
Rapid Actions of the Nuclear Progesterone Receptor through cSrc in Cancer


{kind=link}
{kind=link}
Abstract
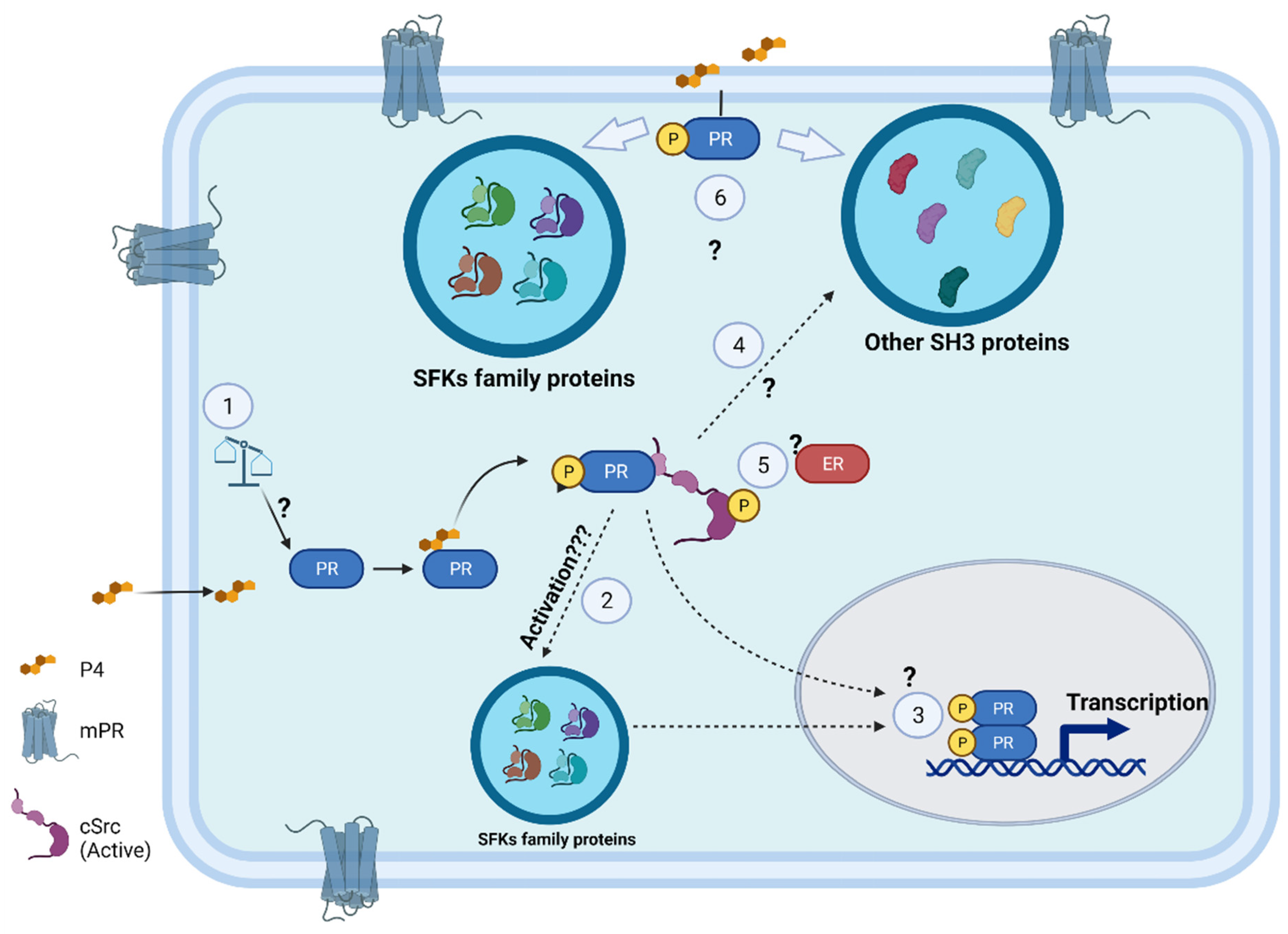
:1. Introduction
2. SH3 Domain–PXPP Motif Interaction: Structural Basis and Functions
3. Functions of the Polyproline Motif of PR in Breast Cancer
4. Role of PR and cSrc in Glioblastoma Progression
4.1. Contribution of cSrc to Malignancy of Glioblastoma
4.2. PR: An Underappreciated Villain in GB Progression
4.3. PR–cSrc Interaction in GBs: Experimental Evidence and Future Perspectives
5. Relevance of Possible cSrc–PR Interaction in Other Cancers
6. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tania Hernandez-Hernandez, O.; Camacho-Arroyo, I. Regulation of Gene Expression by Progesterone in Cancer Cells: Effects on Cyclin D1, EGFR and VEGF. Mini Rev. Med. Chem. 2013, 13, 635–642. [Google Scholar] [CrossRef]
- Valadez-Cosmes, P.; Vázquez-Martínez, E.R.; Cerbón, M.; Camacho-Arroyo, I. Membrane Progesterone Receptors in Reproduction and Cancer. Mol. Cell. Endocrinol. 2016, 434, 166–175. [Google Scholar] [CrossRef]
- Brisken, C.; Scabia, V. 90 years of progesterone: Progesterone Receptor Signaling in the Normal Breast and Its Implications for Cancer. J. Mol. Endocrinol. 2020, 65, T81–T94. [Google Scholar] [CrossRef]
- Singh, M.; Su, C.; Ng, S. Non-Genomic Mechanisms of Progesterone Action in the Brain. Front. Neurosci. 2013, 7, 159. [Google Scholar] [CrossRef] [Green Version]
- Garg, D.; Ng, S.S.M.; Baig, K.M.; Driggers, P.; Segars, J. Progesterone-Mediated Non-Classical Signaling. Trends Endocrinol. Metab. 2017, 28, 656–668. [Google Scholar] [CrossRef]
- Moussatche, P.; Lyons, T.J. Non-Genomic Progesterone Signalling and Its Non-Canonical Receptor. Biochem. Soc. Trans. 2012, 40, 200–204. [Google Scholar] [CrossRef] [Green Version]
- Peluso, J.J.; Pru, J.K. Non-Canonical Progesterone Signaling in Granulosa Cell Function. Reproduction 2014, 147, R169–R178. [Google Scholar] [CrossRef] [Green Version]
- Peluso, J.J.; Pru, J.K. Progesterone Receptor Membrane Component (PGRMC)1 and PGRMC2 and Their Roles in Ovarian and Endometrial Cancer. Cancers 2021, 13, 5953. [Google Scholar] [CrossRef] [PubMed]
- Asperger, H.; Stamm, N.; Gierke, B.; Pawlak, M.; Hofmann, U.; Zanger, U.M.; Marton, A.; Katona, R.L.; Buhala, A.; Vizler, C.; et al. Progesterone Receptor Membrane Component 1 Regulates Lipid Homeostasis and Drives Oncogenic Signaling Resulting in Breast Cancer Progression. Breast Cancer Res. 2020, 22, 75. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Arroyo, I.; Hansberg-Pastor, V.; Vázquez-Martínez, E.R.; Cerbón, M. Mechanism of Progesterone Action in the Brain. In Hormones, Brain and Behavior; Elsevier: Oxford, UK, 2017; pp. 181–214. [Google Scholar]
