
Changes in the Oxidation-Reduction State of Human Dermal Fibroblasts as an Effect of Lomefloxacin Phototoxic Action

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Culture and the Exposure to Lomefloxacin and UVA Radiation
2.3. Cell Viability Assay
2.4. Cell Proliferation Assay
2.5. Cell Cycle Analysis
2.6. H2DCFDA Assay
2.7. Confocal Microscopy Imaging
2.8. Western Blotting Analysis
2.9. The Estimation of Cellular Reduced Glutathione Level
2.10. Statistical Analysis
3. Results
3.1. UVA Radiation Induces Cytotoxicity of Lomefloxacin
3.2. Lomefloxacin Suppresses the Proliferation of Fibroblasts
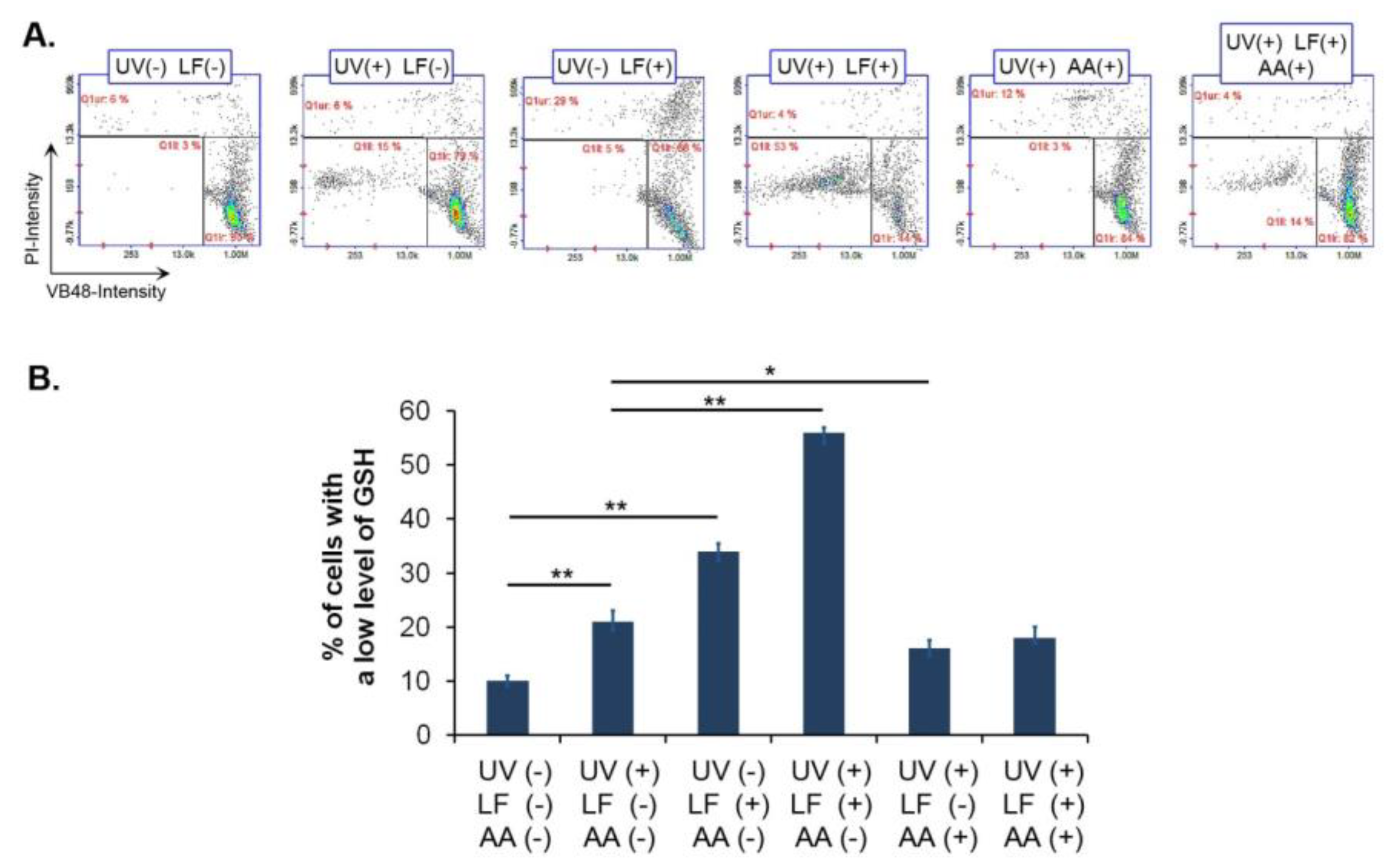
3.3. UVA Radiation Enhances Oxidative Stress Induced by Lomefloxacin
3.4. The Impact of Lomefloxacin and UVA Radiation on Antioxidant Enzymes Expression
3.5. Lomefloxacin and UVA Radiation Decrease Vitality of Fibroblasts
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Al Aboud, D.M.; Nessel, T.A.; Hafsi, W. Cutaneous Adverse Drug Reaction. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK533000/ (accessed on 10 February 2022).
