
Transcriptional Profiling of Insulin-like Growth Factor Signaling Components in Embryonic Lung Development and Idiopathic Pulmonary Fibrosis

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. Human-Derived Lung Material
2.3. Primary Culture of Murine Lung Fibroblasts
2.4. Primary Culture of Human Lung Fibroblasts
2.5. RNA Extraction, cDNA Synthesis and qPCR
2.6. Histology, Immunohistochemistry and Fluorescent Staining
2.7. Western Blotting
2.8. Figure Assembly and Statistical Analysis
3. Results
3.1. Transcriptional Profile of IGF Signaling Components during Embryonic Lung Development
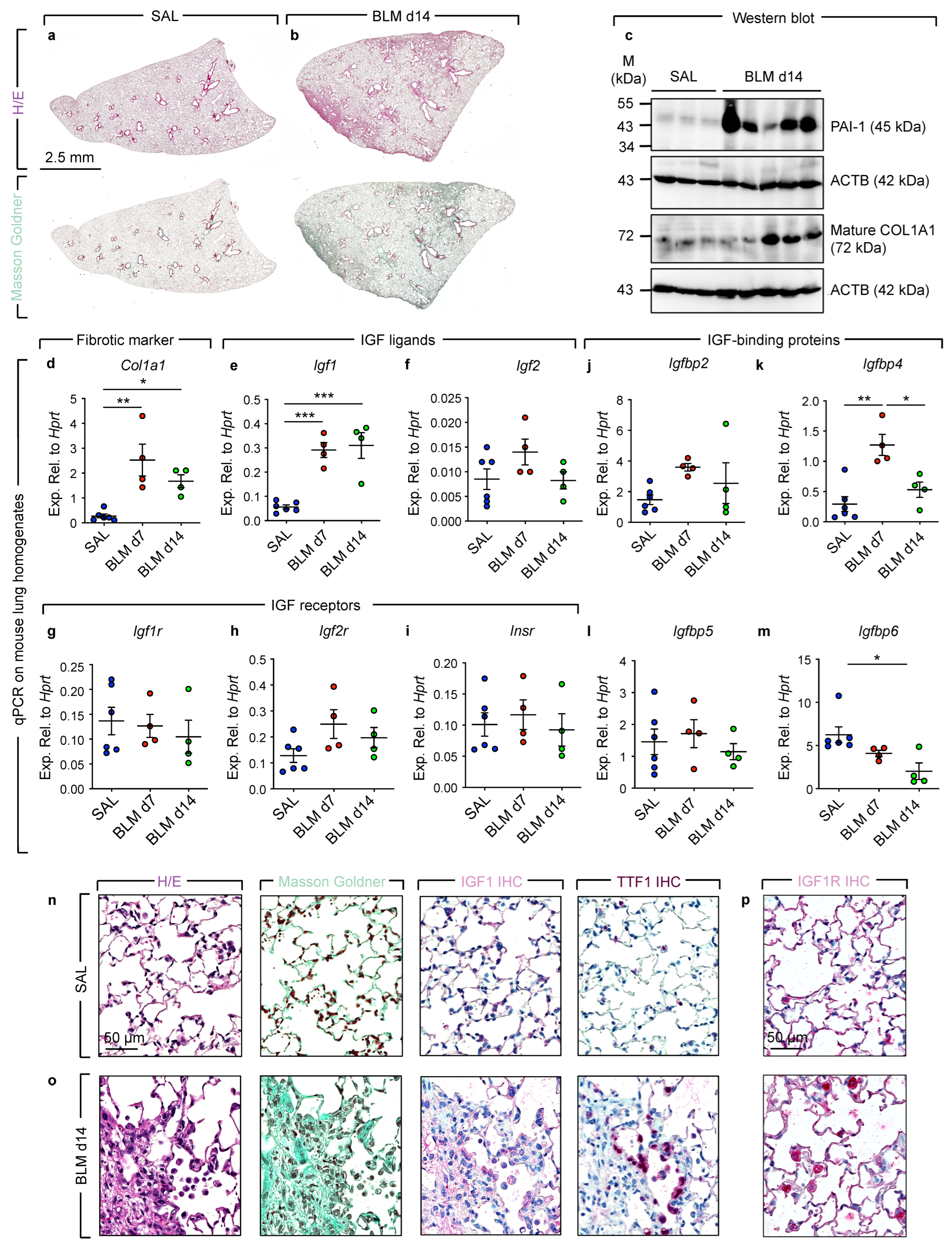
3.2. Increased Igf1 Expression during Fibrosis Development in Bleomycin-Induced Lung Injury
3.3. IGF1 Expression Is Elevated in IPF Lungs
3.4. Treatment of Human Lung Fibroblasts with Recombinant IGF1 Leads to Loss of Lipid Droplets
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, J.P.; Baker, J.; Perkins, A.S.; Robertson, E.J.; Efstratiadis, A. Mice Carrying Null Mutations of the Genes Encoding Insulin-like Growth Factor I (Igf-1) and Type 1 IGF Receptor (Igf1r). Cell 1993, 75, 59–72. [Google Scholar] [CrossRef]
- Keselman, A.C.; Martin, A.; Scaglia, P.A.; Sanguineti, N.M.; Armando, R.; Gutiérrez, M.; Braslavsky, D.; Ballerini, M.G.; Ropelato, M.G.; Ramirez, L.; et al. A Homozygous Mutation in the Highly Conserved Tyr60 of the Mature IGF1 Peptide Broadens the Spectrum of IGF1 Deficiency. Eur. J. Endocrinol. 2019, 181, K43–K53. [Google Scholar] [CrossRef]
- Giabicani, E.; Willems, M.; Steunou, V.; Chantot-Bastaraud, S.; Thibaud, N.; Abi Habib, W.; Azzi, S.; Lam, B.; Bérard, L.; Bony-Trifunovic, H.; et al. Increasing Knowledge in IGF1R Defects: Lessons from 35 New Patients. J. Med. Genet. 2020, 57, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Walenkamp, M.J.E.; Losekoot, M.; Wit, J.M. Molecular IGF-1 and IGF-1 Receptor Defects: From Genetics to Clinical Management. Endocr. Dev. 2013, 24, 128–137. [Google Scholar] [CrossRef]
- Yen, Y.-C.; Hsiao, J.-R.; Jiang, S.S.; Chang, J.S.; Wang, S.-H.; Shen, Y.-Y.; Chen, C.-H.; Chang, I.-S.; Chang, J.-Y.; Chen, Y.-W. Insulin-like Growth Factor-Independent Insulin-like Growth Factor Binding Protein 3 Promotes Cell Migration and Lymph Node Metastasis of Oral Squamous Cell Carcinoma Cells by Requirement of Integrin Β1. Oncotarget 2015, 6, 41837–41855. [Google Scholar] [CrossRef] [Green Version]