- Boonyaratanakornkit, V.; Edwards, D.P. Receptor Mechanisms Mediating Non-Genomic Actions of Sex Steroids. Semin. Reprod. Med. 2007, 25, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Yeatman, T.J. A Renaissance for SRC. Nat. Rev. Cancer 2004, 4, 470–480. [Google Scholar] [CrossRef]
- Migliaccio, A.; Piccolo, D.; Castoria, G.; Di Domenico, M.; Bilancio, A.; Lombardi, M.; Gong, W.; Beato, M.; Auricchio, F. Activation of the Src/P21ras/Erk Pathway by Progesterone Receptor via Cross-Talk with Estrogen Receptor. EMBO J. 1998, 17, 2008. [Google Scholar] [CrossRef] [Green Version]
- Ballaré, C.; Uhrig, M.; Bechtold, T.; Sancho, E.; Di Domenico, M.; Migliaccio, A.; Auricchio, F.; Beato, M. Two Domains of the Progesterone Receptor Interact with the Estrogen Receptor and Are Required for Progesterone Activation of the C-Src/Erk Pathway in Mammalian Cells. Mol. Cell. Biol. 2003, 23, 1994–2008. [Google Scholar] [CrossRef] [Green Version]
- Boonyaratanakornkit, V.; Scott, M.P.; Ribon, V.; Sherman, L.; Anderson, S.M.; Maller, J.L.; Miller, W.T.; Edwards, D.P. Progesterone Receptor Contains a Proline-Rich Motif That Directly Interacts with SH3 Domains and Activates c-Src Family Tyrosine Kinases. Mol. Cell 2001, 8, 269–280. [Google Scholar] [CrossRef]
- Boonyaratanakornkit, V.; McGowan, E.; Sherman, L.; Mancini, M.A.; Cheskis, B.J.; Edwards, D.P. The Role of Extranuclear Signaling Actions of Progesterone Receptor in Mediating Progesterone Regulation of Gene Expression and the Cell Cycle. Mol. Endocrinol. 2007, 21, 359–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, X.D.; Goglia, L.; Sanchez, A.M.; Flamini, M.; Giretti, M.S.; Tosi, V.; Genazzani, A.R.; Simoncini, T. Progesterone Receptor Enhances Breast Cancer Cell Motility and Invasion via Extranuclear Activation of Focal Adhesion Kinase. Endocr. Relat. Cancer 2010, 17, 431–443. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.C.; Lee, W. Sen Molecular Mechanisms Underlying Progesterone-Enhanced Breast Cancer Cell Migration. Sci. Rep. 2016, 6, 31509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, B.J.; Hamaguchi, M.; Hanafusa, H. A Novel Viral Oncogene with Structural Similarity to Phospholipase C. Nature 1988, 332, 272–275. [Google Scholar] [CrossRef]
- Stahl, M.L.; Ferenz, C.R.; Kelleher, K.L.; Kriz, R.W.; Knopf, J.L. Sequence Similarity of Phospholipase C with the Non-Catalytic Region of Src. Nature 1988, 332, 269–272. [Google Scholar] [CrossRef]