- Dawe, R.S.; Ibbotson, S.H. Drug-induced photosensitivity. Dermatol. Clin. 2014, 32, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Drucker, A.M.; Rosen, C.F. Drug-induced photosensitivity: Culprit drugs, management and prevention. Drug. Saf. 2011, 34, 821–837. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.E. Drug-induced cutaneous photosensitivity: Incidence, mechanism, prevention and management. Drug Saf. 2002, 25, 345–372. [Google Scholar] [CrossRef]
- Kowalska, J.; Rok, J.; Rzepka, Z.; Wrześniok, D. Drug-induced photosensitivity-from light and chemistry to biological reactions and clinical symptoms. Pharmaceuticals 2021, 14, 723. [Google Scholar] [CrossRef]
- Hofmann, G.A.; Weber, B. Drug-induced photosensitivity: Culprit drugs, potential mechanisms and clinical consequences. J. Dtsch. Dermatol. Ges. 2021, 19, 19–29. [Google Scholar] [CrossRef]
- Monteiro, A.F.; Rato, M.; Martins, C. Drug-induced photosensitivity: Photoallergic and phototoxic reactions. Clin. Dermatol. 2016, 34, 571–581. [Google Scholar] [CrossRef]
- Dubakiene, R.; Kupriene, M. Scientific problems of photosensitivity. Medicina 2006, 42, 619–624. [Google Scholar] [PubMed]
- Sprandel, K.A.; Rodvold, K.A. Safety and tolerability of fluoroquinolones. Clin. Cornerstone 2003, 3, S29–S36. [Google Scholar] [CrossRef]
- Man, I.; Murphy, J.; Ferguson, J. Fluoroquinolone phototoxicity: A comparison of moxifloxacin and lomefloxacin in normal volunteers. J. Antimicrob. Chemother. 1999, 43, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Eljaaly, K.; Alkhalaf, A.; Alhifany, A.A.; Alshibani, M. Photosensitivity induced by lomefloxacin versus other fluoroquinolones: A meta-analysis. J. Infect. Chemother. 2020, 26, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Lipsky, B.A.; Baker, C.A. Fluoroquinolone toxicity profiles: A review focusing on newer agents. Clin. Infect. Dis. 1999, 28, 352–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Vries, H.; van Henegouwen, G.M. Photochemical decomposition of lomefloxacin in vitro and in vivo. J. Photochem. Photobiol. B. 2000, 58, 6–12. [Google Scholar] [CrossRef]
- Zelmat, Y.; Rousseau, V.; Chebane, L.; Montastruc, J.L.; Bagheri, H.; Sommet, A. Fluoroquinolone-induced photosensitivity: A chemical fragment-based approach by a case/non-case study in VigiBase®. Drug Saf. 2020, 43, 561–566. [Google Scholar] [CrossRef]
- Ray, R.S.; Agrawal, N.; Misra, R.B.; Farooq, M.; Hans, R.K. Radiation-induced in vitro phototoxic potential of some fluoroquinolones. Drug Chem. Toxicol. 2006, 29, 25–38. [Google Scholar] [CrossRef]
- Beberok, A.; Buszman, E.; Zdybel, M.; Pilawa, B.; Wrześniok, D. EPR examination of free radical properties of DOPA–melanin complexes with ciprofloxacin, lomefloxacin, norfloxacin and sparfloxacin. Chem. Phys. Lett. 2010, 497, 115–122. [Google Scholar] [CrossRef]