- Leroith, D.; Scheinman, E.J.; Bitton-Worms, K. The Role of Insulin and Insulin-like Growth Factors in the Increased Risk of Cancer in Diabetes. Rambam Maimonides Med. J. 2011, 2, e0043. [Google Scholar] [CrossRef]
- Pais, R.S.; Moreno-Barriuso, N.; Hernández-Porras, I.; López, I.P.; De Las Rivas, J.; Pichel, J.G. Transcriptome Analysis in Prenatal IGF1-Deficient Mice Identifies Molecular Pathways and Target Genes Involved in Distal Lung Differentiation. PLoS ONE 2013, 8, e83028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epaud, R.; Aubey, F.; Xu, J.; Chaker, Z.; Clemessy, M.; Dautin, A.; Ahamed, K.; Bonora, M.; Hoyeau, N.; Fléjou, J.-F.; et al. Knockout of Insulin-Like Growth Factor-1 Receptor Impairs Distal Lung Morphogenesis. PLoS ONE 2012, 7, e48071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, D.; Venihaki, M.; Guo, W.H.; Lopez, M.F. Igf2 Deficiency Results in Delayed Lung Development at the End of Gestation. Endocrinology 2006, 147, 5584–5591. [Google Scholar] [CrossRef]
- Ahamed, K.; Epaud, R.; Holzenberger, M.; Bonora, M.; Flejou, J.-F.; Puard, J.; Clement, A.; Henrion-Caude, A. Deficiency in Type 1 Insulin-like Growth Factor Receptor in Mice Protects against Oxygen-Induced Lung Injury. Respir. Res. 2005, 6, 31. [Google Scholar] [CrossRef] [Green Version]
- López, I.P.; Piñeiro-Hermida, S.; Pais, R.S.; Torrens, R.; Hoeflich, A.; Pichel, J.G. Involvement of Igf1r in Bronchiolar Epithelial Regeneration: Role during Repair Kinetics after Selective Club Cell Ablation. PLoS ONE 2016, 11, e0166388. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Ramchandran, R.; Chen, J.; Yang, Q.; Raj, J.U. Smooth Muscle Insulin-Like Growth Factor-1 Mediates Hypoxia-Induced Pulmonary Hypertension in Neonatal Mice. Am. J. Respir. Cell Mol. Biol. 2016, 55, 779–791. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.-E.; Lee, S.; Sunde, D.A.; Huizar, I.; Haugk, K.L.; Thannickal, V.J.; Vittal, R.; Plymate, S.R.; Schnapp, L.M. Insulin-like Growth Factor-I Receptor Blockade Improves Outcome in Mouse Model of Lung Injury. Am. J. Respir. Crit. Care Med. 2009, 179, 212–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehan, V.K.; Torday, J.S. The Lung Alveolar Lipofibroblast: An Evolutionary Strategy Against Neonatal Hyperoxic Lung Injury. Antioxid. Redox Signal. 2014, 21, 1893–1904. [Google Scholar] [CrossRef] [Green Version]
