
The Tumor-Specific Expression of L1 Retrotransposons Independently Correlates with Time to Relapse in Hormone-Negative Breast Cancer Patients

 , , , , , , , , , and
, , , , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Breast Cancer Patient Cohort
2.3. Nucleic Acids Extraction
2.4. qRT-PCR of L1-ORF1 and ORF2 Expression
2.5. PAM50 Subtyping
2.6. Mutational Analyses
2.7. L1 Promoter Methylation Analysis
2.8. Statistical Analysis
3. Results
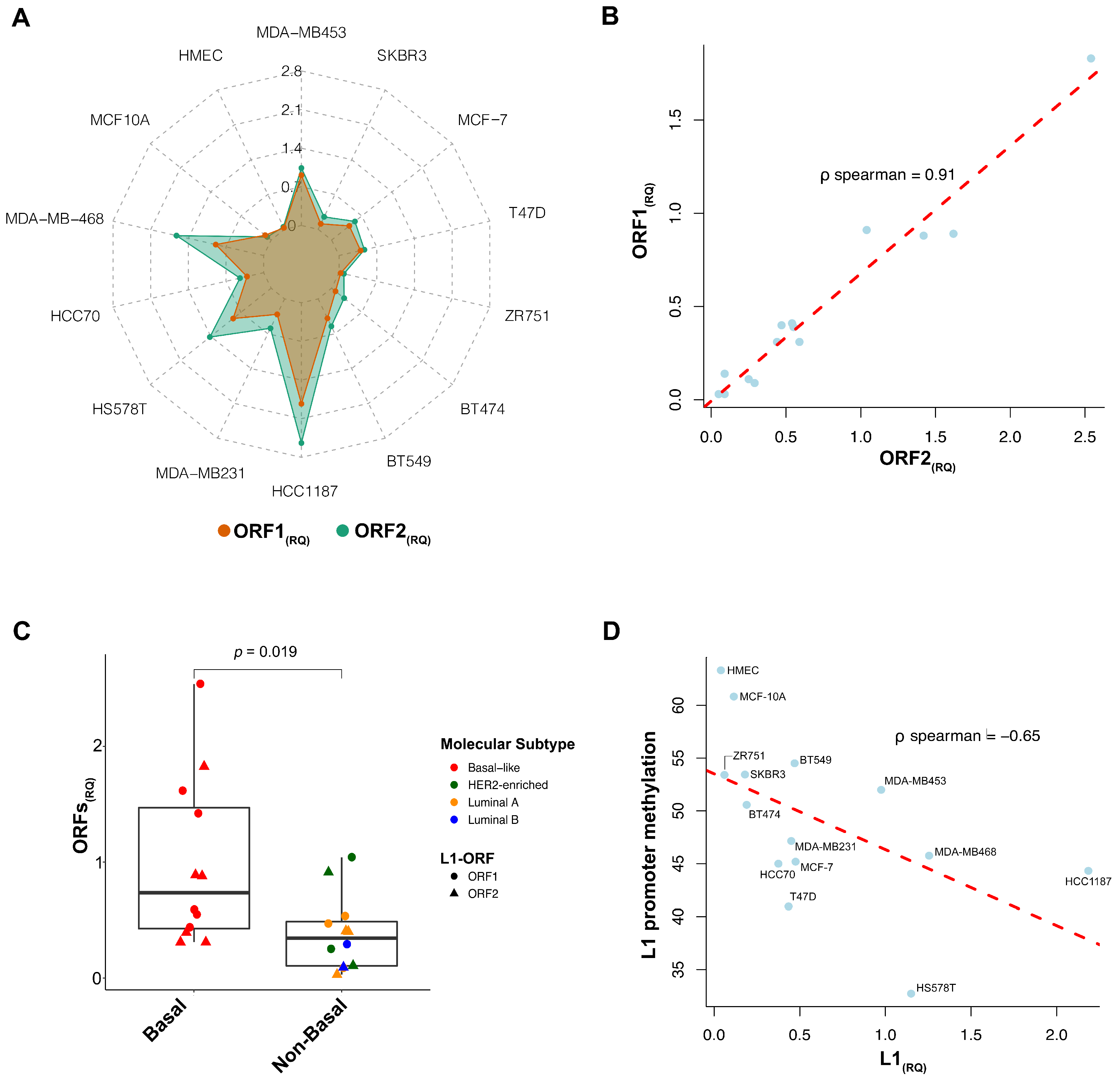
3.1. L1 Expression in Breast Cancer Cell Lines
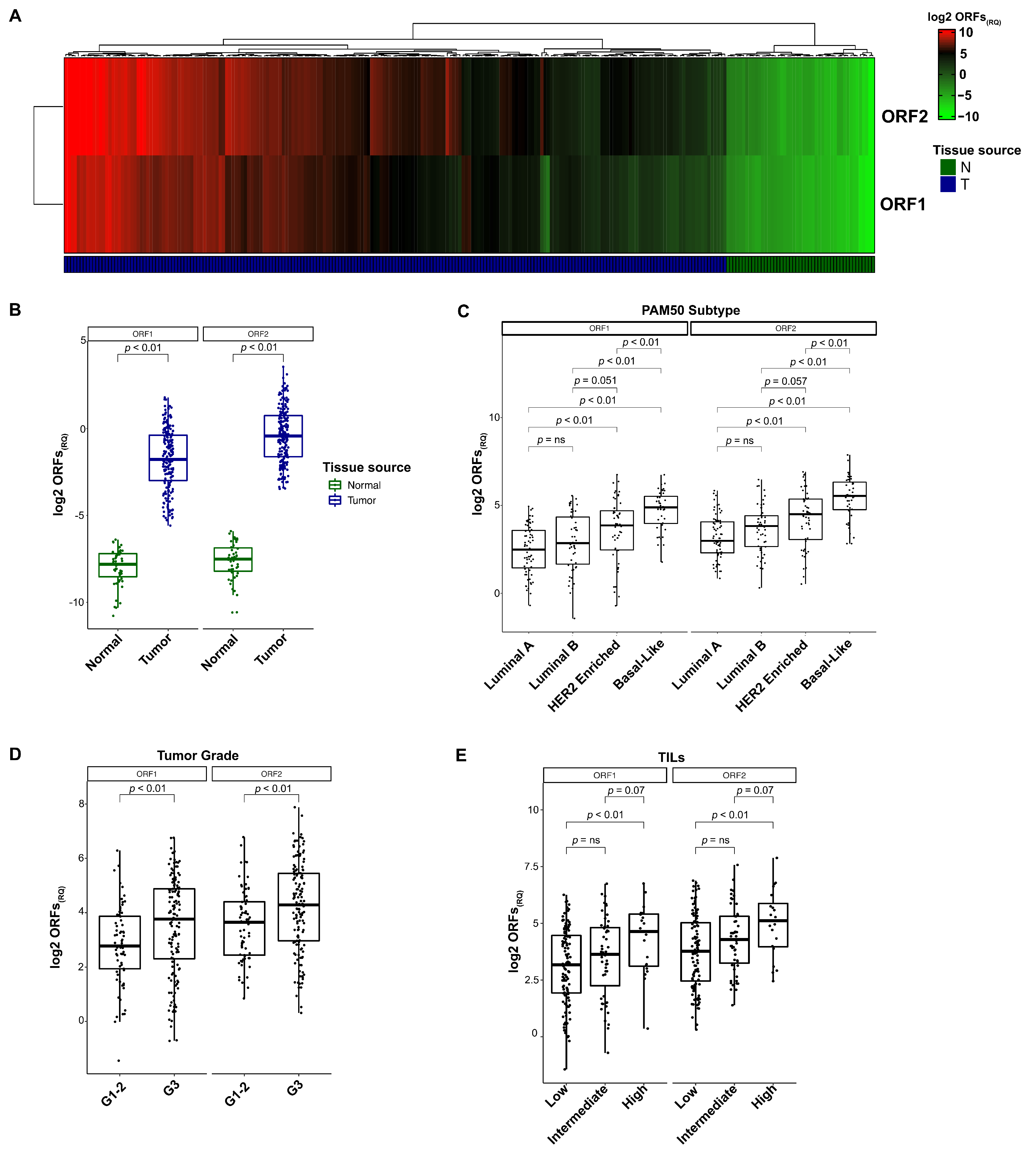
3.2. L1 Expression in Breast Cancer Tissues
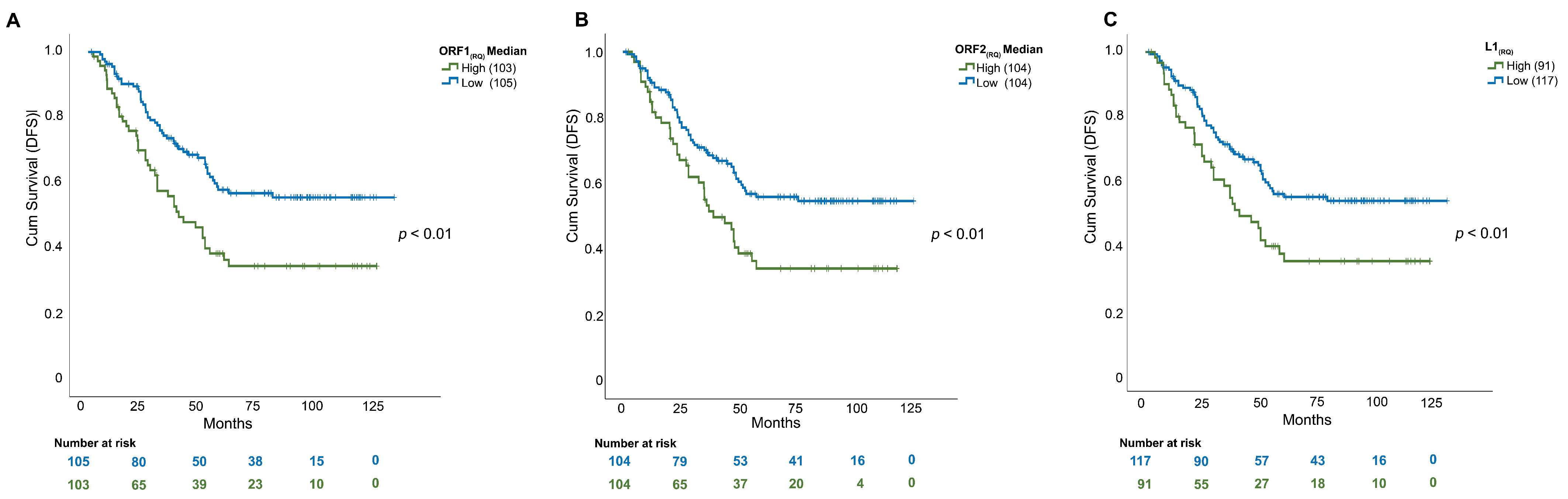
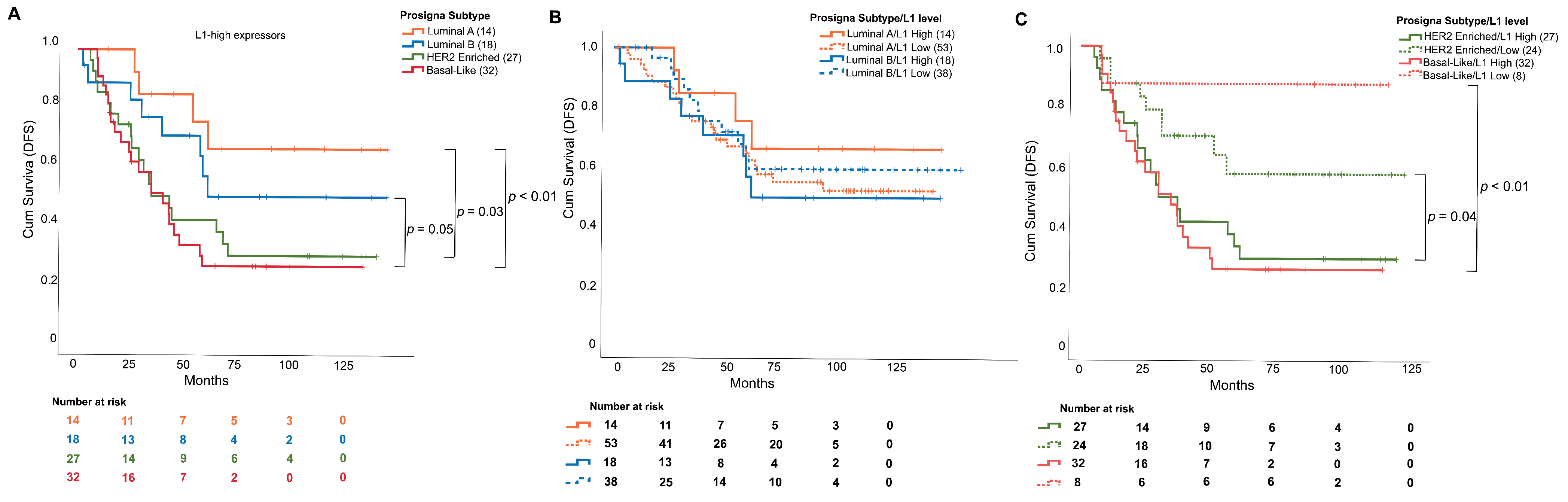
3.3. L1 mRNA Enrichment Was Associated with Relapse and Shorter Disease-Free Survival
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burns, K.H. Transposable elements in cancer. Nat. Rev. Cancer 2017, 17, 415–424. [Google Scholar] [CrossRef]
- Lee, H.; Zhang, Z.; Krause, H.M. Long Noncoding RNAs and Repetitive Elements: Junk or Intimate Evolutionary Partners? Trends Genet. 2019, 35, 892–902. [Google Scholar] [CrossRef] [Green Version]
- Mueller, C.; Aschacher, T.; Wolf, B.; Bergmann, M. A role of LINE-1 in telomere regulation. Front. Biosci. 2018, 23, 1310–1319. [Google Scholar] [CrossRef] [Green Version]
- Richardson, S.R.; Doucet, A.J.; Kopera, H.C.; Moldovan, J.B.; Garcia-Perez, J.L.; Moran, J.V. The Influence of LINE-1 and SINE Retrotransposons on Mammalian Genomes. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodic, N. LINE-1 activity and regulation in cancer. Front. Biosci. 2018, 23, 1680–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewing, A.D.; Smits, N.; Sanchez-Luque, F.J.; Faivre, J.; Brennan, P.M.; Richardson, S.R.; Cheetham, S.W.; Faulkner, G.J. Nanopore Sequencing Enables Comprehensive Transposable Element Epigenomic Profiling. Mol. Cell 2020, 80, 915–928.e915. [Google Scholar] [CrossRef] [PubMed]
- Kazazian, H.H., Jr.; Moran, J.V. Mobile DNA in Health and Disease. N. Engl. J. Med. 2017, 377, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Ponomaryova, A.A.; Rykova, E.Y.; Gervas, P.A.; Cherdyntseva, N.V.; Mamedov, I.Z.; Azhikina, T.L. Aberrant Methylation of LINE-1 Transposable Elements: A Search for Cancer Biomarkers. Cells 2020, 9, 2017. [Google Scholar] [CrossRef]
- Miki, Y.; Nishisho, I.; Horii, A.; Miyoshi, Y.; Utsunomiya, J.; Kinzler, K.W.; Vogelstein, B.; Nakamura, Y. Disruption of the APC gene by a retrotransposal insertion of L1 sequence in a colon cancer. Cancer Res. 1992, 52, 643–645. [Google Scholar]
- Morse, B.; Rotherg, P.G.; South, V.J.; Spandorfer, J.M.; Astrin, S.M. Insertional mutagenesis of the myc locus by a LINE-1 sequence in a human breast carcinoma. Nature 1988, 333, 87–90. [Google Scholar] [CrossRef]
