
P-Cadherin Is Expressed by Epithelial Progenitor Cells and Melanocytes in the Human Corneal Limbus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Isolation
2.2. Fluorescence-Activated Cell Sorting (FACS)
2.3. CFU-E Assay
2.4. Co-cultures
2.5. Real-time RT-PCR
2.6. Flow Cytometry
2.7. Immunohistochemistry of Frozen Sections and Immunocytochemistry
2.8. Histology and Immunohistochemistry—Paraffin
2.9. Recellularization of Decellularized Human Limbal Tissue
2.10. Statistical Analysis
3. Results
3.1. Localization of P-cad at the Corneal Limbus
3.2. Flow Sorting and Characterization of Limbal Cells
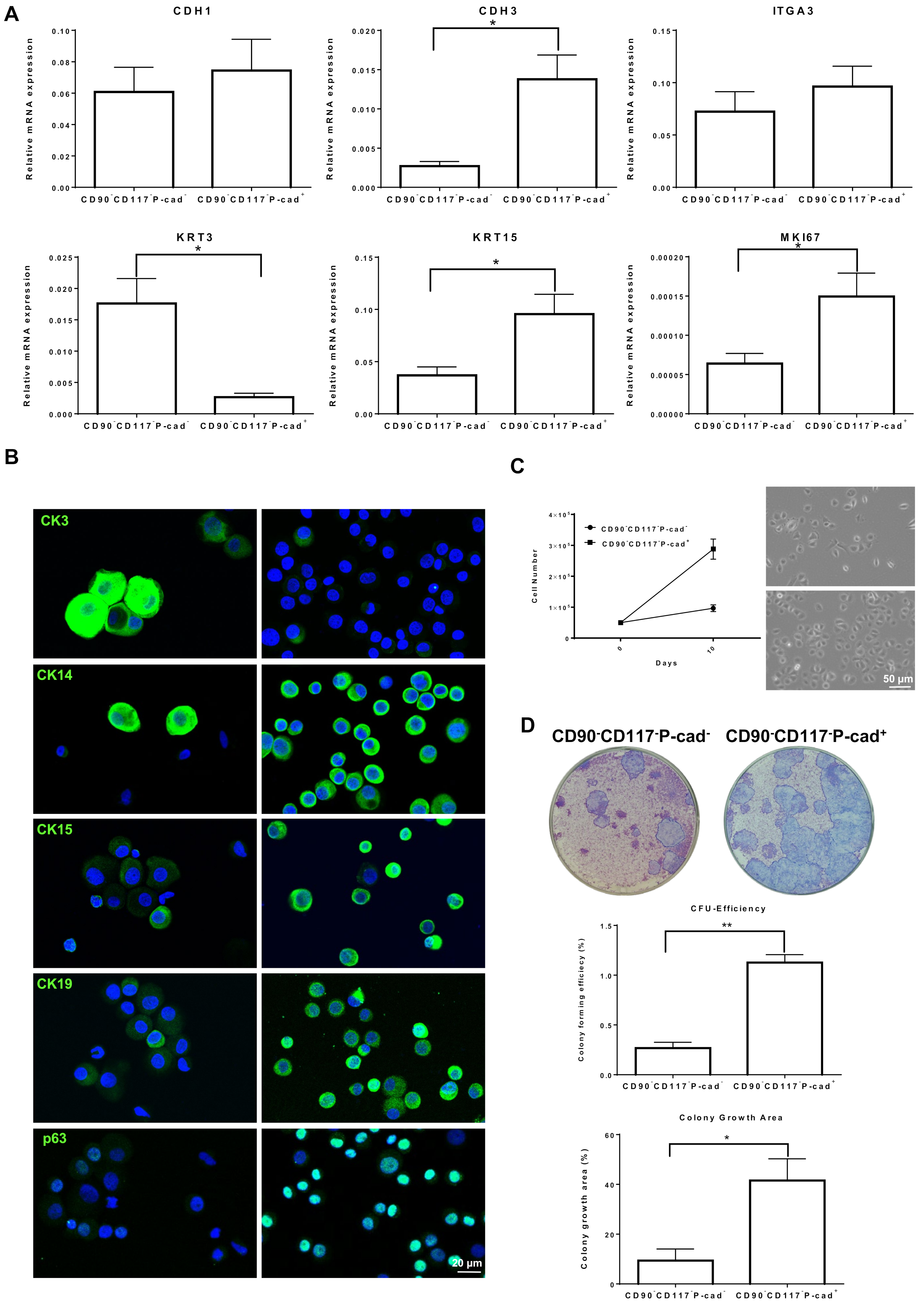
3.3. Progenitor Cell Properties of CD90−CD117−P-cad+ and CD90−CD117−P-cad− Limbal Epithelial Cells
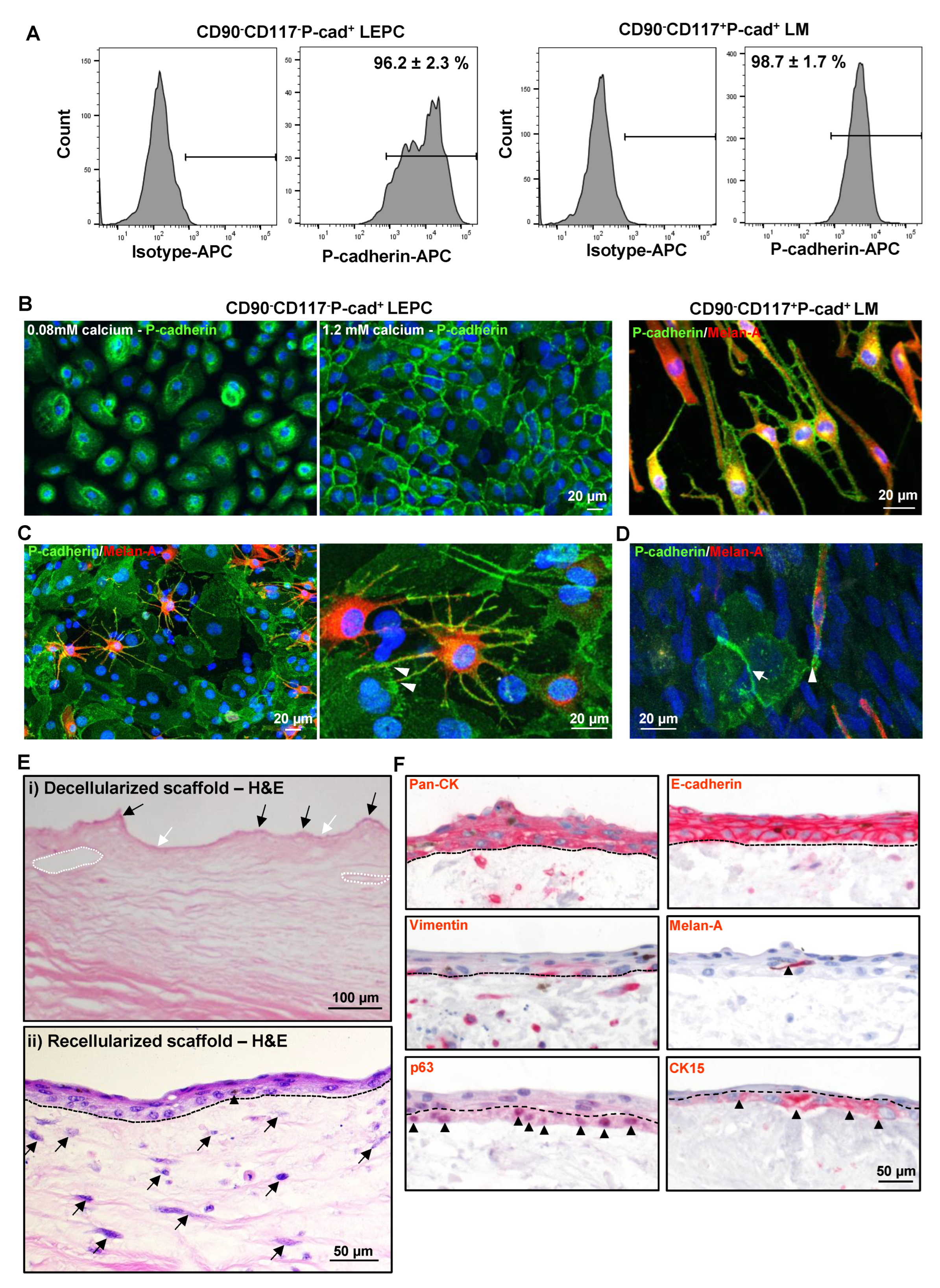
3.4. P-cad Is Associated with LEPC and LM
3.5. Repopulation of Decellularized Limbal Scaffolds
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cotsarelis, G.; Cheng, S.-Z.; Dong, G.; Sun, T.-T.; Lavker, R.M. Existence of slow-cycling limbal epithelial basal cells that can be preferentially stimulated to proliferate: Implications on epithelial stem cells. Cell 1989, 57, 201–209. [Google Scholar] [CrossRef]
- Goldberg, M.F.; Bron, A.J. Limbal Palisades of Vogt. Trans. Am. Ophthalmol. Soc. 1982, 80, 155–171. [Google Scholar] [PubMed]
- Shortt, A.J.; Secker, G.A.; Munro, P.M.; Khaw, P.T.; Tuft, S.J.; Daniels, J.T. Characterization of the Limbal Epithelial Stem Cell Niche: Novel Imaging Techniques Permit In Vivo Observation and Targeted Biopsy of Limbal Epithelial Stem Cells. Stem Cells 2007, 25, 1402–1409. [Google Scholar] [CrossRef] [PubMed]
- Dziasko, M.A.; Tuft, S.J.; Daniels, J.T. Limbal melanocytes support limbal epithelial stem cells in 2D and 3D microenvironments. Exp. Eye Res. 2015, 138, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Tseng, S.C.; He, H.; Zhang, S.; Chen, S.-Y. Niche Regulation of Limbal Epithelial Stem Cells: Relationship between Inflammation and Regeneration. Ocul. Surf. 2016, 14, 100–112. [Google Scholar] [CrossRef] [Green Version]
- Abdul-Al, M.; Kyeremeh, G.; Saeinasab, M.; Keshel, S.H.; Sefat, F. Stem Cell Niche Microenvironment: Review. Bioengineering 2021, 8, 108. [Google Scholar] [CrossRef]
- Haug, J.S.; He, X.C.; Grindley, J.C.; Wunderlich, J.P.; Gaudenz, K.; Ross, J.T.; Paulson, A.; Wagner, K.P.; Xie, Y.; Zhu, R.; et al. N-Cadherin Expression Level Distinguishes Reserved versus Primed States of Hematopoietic Stem Cells. Cell Stem Cell 2008, 2, 367–379. [Google Scholar] [CrossRef] [Green Version]