- Barkauskas, C.E.; Cronce, M.J.; Rackley, C.R.; Bowie, E.J.; Keene, D.R.; Stripp, B.R.; Randell, S.H.; Noble, P.W.; Hogan, B.L.M. Type 2 Alveolar Cells Are Stem Cells in Adult Lung. J. Clin. Investig. 2013, 123, 3025–3036. [Google Scholar] [CrossRef] [PubMed]
- Taghizadeh, S.; Heiner, M.; Vazquez-Armendariz, A.I.; Wilhelm, J.; Herold, S.; Chen, C.; Zhang, J.S.; Bellusci, S. Characterization in Mice of the Resident Mesenchymal Niche Maintaining AT2 Stem Cell Proliferation in Homeostasis and Disease. Stem Cells 2021, 39, 1382–1394. [Google Scholar] [CrossRef] [PubMed]
- El Agha, E.; Moiseenko, A.; Kheirollahi, V.; De Langhe, S.; Crnkovic, S.; Kwapiszewska, G.; Szibor, M.; Kosanovic, D.; Schwind, F.; Schermuly, R.T.; et al. Two-Way Conversion between Lipogenic and Myogenic Fibroblastic Phenotypes Marks the Progression and Resolution of Lung Fibrosis. Cell Stem Cell 2017, 20, 261–273.e3. [Google Scholar] [CrossRef] [Green Version]
- Kheirollahi, V.; Wasnick, R.M.; Biasin, V.; Vazquez-Armendariz, A.I.; Chu, X.; Moiseenko, A.; Weiss, A.; Wilhelm, J.; Zhang, J.-S.; Kwapiszewska, G.; et al. Metformin Induces Lipogenic Differentiation in Myofibroblasts to Reverse Lung Fibrosis. Nat. Commun. 2019, 10, 2987. [Google Scholar] [CrossRef]
- El Agha, E.; Kramann, R.; Schneider, R.K.; Li, X.; Seeger, W.; Humphreys, B.D.; Bellusci, S. Mesenchymal Stem Cells in Fibrotic Disease. Cell Stem Cell 2017, 21, 166–177. [Google Scholar] [CrossRef]
- Rosen, E.D. The Transcriptional Basis of Adipocyte Development. Prostaglandins Leukot. Essent. Fat. Acids 2005, 73, 31–34. [Google Scholar] [CrossRef]
- Al Alam, D.; El Agha, E.; Sakurai, R.; Kheirollahi, V.; Moiseenko, A.; Danopoulos, S.; Shrestha, A.; Schmoldt, C.; Quantius, J.; Herold, S.; et al. Evidence for the Involvement of Fibroblast Growth Factor 10 in Lipofibroblast Formation during Embryonic Lung Development. Development 2015, 142, 4139–4150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Agha, E.; Schwind, F.; Ruppert, C.; Günther, A.; Bellusci, S.; Schermuly, R.T.; Kosanovic, D. Is the Fibroblast Growth Factor Signaling Pathway a Victim of Receptor Tyrosine Kinase Inhibition in Pulmonary Parenchymal and Vascular Remodeling? Am. J. Physiol. Lung Cell. Mol. Physiol. 2018, 315, L248–L252. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, B.; Korfei, M.; Henneke, I.; Sibinska, Z.; Tian, X.; Hezel, S.; Dilai, S.; Wasnick, R.; Schneider, B.; Wilhelm, J.; et al. Increased FGF1-FGFRc Expression in Idiopathic Pulmonary Fibrosis. Respir. Res. 2015, 16, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Agha, E.; Bellusci, S. Walking along the Fibroblast Growth Factor 10 Route: A Key Pathway to Understand the Control and Regulation of Epithelial and Mesenchymal Cell-Lineage Formation during Lung Development and Repair after Injury. Scientifica 2014, 2014, 538379. [Google Scholar] [CrossRef]