- Teyra, J.; Huang, H.; Jain, S.; Guan, X.; Dong, A.; Liu, Y.; Tempel, W.; Min, J.; Tong, Y.; Kim, P.M.; et al. Comprehensive Analysis of the Human SH3 Domain Family Reveals a Wide Variety of Non-Canonical Specificities. Structure 2017, 25, 1598–1610.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, J.M.; Riethmüller, G.; Johnson, J.P. Nck, a Melanoma CDNA Encoding a Cytoplasmic Protein Consisting of the Src Homology Units SH2 and SH3. Nucleic Acids Res. 1990, 18, 1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurochkina, N.; Guha, U. SH3 Domains: Modules of Protein-Protein Interactions. Biophys. Rev. 2013, 5, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Saksela, K.; Permi, P. SH3 Domain Ligand Binding: What’s the Consensus and Where’s the Specificity? FEBS Lett. 2012, 586, 2609–2614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, A.M.; Zondlo, N.J. A Propensity Scale for Type II Polyproline Helices (PPII): Aromatic Amino Acids in Proline-Rich Sequences Strongly Disfavor PPII Due to Proline-Aromatic Interactions. Biochemistry 2012, 51, 5041–5051. [Google Scholar] [CrossRef] [PubMed]
- Chandra, B.R.; Gowthaman, R.; Akhouri, R.R.; Gupta, D.; Sharma, A. Distribution of Proline-rich (PxxP) Motifs in Distinct Proteomes: Functional and Therapeutic Implications for Malaria and Tuberculosis. Protein Eng. Des. Sel. 2004, 17, 175–182. [Google Scholar] [CrossRef]
- Feng, S.; Chen, J.K.; Yu, H.; Simon, J.A.; Schreiber, S.L. Two Binding Orientations for Peptides to the Src SH3 Domain: Development of a General Model for SH3-Ligand Interactions. Science 1994, 266, 1241–1247. [Google Scholar] [CrossRef]
- Yu, H.; Chen, J.K.; Feng, S.; Dalgarno, D.C.; Brauer, A.W.; Schrelber, S.L. Structural Basis for the Binding of Proline-Rich Peptides to SH3 Domains. Cell 1994, 76, 933–945. [Google Scholar] [CrossRef]
- Weng, Z.; Rickles, R.J.; Feng, S.; Richard, S.; Shaw, A.S.; Schreiber, S.L.; Brugge, J.S. Structure-Function Analysis of SH3 Domains: SH3 Binding Specificity Altered by Single Amino Acid Substitutions. Mol. Cell. Biol. 1995, 15, 5627–5634. [Google Scholar] [CrossRef] [Green Version]
- Fajer, M.; Meng, Y.; Roux, B. The Activation of C-Src Tyrosine Kinase: Conformational Transition Pathway and Free Energy Landscape. J. Phys. Chem. B 2017, 121, 3352–3363. [Google Scholar] [CrossRef]
- Sieg, D.J.; Hauck, C.R.; Schlaepfer, D.D. Required Role of Focal Adhesion Kinase (FAK) for Integrin-Stimulated Cell Migration. J. Cell Sci. 1999, 112, 2677–2691. [Google Scholar] [CrossRef]
- Anand-Apte, B.; Zetter, B.R.; Viswanathan, A.; Qiu, R.G.; Chen, J.; Ruggieri, R.; Symons, M. Platelet-Derived Growth Factor and Fibronectin-Stimulated Migration Are Differentially Regulated by the Rac and Extracellular Signal-Regulated Kinase Pathways. J. Biol. Chem. 1997, 272, 30688–30692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weikum, E.R.; Liu, X.; Ortlund, E.A. The Nuclear Receptor Superfamily: A Structural Perspective. Protein Sci. 2018, 27, 1876–1892. [Google Scholar] [CrossRef] [PubMed]