- Ardeljan, D.; Taylor, M.S.; Ting, D.T.; Burns, K.H. The Human Long Interspersed Element-1 Retrotransposon: An Emerging Biomarker of Neoplasia. Clin. Chem. 2017, 63, 816–822. [Google Scholar] [CrossRef] [Green Version]
- Terry, D.M.; Devine, S.E. Aberrantly High Levels of Somatic LINE-1 Expression and Retrotransposition in Human Neurological Disorders. Front. Genet. 2019, 10, 1244. [Google Scholar] [CrossRef] [Green Version]
- Mita, P.; Boeke, J.D. Cycling to Maintain and Improve Fitness: Line-1 Modes of Nuclear Entrance and Retrotransposition. SLAS Discov. 2018, 23, 491–494. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Martin, B.; Alvarez, E.G.; Baez-Ortega, A.; Zamora, J.; Supek, F.; Demeulemeester, J.; Santamarina, M.; Ju, Y.S.; Temes, J.; Garcia-Souto, D.; et al. Pan-cancer analysis of whole genomes identifies driver rearrangements promoted by LINE-1 retrotransposition. Nat. Genet. 2020, 52, 306–319. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Dahlstrom, J.E.; Lee, S.H.; Rangasamy, D. Naturally occurring endo-siRNA silences LINE-1 retrotransposons in human cells through DNA methylation. Epigenetics 2012, 7, 758–771. [Google Scholar] [CrossRef]
- Patnala, R.; Lee, S.H.; Dahlstrom, J.E.; Ohms, S.; Chen, L.; Dheen, S.T.; Rangasamy, D. Inhibition of LINE-1 retrotransposon-encoded reverse transcriptase modulates the expression of cell differentiation genes in breast cancer cells. Breast Cancer Res. Treat. 2014, 143, 239–253. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Dahlstrom, J.E.; Chandra, A.; Board, P.; Rangasamy, D. Prognostic value of LINE-1 retrotransposon expression and its subcellular localization in breast cancer. Breast Cancer Res. Treat. 2012, 136, 129–142. [Google Scholar] [CrossRef] [Green Version]
- Petrelli, A.; Bellomo, S.E.; Sarotto, I.; Kubatzki, F.; Sgandurra, P.; Maggiorotto, F.; Di Virgilio, M.R.; Ponzone, R.; Geuna, E.; Galizia, D.; et al. MiR-100 is a predictor of endocrine responsiveness and prognosis in patients with operable luminal breast cancer. ESMO Open 2020, 5, e000937. [Google Scholar] [CrossRef]
- Salgado, R.; Denkert, C.; Demaria, S.; Sirtaine, N.; Klauschen, F.; Pruneri, G.; Wienert, S.; Van den Eynden, G.; Baehner, F.L.; Penault-Llorca, F.; et al. The evaluation of tumor-infiltrating lymphocytes (TILs) in breast cancer: Recommendations by an International TILs Working Group 2014. Ann. Oncol. 2015, 26, 259–271. [Google Scholar] [CrossRef]
- Gnant, M.; Sestak, I.; Filipits, M.; Dowsett, M.; Balic, M.; Lopez-Knowles, E.; Greil, R.; Dubsky, P.; Stoeger, H.; Rudas, M.; et al. Identifying clinically relevant prognostic subgroups of postmenopausal women with node-positive hormone receptor-positive early-stage breast cancer treated with endocrine therapy: A combined analysis of ABCSG-8 and ATAC using the PAM50 risk of recurrence score and intrinsic subtype. Ann. Oncol. 2015, 26, 1685–1691. [Google Scholar] [CrossRef]
