
T-Cell Defects Associated to Lack of Spike-Specific Antibodies after BNT162b2 Full Immunization Followed by a Booster Dose in Patients with Common Variable Immune Deficiencies

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Patients
2.2. Anti-Spike IgG and IgA antibodies
2.3. Cell Isolation and Cryopreservation, T and B Cell Phenotypes
2.4. Functional Studies
2.5. Statistical Analysis
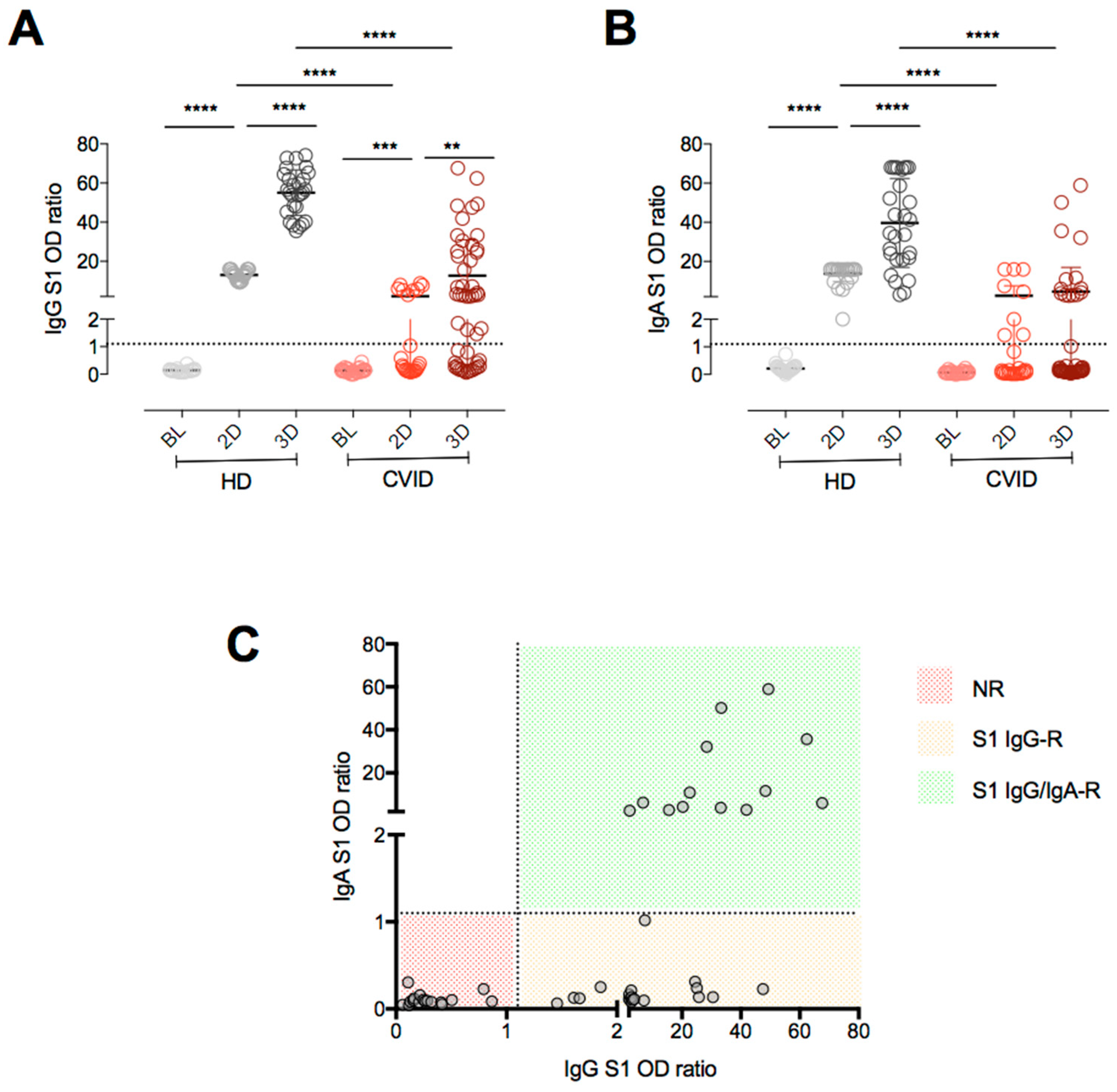
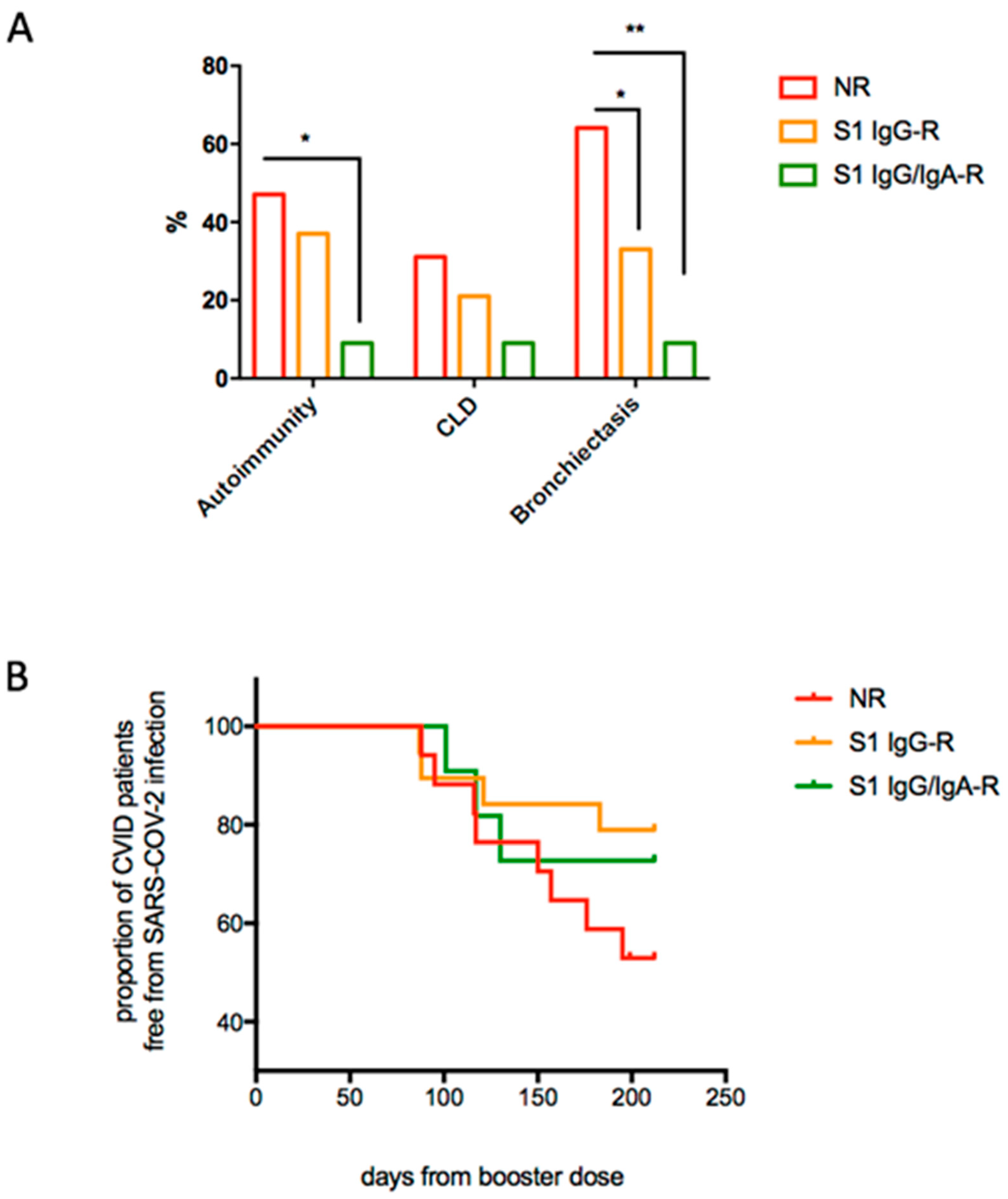
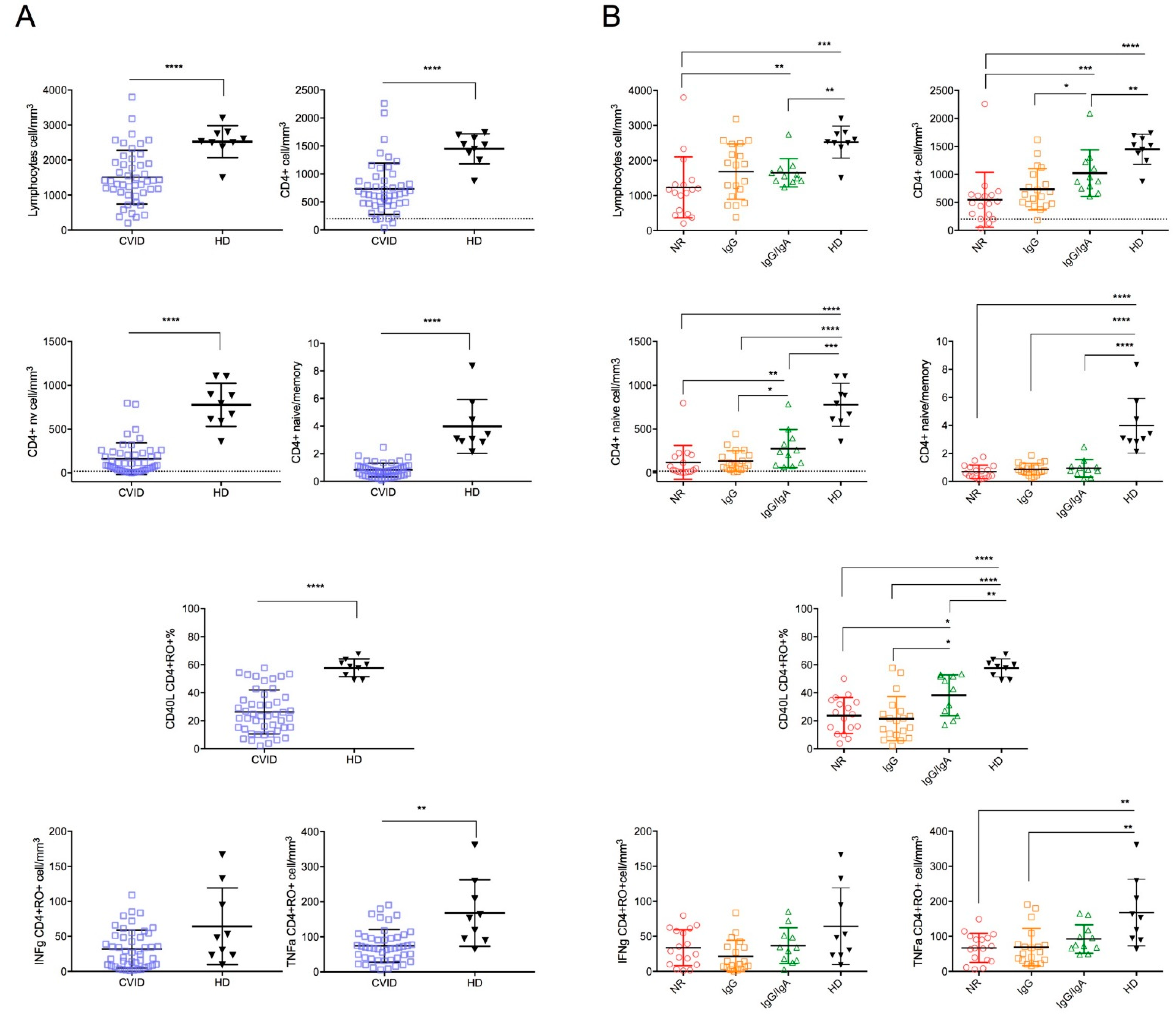
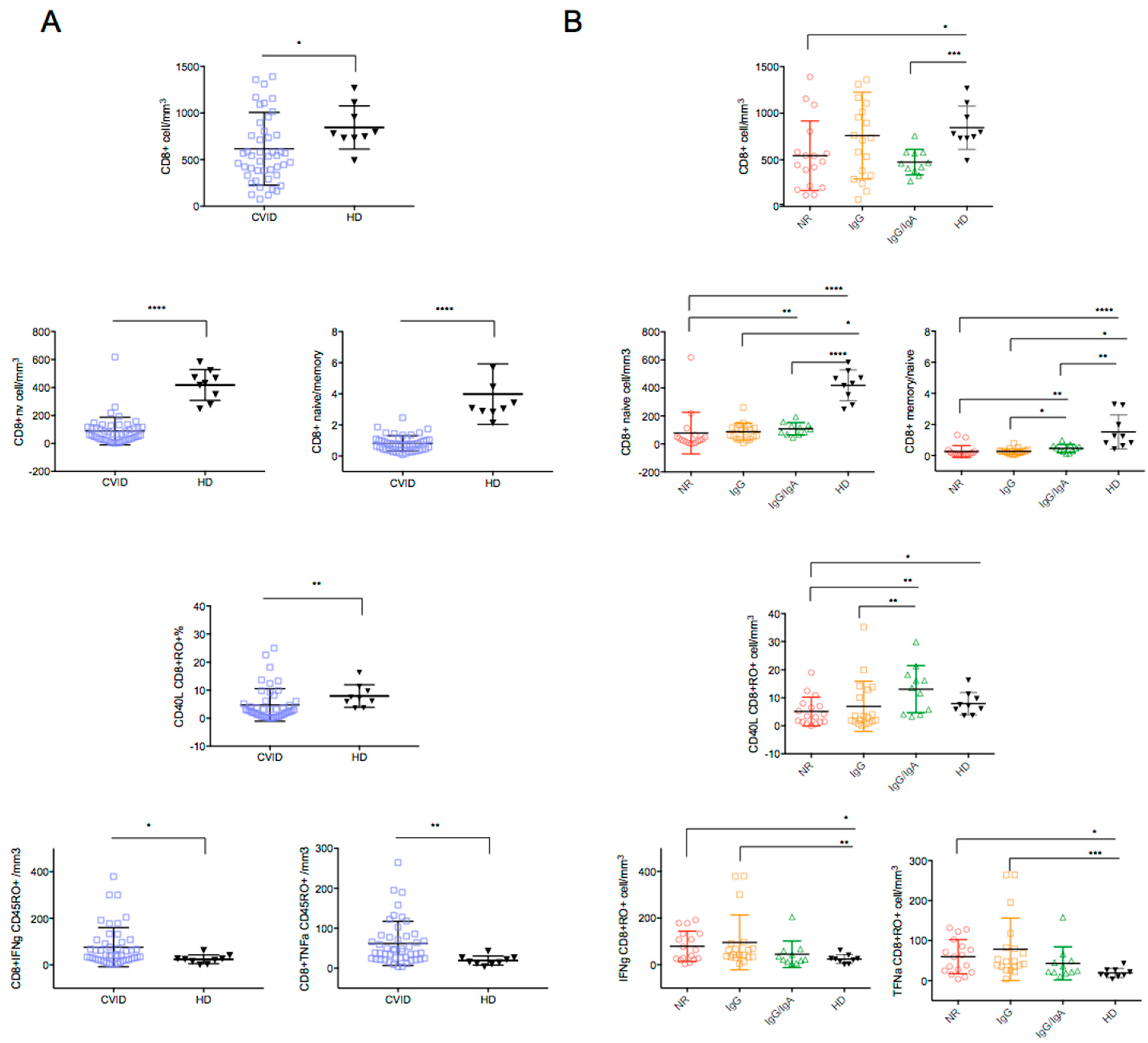
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tangye, S.G.; Al-Herz, W.; Bousfiha, A.; Chatila, T.; Cunningham-Rundles, C.; Etzioni, A.; Franco, J.L.; Holland, S.M.; Klein, C.; Morio, T.; et