- Kastner, P.; Krust, A.; Turcotte, B.; Stropp, U.; Tora, L.; Gronemeyer, H.; Chambon, P. Two Distinct Estrogen-Regulated Promoters Generate Transcripts Encoding the Two Functionally Different Human Progesterone Receptor Forms A and B. EMBO J. 1990, 9, 1603–1614. [Google Scholar] [CrossRef] [PubMed]
- Pedram, A.; Razandi, M.; Sainson, R.C.A.; Kim, J.K.; Hughes, C.C.; Levin, E.R. A Conserved Mechanism for Steroid Receptor Translocation to the Plasma Membrane. J. Biol. Chem. 2007, 282, 22278–22288. [Google Scholar] [CrossRef] [Green Version]
- Razandi, M.; Pedram, A.; Levin, E.R. Heat Shock Protein 27 Is Required for Sex Steroid Receptor Trafficking to and Functioning at the Plasma Membrane. Mol. Cell. Biol. 2010, 30, 3249. [Google Scholar] [CrossRef] [Green Version]
- Shupnik, M.A. Crosstalk between Steroid Receptors and the C-Src-Receptor Tyrosine Kinase Pathways: Implications for Cell Proliferation. Oncogene 2004, 23, 7979–7989. [Google Scholar] [CrossRef] [Green Version]
- Simoncini, T.; Genazzani, A.R. Non-Genomic Actions of Sex Steroid Hormones. Eur. J. Endocrinol. 2003, 148, 281–292. [Google Scholar] [CrossRef] [Green Version]
- Moore, M.R.; Conover, J.L.; Franks, K.M. Progestin Effects on Long-Term Growth, Death, and Bcl-XL in Breast Cancer Cells. Biochem. Biophys. Res. Commun. 2000, 277, 650–654. [Google Scholar] [CrossRef]
- Lange, C.A.; Shen, T.; Horwitz, K.B. Phosphorylation of Human Progesterone Receptors at Serine-294 by Mitogen-Activated Protein Kinase Signals Their Degradation by the 26S Proteasome. Proc. Natl. Acad. Sci. USA 2000, 97, 1032–1037. [Google Scholar] [CrossRef] [Green Version]
- Skildum, A.; Faivre, E.; Lange, C.A. Progesterone Receptors Induce Cell Cycle Progression via Activation of Mitogen-Activated Protein Kinases. Mol. Endocrinol. 2005, 19, 327–339. [Google Scholar] [CrossRef] [Green Version]
- Faivre, E.J.; Daniel, A.R.; Hillard, C.J.; Lange, C.A. Progesterone Receptor Rapid Signaling Mediates Serine 345 Phosphorylation and Tethering to Specificity Protein 1 Transcription Factors. Mol. Endocrinol. 2008, 22, 823–837. [Google Scholar] [CrossRef] [Green Version]
- Proietti, C.; Salatino, M.; Rosemblit, C.; Carnevale, R.; Pecci, A.; Kornblihtt, A.R.; Molinolo, A.A.; Frahm, I.; Charreau, E.H.; Schillaci, R.; et al. Progestins Induce Transcriptional Activation of Signal Transducer and Activator of Transcription 3 (Stat3) via a Jak- and Src-Dependent Mechanism in Breast Cancer Cells. Mol. Cell. Biol. 2005, 25, 4826–4840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, S.K.; Hanson, D.A.; Schlaepfer, D.D. Focal Adhesion Kinase: In Command and Control of Cell Motility. Nat. Rev. Mol. Cell Biol. 2005, 6, 56–68. [Google Scholar] [CrossRef]