- Karpowicz, P.; Willaime-Morawek, S.; Balenci, L.; DeVeale, B.; Inoue, T.; Van Der Kooy, D. E-Cadherin Regulates Neural Stem Cell Self-Renewal. J. Neurosci. 2009, 29, 3885–3896. [Google Scholar] [CrossRef]
- Schlötzer-Schrehardt, U.; Kruse, F.E. Identification and characterization of limbal stem cells. Exp. Eye Res. 2005, 81, 247–264. [Google Scholar] [CrossRef]
- Hayashi, R.; Yamato, M.; Sugiyama, H.; Sumide, T.; Yang, J.; Okano, T.; Tano, Y.; Nishida, K. N-Cadherin Is Expressed by Putative Stem/Progenitor Cells and Melanocytes in the Human Limbal Epithelial Stem Cell Niche. Stem Cells 2006, 25, 289–296. [Google Scholar] [CrossRef]
- Higa, K.; Shimmura, S.; Miyashita, H.; Kato, N.; Ogawa, Y.; Kawakita, T.; Shimazaki, J.; Tsubota, K. N-Cadherin in the Maintenance of Human Corneal Limbal Epithelial Progenitor Cells In Vitro. Investig. Opthalmology Vis. Sci. 2009, 50, 4640–4645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polisetti, N.; Zenkel, M.; Menzel-Severing, J.; Kruse, F.E.; Schlötzer-Schrehardt, U. Cell Adhesion Molecules and Stem Cell-Niche-Interactions in the Limbal Stem Cell Niche. Stem Cells 2015, 34, 203–219. [Google Scholar] [CrossRef] [PubMed]
- Raymond, K.; Deugnier, M.-A.; Faraldo, M.M.A.; Glukhova, M. Adhesion within the stem cell niches. Curr. Opin. Cell Biol. 2009, 21, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Larue, L.; Antos, C.; Butz, S.; Huber, O.; Delmas, V.; Dominis, M.; Kemler, R. A role for cadherins in tissue formation. Development 1996, 122, 3185–3194. [Google Scholar] [CrossRef] [PubMed]
- Tepass, U.; Truong, K.; Godt, D.; Ikura, M.; Peifer, M. Cadherins in embryonic and neural morphogenesis. Nat. Rev. Mol. Cell Biol. 2000, 1, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Kolle, G.; Ho, M.; Zhou, Q.; Chy, H.S.; Krishnan, K.; Cloonan, N.; Bertoncello, I.; Laslett, A.L.; Grimmond, S.M. Identification of Human Embryonic Stem Cell Surface Markers by Combined Membrane-Polysome Translation State Array Analysis and Immunotranscriptional Profiling. Stem Cells 2009, 27, 2446–2456. [Google Scholar] [CrossRef]
- Nguyen, H.P.T.; Xiao, L.; Deane, J.A.; Tan, K.-S.; Cousins, F.L.; Masuda, H.; Sprung, C.N.; Rosamilia, A.; Gargett, C.E. N-cadherin identifies human endometrial epithelial progenitor cells by in vitro stem cell assays. Hum. Reprod. 2017, 32, 2254–2268. [Google Scholar] [CrossRef] [Green Version]