- Volckaert, T.; De Langhe, S.P. Wnt and FGF Mediated Epithelial-Mesenchymal Crosstalk during Lung Development. Dev. Dyn. 2015, 244, 342–366. [Google Scholar] [CrossRef] [Green Version]
- Kina, Y.P.; Khadim, A.; Seeger, W.; El Agha, E. The Lung Vasculature: A Driver or Passenger in Lung Branching Morphogenesis? Front. Cell Dev. Biol. 2021, 8, 623868. [Google Scholar] [CrossRef]
- Chen, H.; Qu, J.; Huang, X.; Kurundkar, A.; Zhu, L.; Yang, N.; Venado, A.; Ding, Q.; Liu, G.; Antony, V.B.; et al. Mechanosensing by the A6-Integrin Confers an Invasive Fibroblast Phenotype and Mediates Lung Fibrosis. Nat. Commun. 2016, 7, 12564. [Google Scholar] [CrossRef] [Green Version]
- Hung, C.F.; Rohani, M.G.; Lee, S.; Chen, P.; Schnapp, L.M. Role of IGF-1 Pathway in Lung Fibroblast Activation. Respir. Res. 2013, 14, 102. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Huang, X.; Hecker, L.; Kurundkar, D.; Kurundkar, A.; Liu, H.; Jin, T.-H.; Desai, L.; Bernard, K.; Thannickal, V.J. Inhibition of Mechanosensitive Signaling in Myofibroblasts Ameliorates Experimental Pulmonary Fibrosis. J. Clin. Investig. 2013, 123, 1096–1108. [Google Scholar] [CrossRef]
- Hinz, B. Tissue Stiffness, Latent TGF-Β1 Activation, and Mechanical Signal Transduction: Implications for the Pathogenesis and Treatment of Fibrosis. Curr. Rheumatol. Rep. 2009, 11, 120. [Google Scholar] [CrossRef]
- Srinivasan, S.; Strange, J.; Awonusonu, F.; Bruce, M.C. Insulin-like Growth Factor I Receptor Is Downregulated after Alveolarization in an Apoptotic Fibroblast Subset. Am. J. Physiol. Lung Cell. Mol. Physiol. 2002, 282, 457–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.; Kitzmiller, J.A.; Sridharan, A.; Perl, A.K.; Bridges, J.P.; Misra, R.S.; Pryhuber, G.S.; Mariani, T.J.; Bhattacharya, S.; Guo, M.; et al. Lung Gene Expression Analysis (LGEA): An Integrative Web Portal for Comprehensive Gene Expression Data Analysis in Lung Development. Thorax 2017, 72, 481–484. [Google Scholar] [CrossRef] [Green Version]