- Ostrom, Q.T.; Cioffi, G.; Waite, K.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2014–2018. Neuro. Oncol. 2021, 23, III1–III105. [Google Scholar] [CrossRef] [PubMed]
- McLendon, R.; Friedman, A.; Bigner, D.; Van Meir, E.G.; Brat, D.J.; Mastrogianakis, G.M.; Olson, J.J.; Mikkelsen, T.; Lehman, N.; Aldape, K.; et al. Comprehensive Genomic Characterization Defines Human Glioblastoma Genes and Core Pathways. Nature 2008, 455, 1061–1068. [Google Scholar] [CrossRef]
- Du, J.; Bernasconi, P.; Clauser, K.R.; Mani, D.R.; Finn, S.P.; Beroukhim, R.; Burns, M.; Julian, B.; Peng, X.P.; Hieronymus, H.; et al. Bead-Based Profiling of Tyrosine Kinase Phosphorylation Identifies SRC as a Potential Target for Glioblastoma Therapy. Nat. Biotechnol. 2009, 27, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Weissenberger, J.; Steinbach, J.P.; Malin, G.; Spada, S.; Rülicke, T.; Aguzzi, A. Development and Malignant Progression of Astrocytomas in GFAP-v-Src Transgenic Mice. Oncogene 1997, 14, 2005–2013. [Google Scholar] [CrossRef] [Green Version]
- Angers-Loustau, A.; Hering, R.; Werbowetski, T.E.; Kaplan, D.R.; Del Maestro, R.F. Src Regulates Actin Dynamics and Invasion of Malignant Glial Cells in Three Dimensions11Alex Pavanel and Franco Di Giovanni Funds for Brain Tumor Research (R.F. Del Maestro), Canadian Cancer Society and National Cancer Institute of Canada (D.R. Kaplan). Mol. Cancer Res. 2004, 2, 595–605. [Google Scholar] [CrossRef]
- Lewis-Tuffin, L.J.; Feathers, R.; Hari, P.; Durand, N.; Li, Z.; Rodriguez, F.J.; Bakken, K.; Carlson, B.; Schroeder, M.; Sarkaria, J.N.; et al. Src Family Kinases Differentially Influence Glioma Growth and Motility. Mol. Oncol. 2015, 9, 1783–1798. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Zhang, W.; Yang, X.; Wheeler, C.G.; Langford, C.P.; Wu, L.; Filippova, N.; Friedman, G.K.; Ding, Q.; Fathallah-Shaykh, H.M.; et al. The Role of Src Family Kinases in Growth and Migration of Glioma Stem Cells. Int. J. Oncol. 2014, 45, 302–310. [Google Scholar] [CrossRef] [Green Version]
- Jaraíz-Rodríguez, M.; Tabernero, M.D.; González-Tablas, M.; Otero, A.; Orfao, A.; Medina, J.M.; Tabernero, A. A Short Region of Connexin43 Reduces Human Glioma Stem Cell Migration, Invasion, and Survival through Src, PTEN, and FAK. Stem Cell Reports 2017, 9, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Galanis, E.; Anderson, S.K.; Twohy, E.L.; Carrero, X.W.; Dixon, J.G.; Tran, D.D.; Jeyapalan, S.A.; Anderson, D.M.; Kaufmann, T.J.; Feathers, R.W.; et al. A Phase I and Randomized Placebo-Controlled Phase II Trial of Bevacizumab plus Dasatinib in Patients with Recurrent Glioblastoma (GBM): Alliance/NCCTG N0872. Cancer 2019, 125, 3790. [Google Scholar] [CrossRef] [PubMed]