- Figueira, E.C.; Di Girolamo, N.; Coroneo, M.T.; Wakefield, D. The Phenotype of Limbal Epithelial Stem Cells. Investig. Opthalmology Vis. Sci. 2007, 48, 144–156. [Google Scholar] [CrossRef] [Green Version]
- Martignani, E.; Ala, U.; Sheehy, P.A.; Thomson, P.C.; Baratta, M. Whole transcriptome analysis of bovine mammary progenitor cells by P-Cadherin enrichment as a marker in the mammary cell hierarchy. Sci. Rep. 2020, 10, 14183. [Google Scholar] [CrossRef]
- Hayashi, R.; Yamato, M.; Saito, T.; Oshima, T.; Okano, T.; Tano, Y.; Nishida, K. Enrichment of corneal epithelial stem/progenitor cells using cell surface markers, integrin α6 and CD71. Biochem. Biophys. Res. Commun. 2008, 367, 256–263. [Google Scholar] [CrossRef]
- Truong, T.T.; Huynh, K.; Nakatsu, M.N.; Deng, S.X. SSEA4 Is a Potential Negative Marker for the Enrichment of Human Corneal Epithelial Stem/Progenitor Cells. Investig. Opthalmology Vis. Sci. 2011, 52, 6315–6320. [Google Scholar] [CrossRef] [PubMed]
- Shaharuddin, B.; Harvey, I.; Ahmad, S.; Ali, S.; Meeson, A. Characterisation of Human Limbal Side Population Cells Isolated Using an Optimised Protocol from an Immortalised Epithelial Cell Line and Primary Limbal Cultures. Stem Cell Rev. Rep. 2013, 10, 240–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bojic, S.; Hallam, D.; Alcada, N.; Ghareeb, A.; Queen, R.; Pervinder, S.; Buck, H.; Lange, A.A.; Figueiredo, G.; Rooney, P.; et al. CD200 Expression Marks a Population of Quiescent Limbal Epithelial Stem Cells with Holoclone Forming Ability. Stem Cells 2018, 36, 1723–1735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norrick, A.; Esterlechner, J.; Niebergall-Roth, E.; Dehio, U.; Sadeghi, S.; Schröder, H.M.; Ballikaya, S.; Stemler, N.; Ganss, C.; Dieter, K.; et al. Process development and safety evaluation of ABCB5+ limbal stem cells as advanced-therapy medicinal product to treat limbal stem cell deficiency. Stem Cell Res. Ther. 2021, 12, 1–21. [Google Scholar] [CrossRef]
- Polisetti, N.; Sharaf, L.; Schlötzer-Schrehardt, U.; Schlunck, G.; Reinhard, T. Efficient Isolation and Functional Characterization of Niche Cells from Human Corneal Limbus. Int. J. Mol. Sci. 2022, 23, 2750. [Google Scholar] [CrossRef]