- Moiseenko, A.; Kheirollahi, V.; Chao, C.-M.; Ahmadvand, N.; Quantius, J.; Wilhelm, J.; Herold, S.; Ahlbrecht, K.; Morty, R.E.; Rizvanov, A.A.; et al. Origin and Characterization of Alpha Smooth Muscle Actin-Positive Cells during Murine Lung Development. Stem Cells 2017, 35, 1566–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oherle, K.; Acker, E.; Bonfield, M.; Wang, T.; Gray, J.; Lang, I.; Bridges, J.; Lewkowich, I.; Xu, Y.; Ahlfeld, S.; et al. Insulin-like Growth Factor 1 Supports a Pulmonary Niche That Promotes Type 3 Innate Lymphoid Cell Development in Newborn Lungs. Immunity 2020, 52, 275–294.e9. [Google Scholar] [CrossRef]
- Garrett, S.M.; Hsu, E.; Thomas, J.M.; Pilewski, J.M.; Feghali-Bostwick, C. Insulin-like Growth Factor (IGF)-II- Mediated Fibrosis in Pathogenic Lung Conditions. PLoS ONE 2019, 14, e0225422. [Google Scholar] [CrossRef]
- Allard, J.B.; Duan, C. IGF-Binding Proteins: Why Do They Exist and Why Are There So Many? Front. Endocrinol. 2018, 9, 117. [Google Scholar] [CrossRef] [Green Version]
- Mouhieddine, O.B.; Cazals, V.; Kuto, E.; Le Bouc, Y.; Clement, A. Glucocorticoid-Induced Growth Arrest of Lung Alveolar Epithelial Cells Is Associated with Increased Production of Insulin-like Growth Factor Binding Protein-2. Endocrinology 1996, 137, 287–295. [Google Scholar] [CrossRef]
- Kim, M.S.; Lee, D.-Y. Insulin-like Growth Factor (IGF)-I and IGF Binding Proteins Axis in Diabetes Mellitus. Ann. Pediatr. Endocrinol. Metab. 2015, 20, 69–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.D.; Yin, L.; Archer, S.; Lu, C.; Zhao, G.; Yao, Y.; Wu, L.; Hsin, M.; Waddell, T.K.; Keshavjee, S.; et al. Metabolic Heterogeneity of Idiopathic Pulmonary Fibrosis: A Metabolomic Study. BMJ Open Respir. Res. 2017, 4, e000183. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Wen, Z.; Wang, R.; Luo, W.; Du, Y.; Wang, W.; Chen, X. Identification of the Lipid Biomarkers from Plasma in Idiopathic Pulmonary Fibrosis by Lipidomics. BMC Pulm. Med. 2017, 17, 174. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, M.C.G.-S.; Carrillo, G.; Pérez-Padilla, R.; Fernández-Plata, M.R.; Buendía-Roldán, I.; Vargas, M.H.; Selman, M. Risk Factors for Idiopathic Pulmonary Fibrosis in a Mexican Population. A Case-Control Study. Respir. Med. 2010, 104, 305–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Ma, Y.; Tong, X.; Zhang, Y.; Fan, H. Diabetes Mellitus Contributes to Idiopathic Pulmonary Fibrosis: A Review From Clinical Appearance to Possible Pathogenesis. Front. Public Health 2020, 8, 196. [Google Scholar] [CrossRef] [PubMed]