- Laenkholm, A.V.; Jensen, M.B.; Eriksen, J.O.; Rasmussen, B.B.; Knoop, A.S.; Buckingham, W.; Ferree, S.; Schaper, C.; Nielsen, T.O.; Haffner, T.; et al. PAM50 Risk of Recurrence Score Predicts 10-Year Distant Recurrence in a Comprehensive Danish Cohort of Postmenopausal Women Allocated to 5 Years of Endocrine Therapy for Hormone Receptor-Positive Early Breast Cancer. J. Clin. Oncol. 2018, 36, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, T.; Wallden, B.; Schaper, C.; Ferree, S.; Liu, S.; Gao, D.; Barry, G.; Dowidar, N.; Maysuria, M.; Storhoff, J. Analytical validation of the PAM50-based Prosigna Breast Cancer Prognostic Gene Signature Assay and nCounter Analysis System using formalin-fixed paraffin-embedded breast tumor specimens. BMC Cancer 2014, 14, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, J.S.; Mullins, M.; Cheang, M.C.; Leung, S.; Voduc, D.; Vickery, T.; Davies, S.; Fauron, C.; He, X.; Hu, Z.; et al. Supervised risk predictor of breast cancer based on intrinsic subtypes. J. Clin. Oncol. 2009, 27, 1160–1167. [Google Scholar] [CrossRef] [PubMed]
- Wallden, B.; Storhoff, J.; Nielsen, T.; Dowidar, N.; Schaper, C.; Ferree, S.; Liu, S.; Leung, S.; Geiss, G.; Snider, J.; et al. Development and verification of the PAM50-based Prosigna breast cancer gene signature assay. BMC Med. Genom. 2015, 8, 54. [Google Scholar] [CrossRef] [Green Version]
- Marchio, C.; Dell’Orto, P.; Annaratone, L.; Geyer, F.C.; Venesio, T.; Berrino, E.; Verdun di Cantogno, L.; Garofoli, A.; Rangel, N.; Casorzo, L.; et al. The Dilemma of HER2 Double-Equivocal Breast Carcinomas: Genomic Profiling and Implications for Treatment. Am. J. Surg. Pathol. 2018, 42, 1190–1200. [Google Scholar] [CrossRef]
- Debernardi, C.; Libera, L.; Berrino, E.; Sahnane, N.; Chiaravalli, A.M.; Laudi, C.; Berselli, M.; Sapino, A.; Sessa, F.; Venesio, T.; et al. Evaluation of global and intragenic hypomethylation in colorectal adenomas improves patient stratification and colorectal cancer risk prediction. Clin. Epigenet. 2021, 13, 154. [Google Scholar] [CrossRef]
- Dong, B.; Ding, Y.; Huang, Q.; Guan, X. Different Triple-Negative Breast Cancer Tumor Cell Lysates (TCLs) Induce Discrepant Anti-Tumor Immunity by PD1/PDL-1 Interaction. Med. Sci. Monit. 2019, 25, 500–515. [Google Scholar] [CrossRef]
- Smith, S.E.; Mellor, P.; Ward, A.K.; Kendall, S.; McDonald, M.; Vizeacoumar, F.S.; Vizeacoumar, F.J.; Napper, S.; Anderson, D.H. Molecular characterization of breast cancer cell lines through multiple omic approaches. Breast Cancer Res. 2017, 19, 65. [Google Scholar] [CrossRef] [Green Version]