- Polisetti, N.; Schlötzer-Schrehardt, U.; Reinhard, T.; Schlunck, G. Isolation and enrichment of melanocytes from human corneal limbus using CD117 (c-Kit) as selection marker. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Polisetti, N.; Gießl, A.; Zenkel, M.; Heger, L.; Dudziak, D.; Naschberger, E.; Stich, L.; Steinkasserer, A.; Kruse, F.E.; Schlötzer-Schrehardt, U. Melanocytes as emerging key players in niche regulation of limbal epithelial stem cells. Ocul. Surf. 2021, 22, 172–189. [Google Scholar] [CrossRef]
- Polisetti, N.; Sorokin, L.; Okumura, N.; Koizumi, N.; Kinoshita, S.; Kruse, F.E.; Schlötzer-Schrehardt, U. Laminin-511 and -521-based matrices for efficient ex vivo-expansion of human limbal epithelial progenitor cells. Sci. Rep. 2017, 7, e3754. [Google Scholar] [CrossRef]
- Polisetti, N.; Roschinski, B.; Schlötzer-Schrehardt, U.; Maier, P.; Schlunck, G.; Reinhard, T. A Decellularized Human Limbal Scaffold for Limbal Stem Cell Niche Reconstruction. Int. J. Mol. Sci. 2021, 22, 10067. [Google Scholar] [CrossRef]
- Lange, C.; Wolf, J.; Auw-Haedrich, C.; Schlecht, A.; Boneva, S.; Lapp, T.; Horres, R.; Agostini, H.; Martin, G.; Reinhard, T.; et al. Expression of the COVID-19 receptor ACE2 in the human conjunctiva. J. Med. Virol. 2020, 92, 2081–2086. [Google Scholar] [CrossRef]
- Molès, J.-P.; Watt, F.M. The Epidermal Stem Cell Compartment: Variation in Expression Levels of E–Cadherin and Catenins Within the Basal Layer of Human Epidermis. J. Histochem. Cytochem. 1997, 45, 867–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tinkle, C.L.; Pasolli, H.A.; Stokes, N.; Fuchs, E. New insights into cadherin function in epidermal sheet formation and maintenance of tissue integrity. Proc. Natl. Acad. Sci. USA 2008, 105, 15405–15410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armitage, W.J.; Jones, M.N.A.; Zambrano, I.; Carley, F.; Tole, D.M. The Suitability of Corneas Stored by Organ Culture for Penetrating Keratoplasty and Influence of Donor and Recipient Factors on 5-Year Graft Survival. Investig. Opthalmology Vis. Sci. 2014, 55, 784–791. [Google Scholar] [CrossRef] [PubMed]
- Polisetti, N.; Gießl, A.; Li, S.; Sorokin, L.; Kruse, F.E.; Schlötzer-Schrehardt, U. Laminin-511-E8 promotes efficient in vitro expansion of human limbal melanocytes. Sci. Rep. 2020, 10, 11074. [Google Scholar] [CrossRef]
- Rhee, H.; Polak, L.; Fuchs, E. Lhx2 Maintains Stem Cell Character in Hair Follicles. Science 2006, 312, 1946–1949. [Google Scholar] [CrossRef] [Green Version]