- Khalid, H.; Shibata, S.; Kishikawa, M.; Yasunaga, A.; Iseki, M.; Hiura, T. Immunohistochemical Analysis of Progesterone Receptor and Ki-67 Labeling Index in Astrocytic Tumors—PubMed. Cancer 1997, 80, 2133–2140. [Google Scholar] [CrossRef]
- Arcos-Montoya, D.; Wegman-Ostrosky, T.; Mejía-Pérez, S.; De la Fuente-Granada, M.; Camacho-Arroyo, I.; García-Carrancá, A.; Velasco-Velázquez, M.A.; Manjarrez-Marmolejo, J.; González-Arenas, A. Progesterone Receptor Together with PKCα Expression as Prognostic Factors for Astrocytomas Malignancy. Onco Targets Ther. 2021, 14, 3757. [Google Scholar] [CrossRef]
- González-Agüero, G.; Gutiérrez, A.A.; González-Espinosa, D.; Solano, J.D.; Morales, R.; González-Arenas, A.; Cabrera-Muñoz, E.; Camacho-Arroyo, I. Progesterone Effects on Cell Growth of U373 and D54 Human Astrocytoma Cell Lines. Endocrine 2007, 32, 129–135. [Google Scholar] [CrossRef]
- Piña-Medina, A.G.; Hansberg-Pastor, V.; González-Arenas, A.; Cerbón, M.; Camacho-Arroyo, I. Progesterone Promotes Cell Migration, Invasion and Cofilin Activation in Human Astrocytoma Cells. Steroids 2016, 105, 19–25. [Google Scholar] [CrossRef]
- Germán-Castelán, L.; Manjarrez-Marmolejo, J.; González-Arenas, A.; Genoveva González-Morán, M.; Camacho-Arroyo, I. Progesterone Induces the Growth and Infiltration of Human Astrocytoma Cells Implanted in the Cerebral Cortex of the Rat. BioMed Res. Int. 2014, 2014, 393174. [Google Scholar] [CrossRef]
- Bello-Alvarez, C.; Moral-Morales, A.D.; González-Arenas, A.; Camacho-Arroyo, I. Intracellular Progesterone Receptor and CSrc Protein Working Together to Regulate the Activity of Proteins Involved in Migration and Invasion of Human Glioblastoma Cells. Front. Endocrinol. 2021, 12, 640298. [Google Scholar] [CrossRef]
- Hernández-Hernández, O.T.; González-García, T.K.; Camacho-Arroyo, I. Progesterone Receptor and SRC-1 Participate in the Regulation of VEGF, EGFR and Cyclin D1 Expression in Human Astrocytoma Cell Lines. J. Steroid Biochem. Mol. Biol. 2012, 132, 127–134. [Google Scholar] [CrossRef]
- Trabert, B.; Sherman, M.E.; Kannan, N.; Stanczyk, F.Z. Progesterone and Breast Cancer. Endocr. Rev. 2020, 41, 320–344. [Google Scholar] [CrossRef]
- Stettner, M.R.; Wang, W.; Nabors, L.B.; Bharara, S.; Flynn, D.C.; Grammer, J.R.; Gillespie, G.Y.; Gladson, C.L. Lyn Kinase Activity Is the Predominant Cellular Src Kinase Activity in Glioblastoma Tumor Cells. Cancer Res. 2005, 65, 5535–5543. [Google Scholar] [CrossRef] [Green Version]
- Tavares, C.B.; Gomes-Braga, F.d.C.S.A.; Costa-Silva, D.R.; Escórcio-Dourado, C.S.; Borges, U.S.; Conde-Junior, A.M.; Barros-Oliveira, M.d.C.; Sousa, E.B.; Barros, L.d.R.; Martins, L.M.; et al. Expression of Estrogen and Progesterone Receptors in Astrocytomas: A Literature Review. Clinics 2016, 71, 481–486. [Google Scholar] [CrossRef]