- Sommers, C.L.; Byers, S.W.; Thompson, E.W.; Torri, J.A.; Gelmann, E.P. Differentiation state and invasiveness of human breast cancer cell lines. Breast Cancer Res. Treat. 1994, 31, 325–335. [Google Scholar] [CrossRef]
- Furlan, D.; Trapani, D.; Berrino, E.; Debernardi, C.; Panero, M.; Libera, L.; Sahnane, N.; Riva, C.; Tibiletti, M.G.; Sessa, F.; et al. Oxidative DNA damage induces hypomethylation in a compromised base excision repair colorectal tumourigenesis. Br. J. Cancer 2017, 116, 793–801. [Google Scholar] [CrossRef] [Green Version]
- Zeggar, H.R.; How-Kit, A.; Daunay, A.; Bettaieb, I.; Sahbatou, M.; Rahal, K.; Adouni, O.; Gammoudi, A.; Douik, H.; Deleuze, J.F.; et al. Tumor DNA hypomethylation of LINE-1 is associated with low tumor grade of breast cancer in Tunisian patients. Oncol. Lett. 2020, 20, 1999–2006. [Google Scholar] [CrossRef]
- Yang, F.; Wang, P.J. Multiple LINEs of retrotransposon silencing mechanisms in the mammalian germline. Semin. Cell Dev. Biol. 2016, 59, 118–125. [Google Scholar] [CrossRef] [Green Version]
- Belgnaoui, S.M.; Gosden, R.G.; Semmes, O.J.; Haoudi, A. Human LINE-1 retrotransposon induces DNA damage and apoptosis in cancer cells. Cancer Cell Int. 2006, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Kemp, J.R.; Longworth, M.S. Crossing the LINE Toward Genomic Instability: LINE-1 Retrotransposition in Cancer. Front. Chem. 2015, 3, 68. [Google Scholar] [CrossRef] [Green Version]
- Pisanic, T.R., 2nd; Asaka, S.; Lin, S.F.; Yen, T.T.; Sun, H.; Bahadirli-Talbott, A.; Wang, T.H.; Burns, K.H.; Wang, T.L.; Shih, I.M. Long Interspersed Nuclear Element 1 Retrotransposons Become Deregulated during the Development of Ovarian Cancer Precursor Lesions. Am. J. Pathol. 2019, 189, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Henry, N.L.; Hayes, D.F. Cancer biomarkers. Mol. Oncol. 2012, 6, 140–146. [Google Scholar] [CrossRef] [Green Version]
- Chalitchagorn, K.; Shuangshoti, S.; Hourpai, N.; Kongruttanachok, N.; Tangkijvanich, P.; Thong-ngam, D.; Voravud, N.; Sriuranpong, V.; Mutirangura, A. Distinctive pattern of LINE-1 methylation level in normal tissues and the association with carcinogenesis. Oncogene 2004, 23, 8841–8846. [Google Scholar] [CrossRef] [Green Version]
- Budania, S.; Sur, D.; Nangal, J.; Pilli, S.; Mukherjee, K.; Biswas, M.; Prasad, R.; Saxena, S.; Mandal, P.K. LINE-1 retrotransposon encoded ORF1p expression and promoter methylation in oral squamous cell carcinoma: A pilot study. Cancer Genet. 2020, 244, 21–29. [Google Scholar] [CrossRef]
- Vafadar-Isfahani, N.; Parr, C.; McMillan, L.E.; Sanner, J.; Yeo, Z.; Saddington, S.; Peacock, O.; Cruickshanks, H.A.; Meehan, R.R.; Lund, J.N.; et al. Decoupling of DNA methylation and activity of intergenic LINE-1 promoters in colorectal cancer. Epigenetics 2017, 12, 465–475. [Google Scholar] [CrossRef]