- Beberok, A.; Otręba, M.; Wrześniok, D.; Buszman, E. Cytotoxic effect of lomefloxacin in culture of human epidermal melanocytes. Pharmacol. Rep. 2013, 65, 689–699. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, J.; Banach, K.; Rok, J.; Beberok, A.; Rzepka, Z.; Wrześniok, D. Molecular and biochemical basis of fluoroquinolones-induced phototoxicity-the study of antioxidant system in human melanocytes exposed to UV-A radiation. Int. J. Mol. Sci. 2020, 21, 9714. [Google Scholar] [CrossRef]
- Kowalska, J.; Banach, K.; Beberok, A.; Rok, J.; Rzepka, Z.; Wrześniok, D. The biochemical and molecular analysis of changes in melanogenesis induced by UVA-activated fluoroquinolones-in vitro study on human normal melanocytes. Cells 2021, 10, 2900. [Google Scholar] [CrossRef]
- Slominski, A.T.; Zmijewski, M.A.; Plonka, P.M.; Szaflarski, J.P.; Paus, R. How UV light touches the brain and endocrine system through skin, and why. Endocrinology 2018, 159, 1992–2007. [Google Scholar] [CrossRef] [Green Version]
- Chambers, E.S.; Vukmanovic-Stejic, M. Skin barrier immunity and ageing. Immunology 2020, 160, 116–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rippa, A.L.; Kalabusheva, E.P.; Vorotelyak, E.A. Regeneration of dermis: Scarring and cells involved. Cells 2019, 8, 607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thulabandu, V.; Chen, D.; Atit, R.P. Dermal fibroblast in cutaneous development and healing. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, e307. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Viennet, C.; Robin, S.; Berthon, J.Y.; He, L.; Humbert, P. Precise role of dermal fibroblasts on melanocyte pigmentation. J. Dermatol. Sci. 2017, 88, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Cell Vitality Assay. Detection of Cell Vitality and Thiol Levels Using VitaBright-48™. Available online: https://chemometec.com/resources/cell-assays/cell-vitality-assay/ (accessed on 24 May 2022).
- Bickers, D.R.; Athar, M. Oxidative stress in the pathogenesis of skin disease. J. Investig. Dermatol. 2006, 126, 2565–2575. [Google Scholar] [CrossRef] [Green Version]
- Trouba, K.J.; Hamadeh, H.K.; Amin, R.P.; Germolec, D.R. Oxidative stress and its role in skin disease. Antioxid. Redox Signal. 2002, 4, 665–673. [Google Scholar] [CrossRef]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [Green Version]
- Solano, F. Melanins: Skin pigments and much more—types, structural models, biological functions, and formation routes. New J. Sci. 2014, 2014, 498276. [Google Scholar] [CrossRef] [Green Version]
- Larsson, B.S. Interaction between chemicals and melanin. Pigment Cell Res. 1993, 6, 127–133. [Google Scholar] [CrossRef]
- Meyskens, F.L., Jr.; Farmer, P.; Fruehauf, J.P. Redox regulation in human melanocytes and melanoma. Pigment Cell Res. 2001, 14, 148–154. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Tonissen, K.F.; Di Trapani, G. Modulating skin colour: Role of the thioredoxin and glutathione systems in regulating melanogenesis. Biosci. Rep. 2021, 41, BSR20210427. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef] [PubMed]
- Slominski, R.M.; Sarna, T.; Płonka, P.M.; Raman, C.; Brożyna, A.A.; Slominski, A.T. Melanoma, melanin, and melanogenesis: The yin and yang relationship. Front. Oncol. 2022, 12, 842496. [Google Scholar] [CrossRef] [PubMed]
