF | TCTTACAAATCGCGCAATGG | |
| Vg-qR | GACTTGGAGACGTTCTTGAC | |
| OR53-qF | GGAGCTATTACCTACGTGAAGC | |
| OR53-qR | TTAAGCGCAGGCTGCGTTCATG | |
| PBP2-qF | ATGTGCTCGATGAGCGTTGT | |
| PBP2-qR | CTTGGATGAAAGGCAGAGGAT | |
| PBP3-qF | AAGACGCTTGTGGTGATGGCA | |
| PBP3-qR | GATCAGTTGTAATCCTGTGGC | |
| Optix-qF | GCCCATTATCAGGAAGCAGA | |
| Optix-qR | CAGCTCCCTCTTCTTTGTCG | |
| 206617-qF | ATGGATACGAGGCACAAAGC | |
| 206617-qR | GAGGATCAGTGTGCAAAGCA | |
| APA-qF | TATGGCGGTACGACACTTTG | |
| APA-qR | GGAAGGCAGTCCGTCTTGTA | |
| Ddc-qF | TTGGTTCGTCTTGAGGCTTT | |
| Ddc-qR | CCATTAATGCGCTTCAACAA | |
| Tan-qF | TCATCGCGACGTATGCTAAC | |
| Tan-qR | ATGGTTCCAATGAGGTCGTC | |
| Ebony-qF | CGTCTGCCCTATTCAGCAAT | |
| Ebony-qR | CACCAGCTTCTGAGGGTCTC | |
| Yellow-qF | TGTTGGAATTCCGCTCTTTC | |
| Yellow-qR | ACGGGACCGTGTAAATTCTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bi, H.; Li, X.; Xu, X.; Wang, Y.; Zhou, S.; Huang, Y. Masculinizer and Doublesex as Key Factors Regulate Sexual Dimorphism in Ostrinia furnacalis. Cells 2022, 11, 2161. https://doi.org/10.3390/cells11142161
Bi H, Li X, Xu X, Wang Y, Zhou S, Huang Y. Masculinizer and Doublesex as Key Factors Regulate Sexual Dimorphism in Ostrinia furnacalis. Cells. 2022; 11(14):2161. https://doi.org/10.3390/cells11142161
Chicago/Turabian StyleBi, Honglun, Xiaowei Li, Xia Xu, Yaohui Wang, Shutang Zhou, and Yongping Huang. 2022. "Masculinizer and Doublesex as Key Factors Regulate Sexual Dimorphism in Ostrinia furnacalis" Cells 11, no. 14: 2161. https://doi.org/10.3390/cells11142161