- Kim, S.; Park, C.; Ji, Y.; Kim, D.G.; Bae, H.; van Vrancken, M.; Kim, D.H.; Kim, K.M. Deamination Effects in Formalin-Fixed, Paraffin-Embedded Tissue Samples in the Era of Precision Medicine. J. Mol. Diagn. 2017, 19, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Penzkofer, T.; Dandekar, T.; Zemojtel, T. L1Base: From functional annotation to prediction of active LINE-1 elements. Nucleic Acids Res. 2005, 33, D498–D500. [Google Scholar] [CrossRef] [Green Version]
- Miglio, U.; Berrino, E.; Panero, M.; Ferrero, G.; Coscujuela Tarrero, L.; Miano, V.; Dell’Aglio, C.; Sarotto, I.; Annaratone, L.; Marchio, C.; et al. The expression of LINE1-MET chimeric transcript identifies a subgroup of aggressive breast cancers. Int. J. Cancer 2018, 143, 2838–2848. [Google Scholar] [CrossRef] [Green Version]
- Beck, C.R.; Garcia-Perez, J.L.; Badge, R.M.; Moran, J.V. LINE-1 elements in structural variation and disease. Annu. Rev. Genom. Hum. Genet. 2011, 12, 187–215. [Google Scholar] [CrossRef] [Green Version]
- Van Hoesel, A.Q.; van de Velde, C.J.; Kuppen, P.J.; Liefers, G.J.; Putter, H.; Sato, Y.; Elashoff, D.A.; Turner, R.R.; Shamonki, J.M.; de Kruijf, E.M.; et al. Hypomethylation of LINE-1 in primary tumor has poor prognosis in young breast cancer patients: A retrospective cohort study. Breast Cancer Res. Treat. 2012, 134, 1103–1114. [Google Scholar] [CrossRef]
- Chiou, P.T.; Ohms, S.; Board, P.G.; Dahlstrom, J.E.; Rangasamy, D.; Casarotto, M.G. Efavirenz as a potential drug for the treatment of triple-negative breast cancers. Clin. Transl. Oncol. 2021, 23, 353–363. [Google Scholar] [CrossRef]
- McKerrow, W.; Wang, X.; Mendez-Dorantes, C.; Mita, P.; Cao, S.; Grivainis, M.; Ding, L.; LaCava, J.; Burns, K.H.; Boeke, J.D.; et al. LINE-1 expression in cancer correlates with p53 mutation, copy number alteration, and S phase checkpoint. Proc. Natl. Acad. Sci. USA 2022, 119, e2115999119. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, R.; Yu, J. New Understanding of the Relevant Role of LINE-1 Retrotransposition in Human Disease and Immune Modulation. Front. Cell Dev. Biol. 2020, 8, 657. [Google Scholar] [CrossRef]
- Park, J.H.; Jonas, S.F.; Bataillon, G.; Criscitiello, C.; Salgado, R.; Loi, S.; Viale, G.; Lee, H.J.; Dieci, M.V.; Kim, S.B.; et al. Prognostic value of tumor-infiltrating lymphocytes in patients with early-stage triple-negative breast cancers (TNBC) who did not receive adjuvant chemotherapy. Ann. Oncol. 2019, 30, 1941–1949. [Google Scholar] [CrossRef]