- Yohn, J.J.; Norris, D.A.; Yrastorza, D.G.; Buno, I.J.; Leff, J.A.; Hake, S.S.; Repine, J.E. Disparate antioxidant enzyme activities in cultured human cutaneous fibroblasts, keratinocytes, and melanocytes. J. Investig. Dermatol. 1991, 97, 405–409. [Google Scholar] [CrossRef] [Green Version]
- Beberok, A.; Wrześniok, D.; Rzepka, Z.; Rok, J.; Delijewski, M.; Otręba, M.; Respondek, M.; Buszman, E. Effect of fluoroquinolones on melanogenesis in normal human melanocytes HEMn-DP: A comparative in vitro study. Cutan. Ocul. Toxicol. 2017, 36, 169–175. [Google Scholar] [CrossRef]
- ICH Safety Guidelines S10 Photosafety Evaluation of Pharmaceuticals. Available online: https://database.ich.org/sites/default/files/S10_Guideline.pdf (accessed on 24 May 2022).
- Hamanaka, H.; Mizutani, H.; Asahig, K.; Shimizu, M. Melanocyte melanin augments sparfloxacin-induced phototoxicity. J. Dermatol. Sci. 1999, 21, 27–33. [Google Scholar] [CrossRef]
- Zhao, B.; Chignell, C.; Rammal, M.; Smith, F.; Hamilton, M.; Andley, U.; Roberts, J. Detection and prevention of ocular phototoxicity of ciprofloxacin and other fluoroquinolone antibiotics. Photochem. Photobiol. 2010, 86, 798–805. [Google Scholar] [CrossRef] [Green Version]
- Aloisi, G.; Barbafina, A.; Canton, M.; Dall’Acqua, F.; Elisei, F.; Facciolo, L.; Latterini, L.; Viola, G. Photophysical and photobiological behavior of antimalarial drugs in aqueous solutions. Photochem. Photobiol. 2004, 79, 248–258. [Google Scholar] [CrossRef]
- Williams, R.J., 3rd; Attia, E.; Wickiewicz, T.L.; Hannafin, J.A. The effect of ciprofloxacin on tendon, paratenon, and capsular fibroblast metabolism. Am. J. Sports Med. 2000, 28, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Herold, C.; Ocker, M.; Ganslmayer, M.; Gerauer, H.; Hahn, E.G.; Schuppan, D. Ciprofloxacin induces apoptosis and inhibits proliferation of human colorectal carcinoma cells. Br. J. Cancer 2002, 86, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Yadav, V.; Sultana, S.; Yadav, J.; Saini, N. Gatifloxacin induces S and G2-phase cell cycle arrest in pancreatic cancer cells via p21/p27/p53. PLoS ONE 2012, 7, e47796. [Google Scholar] [CrossRef] [PubMed]
- Beberok, A.; Rok, J.; Rzepka, Z.; Marciniec, K.; Boryczka, S.; Wrześniok, D. The role of MITF and Mcl-1 proteins in the antiproliferative and proapoptotic effect of ciprofloxacin in amelanotic melanoma cells: In silico and in vitro study. Toxicol. Vitr. 2020, 66, 104884. [Google Scholar] [CrossRef] [PubMed]
- Beberok, A.; Rzepka, Z.; Respondek, M.; Rok, J.; Stradowski, M.; Wrześniok, D. Moxifloxacin as an inducer of apoptosis in melanoma cells: A study at the cellular and molecular level. Toxicol. Vitr. 2019, 55, 75–92. [Google Scholar] [CrossRef] [PubMed]
- Marrot, L.; Belaïdi, J.P.; Jones, C.; Perez, P.; Riou, L.; Sarasin, A.; Meunier, J.R. Molecular responses to photogenotoxic stress induced by the antibiotic lomefloxacin in human skin cells: From DNA damage to apoptosis. J. Investig. Dermatol. 2003, 121, 596–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuht, H.; Lode, H.; Koeppe, P.; Rost, K.L.; Schaberg, T. Interaction study of lomefloxacin and ciprofloxacin with omeprazole and comparative pharmacokinetics. Antimicrob. Agents Chemother. 1995, 39, 1045–1049. [Google Scholar] [CrossRef] [Green Version]