- Rangarajan, S.; Bone, N.B.; Zmijewska, A.A.; Jiang, S.; Park, D.W.; Bernard, K.; Locy, M.L.; Ravi, S.; Deshane, J.; Mannon, R.B.; et al. Metformin Reverses Established Lung Fibrosis in a Bleomycin Model. Nat. Med. 2018, 24, 1121–1127. [Google Scholar] [CrossRef]
- Choi, S.M.; Jang, A.-H.; Kim, H.; Lee, K.H.; Kim, Y.W. Metformin Reduces Bleomycin-Induced Pulmonary Fibrosis in Mice. J. Korean Med. Sci 2016, 31, 1419–1425. [Google Scholar] [CrossRef]
- Sato, N.; Takasaka, N.; Yoshida, M.; Tsubouchi, K.; Minagawa, S.; Araya, J.; Saito, N.; Fujita, Y.; Kurita, Y.; Kobayashi, K.; et al. Metformin Attenuates Lung Fibrosis Development via NOX4 Suppression. Respir. Res. 2016, 17, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilewski, J.M.; Liu, L.; Henry, A.C.; Knauer, A.V.; Feghali-Bostwick, C.A. Insulin-like Growth Factor Binding Proteins 3 and 5 Are Overexpressed in Idiopathic Pulmonary Fibrosis and Contribute to Extracellular Matrix Deposition. Am. J. Pathol. 2005, 166, 399–407. [Google Scholar] [CrossRef] [Green Version]
- Guiot, J.; Bondue, B.; Henket, M.; Corhay, J.L.; Louis, R. Raised Serum Levels of IGFBP-1 and IGFBP-2 in Idiopathic Pulmonary Fibrosis. BMC Pulm. Med. 2016, 16, 86. [Google Scholar] [CrossRef] [Green Version]
- Guiot, J.; Henket, M.; Corhay, J.-L.; Louis, R. Serum IGFBP2 as a Marker of Idiopathic Pulmonary Fibrosis. Eur. Respir. J. 2015, 46, PA3840. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′-3′) |
|---|---|
| hACTA2 Fwd | CTGTTCCAGCCATCCTTCAT |
| hACTA2 Rev | TCATGATGCTGTTGTAGGTGGT |
| hCOL1A1 Fwd | ATGTTCAGCTTTGTGGACCTC |
| hCOL1A1 Rev | CTGTACGCAGGTGATTGGTG |
| hIGF1 Fwd | TGTGGAGACAGGGGCTTTTA |
| hIGF1 Rev | ATCCACGATGCCTGTCTGA |
| hIGF1R Fwd | GAGAATTTCCTTCACAATTCCATC |
| hIGF1R Rev | CACTTGCATGACGTCTCTCC |
| hIGF2 Fwd | CAAACCGAGCTGGGCG |
| hIGF2 Rev | CACAGAGAAGCGGAGGGA |
| hIGF2R Fwd | TCTCCAGTGGACTGCCAAGT |
| hIGF2R Rev | GTGCTTAGGCCAGTCAGGTC |
| hIGFBP1 Fwd | AATGGATTTTATCACAGCAGACAG |
| hIGFBP1 Rev | GGTAGACGCACCAGCAGAGT |
| hIGFBP2 Fwd | AAGGGTGGCAAGCATCAC |
| hIGFBP2 Rev | CTGGTCCAGTTCCTGTTGG |
| hIGFBP3 Fwd | AACGCTAGTGCCGTCAGC |
| hIGFBP3 Rev | CGGTCTTCCTCCGACTCAC |
| hIGFBP4 Fwd | CCTCTACATCATCCCCATCC |
| hIGFBP4 Rev | GGTCCACACACCAGCACTT |
| hIGFBP5 Fwd | AGAGCTACCGCGAGCAAGT |
| hIGFBP5 Rev | GTAGGTCTCCTCGGCCATCT |
| hIGFBP6 Fwd | TGACCATCGAGGCTTCTACC |
| hIGFBP6 Rev | CATCCGATCCACACACCA |
| hINSRA Fwd | TTTTCGTCCCCAGGCCATC |
| hINSRB Fwd | CCCCAGAAAAACCTCTTCAGG |
| hINSR Rev | GTCACATTCCCAACATCGCC |