- Nose, A.; Takeichi, M. A novel cadherin cell adhesion molecule: Its expression patterns associated with implantation and organogenesis of mouse embryos. J. Cell Biol. 1986, 103, 2649–2658. Available online: https://pubmed.ncbi.nlm.nih.gov/3539943/ (accessed on 8 May 2022). [CrossRef] [Green Version]
- Figueiredo, F.C.; Glanville, J.M.; Arber, M.; Carr, E.; Rydevik, G.; Hogg, J.; Okonkwo, A.; Figueiredo, G.; Lako, M.; Whiter, F.; et al. A systematic review of cellular therapies for the treatment of limbal stem cell deficiency affecting one or both eyes. Ocul. Surf. 2021, 20, 48–61. [Google Scholar] [CrossRef]
- Schlereth, S.L.; Hos, D.; Matthaei, M.; Hamrah, P.; Schmetterer, L.; O’Leary, O.; Ullmer, C.; Horstmann, J.; Bock, F.; Wacker, K.; et al. New Technologies in Clinical Trials in Corneal Diseases and Limbal Stem Cell Deficiency—Review from the European Vision Institute (EVI) Special Interest Focus Group Meeting. Ophthalmic Res. 2021, 64, 145–167. [Google Scholar] [CrossRef]
- Badylak, S.F. The extracellular matrix as a scaffold for tissue reconstruction. Semin. Cell Dev. Biol. 2002, 13, 377–383. [Google Scholar] [CrossRef]
- Fernández-Pérez, J.; Ahearne, M. Decellularization and recellularization of cornea: Progress towards a donor alternative. Methods 2019, 171, 86–96. [Google Scholar] [CrossRef]
- Polisetti, N.; Schmid, A.; Schlötzer-Schrehardt, U.; Maier, P.; Lang, S.J.; Steinberg, T.; Schlunck, G.; Reinhard, T. A decellularized human corneal scaffold for anterior corneal surface reconstruction. Sci. Rep. 2021, 11, 2992. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polisetti, N.; Sharaf, L.; Martin, G.; Schlunck, G.; Reinhard, T. P-Cadherin Is Expressed by Epithelial Progenitor Cells and Melanocytes in the Human Corneal Limbus. Cells 2022, 11, 1975. https://doi.org/10.3390/cells11121975
Polisetti N, Sharaf L, Martin G, Schlunck G, Reinhard T. P-Cadherin Is Expressed by Epithelial Progenitor Cells and Melanocytes in the Human Corneal Limbus. Cells. 2022; 11(12):1975. https://doi.org/10.3390/cells11121975
Chicago/Turabian StylePolisetti, Naresh, Lyne Sharaf, Gottfried Martin, Günther Schlunck, and Thomas Reinhard. 2022. "P-Cadherin Is Expressed by Epithelial Progenitor Cells and Melanocytes in the Human Corneal Limbus" Cells 11, no. 12: 1975. https://doi.org/10.3390/cells11121975