- Cortes, J.; Cescon, D.W.; Rugo, H.S.; Nowecki, Z.; Im, S.A.; Yusof, M.M.; Gallardo, C.; Lipatov, O.; Barrios, C.H.; Holgado, E.; et al. Pembrolizumab plus chemotherapy versus placebo plus chemotherapy for previously untreated locally recurrent inoperable or metastatic triple-negative breast cancer (KEYNOTE-355): A randomised, placebo-controlled, double-blind, phase 3 clinical trial. Lancet 2020, 396, 1817–1828. [Google Scholar] [CrossRef]
- Schmid, P.; Adams, S.; Rugo, H.S.; Schneeweiss, A.; Barrios, C.H.; Iwata, H.; Dieras, V.; Hegg, R.; Im, S.A.; Shaw Wright, G.; et al. Atezolizumab and Nab-Paclitaxel in Advanced Triple-Negative Breast Cancer. N. Engl. J. Med. 2018, 379, 2108–2121. [Google Scholar] [CrossRef]
- Annaratone, L.; Cascardi, E.; Vissio, E.; Sarotto, I.; Chmielik, E.; Sapino, A.; Berrino, E.; Marchio, C. The Multifaceted Nature of Tumor Microenvironment in Breast Carcinomas. Pathobiology 2020, 87, 125–142. [Google Scholar] [CrossRef]
- Emens, L.A.; Molinero, L.; Loi, S.; Rugo, H.S.; Schneeweiss, A.; Dieras, V.; Iwata, H.; Barrios, C.H.; Nechaeva, M.; Nguyen-Duc, A.; et al. Atezolizumab and nab-Paclitaxel in Advanced Triple-Negative Breast Cancer: Biomarker Evaluation of the IMpassion130 Study. J. Natl. Cancer Inst. 2021, 113, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Marasca, F.; Sinha, S.; Vadala, R.; Polimeni, B.; Ranzani, V.; Paraboschi, E.M.; Burattin, F.V.; Ghilotti, M.; Crosti, M.; Negri, M.L.; et al. LINE1 are spliced in non-canonical transcript variants to regulate T cell quiescence and exhaustion. Nat. Genet. 2022, 54, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.L.; Allen, C.; Henson, D.E. Relation of tumor size, lymph node status, and survival in 24,740 breast cancer cases. Cancer 1989, 63, 181–187. [Google Scholar] [CrossRef]
- Kanno, M.; Nakamura, S.; Uotani, C.; Yamanaka, S.; Terasaki, Y.; Tsugawa, K.; Noguchi, M. Prognosis of node-positive breast cancer patients who underwent parasternal lymph node biopsy during surgery followed by doxorubicin- or mitoxantrone-containing adjuvant chemotherapy. J. Chemother. 2000, 12, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Tonellotto, F.; Bergmann, A.; de Souza Abrahao, K.; de Aguiar, S.S.; Bello, M.A.; Thuler, L.C.S. Impact of Number of Positive Lymph Nodes and Lymph Node Ratio on Survival of Women with Node-Positive Breast Cancer. Eur. J. Breast Health 2019, 15, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Si, X.; Zhang, E.; Zang, R.; Yang, N.; Cheng, H.; Zhang, Z.; Pan, B.; Sun, Y. Paclitaxel-induced stress granules increase LINE-1 mRNA stability to promote drug resistance in breast cancer cells. J. Biomed. Res. 2021, 35, 411–424. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # of Patients (%) | ORF1(RQ) Mean (SD) | p-Value | ORF2(RQ) Mean ± SD | p-Value | ||
|---|---|---|---|---|---|---|