- Hernández-Vega, A.M.; Del Moral-Morales, A.; Zamora-Sánchez, C.J.; Piña-Medina, A.G.; González-Arenas, A.; Camacho-Arroyo, I. Estradiol Induces Epithelial to Mesenchymal Transition of Human Glioblastoma Cells. Cells 2020, 9, 1930. [Google Scholar] [CrossRef] [PubMed]
- Castoria, G.; Giovannelli, P.; Lombardi, M.; De Rosa, C.; Giraldi, T.; De Falco, A.; Barone, M.V.; Abbondanza, C.; Migliaccio, A.; Auricchio, F. Tyrosine Phosphorylation of Estradiol Receptor by Src Regulates Its Hormone-Dependent Nuclear Export and Cell Cycle Progression in Breast Cancer Cells. Oncogene 2012, 31, 4868–4877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawprasertsri, S.; Pietras, R.J.; Marquez-Garban, D.C.; Boonyaratanakornkit, V. Progesterone Receptor (PR) Polyproline Domain (PPD) Mediates Inhibition of Epidermal Growth Factor Receptor (EGFR) Signaling in Non-Small Cell Lung Cancer Cells. Cancer Lett. 2016, 374, 279–291. [Google Scholar] [CrossRef] [Green Version]
- Oprita, A.; Baloi, S.C.; Staicu, G.A.; Alexandru, O.; Tache, D.E.; Danoiu, S.; Micu, E.S.; Sevastre, A.S. Updated Insights on EGFR Signaling Pathways in Glioma. Int. J. Mol. Sci. 2021, 22, 587. [Google Scholar] [CrossRef]
- Giaccone, G.; Zucali, P.A. Src as a Potential Therapeutic Target in Non-Small-Cell Lung Cancer. Ann. Oncol. 2008, 19, 1219–1223. [Google Scholar] [CrossRef]
- Rothschild, S.; Gautschi, O.; Haura, E.; Johnson, F. Src Inhibitors in Lung Cancer: Current Status and Future Directions. Clin. Lung Cancer 2010, 11, 238–242. [Google Scholar] [CrossRef]
- Jones, R.J.; Avizienyte, E.; Wyke, A.W.; Owens, D.W.; Brunton, V.G.; Frame, M.C. Elevated C-Src Is Linked to Altered Cell—Matrix Adhesion Rather than Proliferation in KM12C Human Colorectal Cancer Cells. Br. J. Cancer 2002, 87, 1128–1135. [Google Scholar] [CrossRef] [Green Version]
- Kopetz, S.; Kopetz, S. Targeting Src and Epidermal Growth Factor Receptor in Colorectal Cancer: Rationale and Progress Into the Clinic. Gastrointest. Cancer Res. 2007, 1, S37. [Google Scholar]
- Xu, H.; Zhang, L.; Yuan, M.; Xu, L.F.; Zhang, J.; Kong, S.; Wu, M.; Lao, Y. SRC and MEK Co-Inhibition Synergistically Enhances the Anti-Tumor Effect in Both Non-Small Cell Lung Cancer (NSCLC) and Erlotinib-Resistant NSCLC. Front. Oncol. 2019, 9, 586. [Google Scholar] [CrossRef]