| hPBGD Fwd | TGTCTGGTAACGGCAATGCG |
| hPBGD Rev | CCCACGCGAATCACTCTCAT |
| hPLIN2 Fwd | TCAGCTCCATTCTACTGTTCACC |
| hPLIN2 Rev | CCTGAATTTTCTGATTGGCAC |
| hPPARG Fwd | TTGCTGTCATTATTCTCAGTGGA |
| hPPARG Rev | GAGGACTCAGGGTGGTTCAG |
| mCol1a1 Fwd | CCAAGAAGACATCCCTGAAGTCA |
| mCol1a1 Rev | TGCACGTCATCGCACACA |
| mHprt Fwd | CCTAAGATGAGCGCAAGTTGAA |
| mHprt Rev | CCACAGGACTAGAACACCTGCTAA |
| mIgf1 Fwd | AGCAGCCTTCCAACTCAATTAT |
| mIgf1 Rev | GAAGACGACATGATGTGTATCTTTATC |
| mIgf1r Fwd | AGAATTTCCTTCACAATTCCATC |
| mIgf1r Rev | CACTTGCATGACGTCTCTCC |
| mIgf2 Fwd | CGCTTCAGTTTGTCTGTTCG |
| mIgf2 Rev | GCAGCACTCTTCCACGATG |
| mIgf2r Fwd | CCTTCTCTAGTGGATTGTCAAGTG |
| migf2r Rev | AGGGCGCTCAAGTCATACTC |
| mIgfbp1 Fwd | TGGTCAGGGAGCCTGTGTA |
| mIgfbp1 Rev | ACAGCAGCCTTTGCCTCTT |
| mIgfbp2 Fwd | GCGGGTACCTGTGAAAAGAG |
| mIgfbp2 Rev | CCTCAGAGTGGTCGTCATCA |
| mIgfbp3 Fwd | GACGACGTACATTGCCTCAG |
| mIgfbp3 Rev | GACGACGTACATTGCCTCAG |
| mIgfbp4 Fwd | GACACCTCGGGAGGAACC |
| mIgfbp4 Rev | AAGAGGTCTTCGTGGGTACG |
| mIgfbp5 Fwd | GGCGAGCAAACCAAGATAGA |
| mIgfbp5 Rev | AGGTCTCTTCAGCCATCTCG |
| mIgfbp6 Fwd | GGGCTCTATGTGCCAAACTG |
| mIgfbp6 Rev | CCTGCGAGGAACGACACT |
| mInsr Fwd | TCTTTCTTCAGGAAGCTACATCTG |
| mInsr Rev | TGTCCAAGGCATAAAAAGAATAGTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kheirollahi, V.; Khadim, A.; Kiliaris, G.; Korfei, M.; Barroso, M.M.; Alexopoulos, I.; Vazquez-Armendariz, A.I.; Wygrecka, M.; Ruppert, C.; Guenther, A.; et al. Transcriptional Profiling of Insulin-like Growth Factor Signaling Components in Embryonic Lung Development and Idiopathic Pulmonary Fibrosis. Cells 2022, 11, 1973. https://doi.org/10.3390/cells11121973
Kheirollahi V, Khadim A, Kiliaris G, Korfei M, Barroso MM, Alexopoulos I, Vazquez-Armendariz AI, Wygrecka M, Ruppert C, Guenther A, et al. Transcriptional Profiling of Insulin-like Growth Factor Signaling Components in Embryonic Lung Development and Idiopathic Pulmonary Fibrosis. Cells. 2022; 11(12):1973. https://doi.org/10.3390/cells11121973
Chicago/Turabian StyleKheirollahi, Vahid, Ali Khadim, Georgios Kiliaris, Martina Korfei, Margarida Maria Barroso, Ioannis Alexopoulos, Ana Ivonne Vazquez-Armendariz, Malgorzata Wygrecka, Clemens Ruppert, Andreas Guenther, and et al. 2022. "Transcriptional Profiling of Insulin-like Growth Factor Signaling Components in Embryonic Lung Development and Idiopathic Pulmonary Fibrosis" Cells 11, no. 12: 1973. https://doi.org/10.3390/cells11121973