| PAM50 molecular subtype (n = 210) | Luminal A | 67 (32%) | 8.33 (7.6) | p < 0.0001 | 12.99 ± 12.1 | p < 0.0001 |
| Luminal B | 51 (24%) | 12.93 (12.0) | 19.13 ± 18.2 | |||
| HER2-E | 52 (25%) | 21.86 (18.3) | 31.12 ± 28.3 | |||
| Basal-like | 40 (19%) | 34.04 (23.5) | 58.76 ± 40.2 | |||
| Histotype (n = 210) | Ductal | 16 (8%) | 19.01 (16.6) | p = 0.34 | 28.03 (26.2) | p = 0.07 |
| Lobular | 171 (81%) | 8.30 (5.3) | 12.08 (10.4) | |||
| Other | 23 (11%) | 14.40 (12.8) | 23.62 (21.8) | |||
| Ki67 (n = 210) | Ki67 < 20% | 48 (23%) | 10.30 (8.9) | p = 0.0024 | 13.38 (11.9) | p = 0.0005 |
| ki67 > 20% | 162 (77%) | 19.91 (14.2) | 31.93 (25.3) | |||
| Tumor size (n = 210) | <20 mm | 71 (34%) | 14.79 (11.4) | p = 0.11 | 24.08 (21.8) | p = 0.23 |
| >20 mm | 139 (66%) | 19.21 (16.8) | 29.69 (22.9) | |||
| Grade (n = 210) | G1–G2 | 69 (33%) | 11.21 (9.9) | p = 0.0006 | 17.80 (16.4) | p = 0.0019 |
| G3 | 141 (67%) | 20.87 (17.0) | 32.53 (26.3) | |||
| TILs (n = 210) | Low (0–10%) | 127 (60%) | 15.23 (14.80) | p = 0.02 | 23.78 (17.1) | p = 0.01 |
| Intermediate (10–30%) | 58 (28%) | 20.73 (18.2) | 31.41 (24.9) | |||
| High (>30%) | 25 (12%) | 30.28 (26.5) | 51.29 (46.95) | |||
| Node (n = 210) | Negative | 70 (33%) | 14.24 (13.8) | p = 0.15 | 23.15 (21.2) | p = 0.38 |
| 1–3 nodes | 90 (43%) | 18.51 (17.3) | 30.43 (29.3) | |||
| >3 nodes | 50 (24%) | 21.00 (17.5) | 29.97 (27.4) | |||
| Relapse (n = 208) | Yes | 112 (54%) | 21.96 (20.7) | p = 0.0034 | 34.02 (30.9) | p = 0.0096 |
| No | 96(46%) | 14.13 (13.0) | 22.33 (19.6) |
| DFS | ||||||
|---|---|---|---|---|---|---|
| Univariate Analysis | Multivariate Analysis | |||||
| Median Months (95% CI) | HR (95% CI) | p-Value | HR (95% CI) | p-Value | ||
| Grade | G1–G2 | 54 (1.4 to 128.4) | 1 | 1 | ||
| G3 | 44 (2.5 to 120.9) | 1.7 (1.1 to 2.6) | 0.0089 | 1.3 (0.8 to 2.2) | 0.301 | |
| Histotype | Ductal | 44 (1.4 to 121.1) | 1 | 1 | ||
| Lobular | 47 (2.1 to 118.2) | 1.0 (0.5 to 2.0) | 0.983 | 1.2 (0.5 to 2.5) | 0.746 | |
| Other | 41 (8.8 to 128.4) | 1.0 (0.5 to 2.0) | 0.934 | 1.1 (0.5 to 2.4) | 0.634 | |
| Ki67 | Ki67 < 20% | 85 (1.4 to 118.2) | 1 | 1 | ||
| ki67 > 20% | 42 (2.1 to 128.4) | 1.3 (0.8 to 2.1) | 0.260 | 1.1 (0.6 to 2.1) | 0.501 | |
| Node | Negative | 48 (7.7 to 128.4) | 1 | 1 | ||
| 1–3 nodes | 52 (1.4 to 119.7) | 1.0 (0.5 to 1.6) | 0.944 | 0.9 (0.5 to 1.5) | 0.702 | |
| >3 nodes | 29 (5.0 to 118.2) | 2.9 (1.7 to 4.9) | 0.0001 | 2.3 (1.4 to 3.9) | 0.002 | |
| PAM50 molecular subtype | Luminal A | 51 (1.4 to 121.1) | 1 | 1 | ||
| Luminal B | 49 (2.1 to 128.4) | 1.0 (0.6 to 1.8) | 0.996 | 0.8 (0.4 to 1.6) | 0.528 | |
| HER2-E | 37 (5.0 to 119.7) | 1.5 (0.9 to 2.7) | 0.101 | 1.1 (0.6 to 2.3) | 0.710 | |
| Basal-like | 34 (7.6 to 113.6) | 2.0 (1.1 to 3.7) | 0.027 | 1.3 (0.6 to 2.7) | 0.568 | |