- Zhang, J.; Kalyankrishna, S.; Wislez, M.; Thilaganathan, N.; Saigal, B.; Wei, W.; Ma, L.; Wistuba, I.I.; Johnson, F.M.; Kurie, J.M. Src-Family Kinases Are Activated in Non-Small Cell Lung Cancer and Promote the Survival of Epidermal Growth Factor Receptor-Dependent Cell Lines. Am. J. Pathol. 2007, 170, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishibashi, H.; Suzuki, T.; Suzuki, S.; Niikawa, H.; Lu, L.; Miki, Y.; Moriya, T.; Hayashi, S.I.; Handa, M.; Kondo, T.; et al. Progesterone Receptor in Non-Small Cell Lung Cancer--a Potent Prognostic Factor and Possible Target for Endocrine Therapy. Cancer Res. 2005, 65, 6450–6458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skjefstad, K.; Richardsen, E.; Donnem, T.; Andersen, S.; Kiselev, Y.; Grindstad, T.; Hald, S.M.; Al-Shibli, K.; Bremnes, R.M.; Busund, L.T.; et al. The Prognostic Role of Progesterone Receptor Expression in Non-Small Cell Lung Cancer Patients: Gender-Related Impacts and Correlation with Disease-Specific Survival. Steroids 2015, 98, 29–36. [Google Scholar] [CrossRef]
- Faivre, E.J.; Lange, C.A. Progesterone Receptors Upregulate Wnt-1 to Induce Epidermal Growth Factor Receptor Transactivation and c-Src-Dependent Sustained Activation of Erk1/2 Mitogen-Activated Protein Kinase in Breast Cancer Cells. Mol. Cell. Biol. 2007, 27, 466–480. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.T.; Lee, W. Sen Progesterone Receptor Activation Is Required for Folic Acid-Induced Anti-Proliferation in Colorectal Cancer Cell Lines. Cancer Lett. 2016, 378, 104–110. [Google Scholar] [CrossRef]
- Atif, F.; Yousuf, S.; Stein, D.G. Anti-Tumor Effects of Progesterone in Human Glioblastoma Multiforme: Role of PI3K/Akt/MTOR Signaling. J. Steroid Biochem. Mol. Biol. 2015, 146, 62–73. [Google Scholar] [CrossRef]
- Altinoz, M.A.; Ucal, Y.; Yilmaz, M.C.; Kiris, İ.; Ozisik, O.; Sezerman, U.; Ozpinar, A.; Elmaci, İ. Progesterone at High Doses Reduces the Growth of U87 and A172 Glioblastoma Cells: Proteomic Changes Regarding Metabolism and Immunity. Cancer Med. 2020, 9, 5767–5780. [Google Scholar] [CrossRef]
- Migliaccio, A.; Varricchio, L.; De Falco, A.; Castoria, G.; Arra, C.; Yamaguchi, H.; Ciociola, A.; Lombardi, M.; Di Stasio, R.; Barbieri, A.; et al. Inhibition of the SH3 Domain-Mediated Binding of Src to the Androgen Receptor and Its Effect on Tumor Growth. Oncogene 2007, 26, 6619–6629. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bello-Alvarez, C.; Zamora-Sánchez, C.J.; Camacho-Arroyo, I. Rapid Actions of the Nuclear Progesterone Receptor through cSrc in Cancer. Cells 2022, 11, 1964. https://doi.org/10.3390/cells11121964
Bello-Alvarez C, Zamora-Sánchez CJ, Camacho-Arroyo I. Rapid Actions of the Nuclear Progesterone Receptor through cSrc in Cancer. Cells. 2022; 11(12):1964. https://doi.org/10.3390/cells11121964
Chicago/Turabian StyleBello-Alvarez, Claudia, Carmen J. Zamora-Sánchez, and Ignacio Camacho-Arroyo. 2022. "Rapid Actions of the Nuclear Progesterone Receptor through cSrc in Cancer" Cells 11, no. 12: 1964. https://doi.org/10.3390/cells11121964