- Matés, J.M.; Pérez-Gómez, C.; Núñez de Castro, I. Antioxidant enzymes and human diseases. Clin. Biochem. 1999, 32, 595–603. [Google Scholar] [CrossRef]
- Zabłocka, A.; Janusz, M. Dwa oblicza wolnych rodników tlenowych [The two faces of reactive oxygen species]. Postepy Hig. Med. Dosw. 2008, 62, 118–124. [Google Scholar]
- Kim, G.H.; Kim, J.E.; Rhie, S.J.; Yoon, S. The role of oxidative stress in neurodegenerative diseases. Exp. Neurobiol. 2015, 24, 325–340. [Google Scholar] [CrossRef]
- Espinosa-Diez, C.; Miguel, V.; Mennerich, D.; Kietzmann, T.; Sánchez-Pérez, P.; Cadenas, S.; Lamas, S. Antioxidant responses and cellular adjustments to oxidative stress. Redox. Biol. 2015, 6, 183–197. [Google Scholar] [CrossRef] [Green Version]
- Adeoye, O.; Olawumi, J.; Opeyemi, A.; Christiania, O. Review on the role of glutathione on oxidative stress and infertility. JBRA Assist. Reprod. 2018, 22, 61–66. [Google Scholar] [CrossRef]
- Golden, T.R.; Hinerfeld, D.A.; Melov, S. Oxidative stress and aging: Beyond correlation. Aging Cell 2002, 1, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Peng, S.; Sheng, Z.; Wang, Y. Ofloxacin induces oxidative damage to joint chondrocytes of juvenile rabbits: Excessive production of reactive oxygen species, lipid peroxidation and DNA damage. Eur. J. Pharmacol. 2010, 626, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Beberok, A.; Wrześniok, D.; Otręba, M.; Buszman, E. Impact of sparfloxacin on melanogenesis and antioxidant defense system in normal human melanocytes HEMa-LP—An in vitro study. Pharmacol. Rep. 2015, 67, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Cuquerella, M.C.; Miranda, M.A.; Bosca, F. Generation of detectable singlet aryl cations by photodehalogenation of fluoroquinolones. J. Phys. Chem. B 2006, 110, 6441–6443. [Google Scholar] [CrossRef] [PubMed]
- Marklund, S.L. Extracellular superoxide dismutase in human tissues and human cell lines. J. Clin. Investig. 1984, 74, 1398–1403. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalska, J.; Banach, K.; Rzepka, Z.; Rok, J.; Karkoszka, M.; Wrześniok, D. Changes in the Oxidation-Reduction State of Human Dermal Fibroblasts as an Effect of Lomefloxacin Phototoxic Action. Cells 2022, 11, 1971. https://doi.org/10.3390/cells11121971
Kowalska J, Banach K, Rzepka Z, Rok J, Karkoszka M, Wrześniok D. Changes in the Oxidation-Reduction State of Human Dermal Fibroblasts as an Effect of Lomefloxacin Phototoxic Action. Cells. 2022; 11(12):1971. https://doi.org/10.3390/cells11121971
Chicago/Turabian StyleKowalska, Justyna, Klaudia Banach, Zuzanna Rzepka, Jakub Rok, Marta Karkoszka, and Dorota Wrześniok. 2022. "Changes in the Oxidation-Reduction State of Human Dermal Fibroblasts as an Effect of Lomefloxacin Phototoxic Action" Cells 11, no. 12: 1971. https://doi.org/10.3390/cells11121971