| TILs | Low (0–10%) | 41 (1.4 to 128.4) | 1 | 1 | ||
| Intermediate (10–30%) | 53 (6.0 to 120.9) | 1.0 (0.6 to 1.7) | 0.907 | 1.1 (0.6 to 1.7) | 0.820 | |
| High (>30%) | 49 (5.0 to 111.9) | 1.3 (0.6 to 2.6) | 0.491 | 0.9 (0.4 to 1.8) | 0.840 | |
| Tumor Size | <20 mm | 54 (1.4 to 121.1) | 1 | 1 | ||
| >20 mm | 38 (2.1 to 128.4) | 1.7 (1.1 to 2.5) | 0.020 | 1.5 (0.9 to 2.5) | 0.067 | |
| ORF1(RQ) Mean | Low | 49 (1.4 to 128.4) | 1 | 1 | ||
| High | 31 (2.1 to 121.2) | 2 (1.3 to 3.1) | 0.0017 | 1.7 (1.1 to 2.56) | 0.012 | |
| ORF1(RQ) Median | Low | 49 (1.4 to 128.4) | 1 | 1 | ||
| High | 37 (2.1 to 121.1) | 1.7 (1.1 to 2.5) | 0.0172 | 1.2 (1.1 to 1.5) | 0.043 | |
| ORF2(RQ) Mean | Low | 49 (1.4 to 128.4) | 1 | 1 | ||
| High | 36 (2.5 to 121.1) | 1.9 (1.2 to 2.9) | 0.0065 | 1.2 (1.1 to 1.5) | 0.037 | |
| ORF2(RQ) Median | Low | 51 (1.4 to 128.4) | 1 | 1 | ||
| High | 36 (2.1 to 121.1) | 1.8 (1.4 to 2.5) | 0.0097 | 1.3 (1.1 to 1.6) | 0.020 | |
| L1(RQ) Median | Low | 50 (1.4 to 128.4) | 1 | 1 | ||
| High | 35 (2.1 to 121.1) | 2 (1.3 to 3.0) | 0.0014 | 1.8 (1.2 to 2.7) | 0.005 | |
| L1(RQ) Mean | Low | 48 (1.4 to 128.4) | 1 | 1 | ||
| High | 36 (2.5 to 121.1) | 1.8 (1.1 to 2.9) | 0.0089 | 1.4 (1.1 to 1.7) | 0.038 | |
| DFS | |||
|---|---|---|---|
| Univariate analysis | |||
| PAM50 molecular subtype | Median months (95% CI) | HR (95% CI) | p-value |
| Luminal A | 54 (11.9 to 121.1) | 1 | |
| Luminal B | 39 (2.1 to 120.9) | 1.7 (0.5 to 5.4) | 0.363 |
| HER2-E | 28 (5.0 to 116.9) | 2.6 (1.1 to 6.1) | 0.027 |
| Basal-like | 28 (7.6 to 111.7) | 2.9 (1.3 to 6.4) | 0.0096 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berrino, E.; Miglio, U.; Bellomo, S.E.; Debernardi, C.; Bragoni, A.; Petrelli, A.; Cascardi, E.; Giordano, S.; Montemurro, F.; Marchiò, C.; et al. The Tumor-Specific Expression of L1 Retrotransposons Independently Correlates with Time to Relapse in Hormone-Negative Breast Cancer Patients. Cells 2022, 11, 1944. https://doi.org/10.3390/cells11121944
Berrino E, Miglio U, Bellomo SE, Debernardi C, Bragoni A, Petrelli A, Cascardi E, Giordano S, Montemurro F, Marchiò C, et al. The Tumor-Specific Expression of L1 Retrotransposons Independently Correlates with Time to Relapse in Hormone-Negative Breast Cancer Patients. Cells. 2022; 11(12):1944. https://doi.org/10.3390/cells11121944
Chicago/Turabian StyleBerrino, Enrico, Umberto Miglio, Sara Erika Bellomo, Carla Debernardi, Alberto Bragoni, Annalisa Petrelli, Eliano Cascardi, Silvia Giordano, Filippo Montemurro, Caterina Marchiò, and et al. 2022. "The Tumor-Specific Expression of L1 Retrotransposons Independently Correlates with Time to Relapse in Hormone-Negative Breast Cancer Patients" Cells 11, no. 12: 1944. https://doi.org/10.3390/cells11121944