al. Human Inborn Errors of Immunity: 2019 Update on the Classification from the International Union of Immunological Societies Expert Committee. J. Clin. Immunol. 2020, 1, 24–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyts, I.; Bucciol, G.; Quinti, I.; Neven, B.; Fischer, A.; Seoane, E.; Lopez-Granados, E.; Gianelli, C.; Robles-Marhuenda, A.; Jeandel, P.Y.; et al. Coronavirus disease 2019 in patients with inborn errors of immunity: An international study. J. Allergy Clin. Immunol. 2021, 2, 520–531. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Chen, Y.; Liu, M.; Shi, S.; Tian, J. Impacts of immunosuppression and immunodeficiency on COVID-19: A systematic review and meta-analysis. J. Infect. 2020, 81, e93–e95. [Google Scholar] [CrossRef] [PubMed]
- Salinas, A.F.; Mortari, E.P.; Terreri, S.; Quintarelli, C.; Pulvirenti, F.; Di Cecca, S.; Guercio, M.; Milito, C.; Bonanni, L.; Auria, S.; et al. SARS-CoV-2 Vaccine Induced Atypical Immune Responses in Antibody Defects: Everybody Does their Best. J. Clin. Immunol. 2021, 8, 1709–1722. [Google Scholar] [CrossRef]
- Pulvirenti, F.; Fernandez Salinas, A.; Milito, C.; Terreri, S.; Piano Mortari, E.; Quintarelli, C.; Di Cecca, S.; Lagnese, G.; Punziano, A.; Guercio, M.; et al. B Cell Response Induced by SARS-CoV-2 Infection Is Boosted by the BNT162b2 Vaccine in Primary Antibody Deficiencies. Cells 2021, 11, 2915. [Google Scholar] [CrossRef]
- Matson, E.M.; Abyazi, M.L.; Bell, K.A.; Hayes, K.M.; Maglione, P.J. B Cell Dysregulation in Common Variable Immunodeficiency Interstitial Lung Disease. Front. Immunol. 2021, 11, 622114. [Google Scholar] [CrossRef]
- Hultberg, J.; Ernerudh, J.; Larsson, M.; Nilsdotter-Augustinsson, Å.; Nyström, S. Plasma protein profiling reflects TH1-driven immune dysregulation in common variable immunodeficiency. J. Allergy Clin. Immunol. 2020, 2, 417–428. [Google Scholar] [CrossRef] [Green Version]
- Cunningham-Rundles, C.; Radigan, L. Deficient IL-12 and dendritic cell function in common variable immune deficiency. Clin. Immunol. 2005, 115, 147–153. [Google Scholar] [CrossRef]
- Sauerwein, K.M.T.; Geier, C.B.; Stemberger, R.F.; Akyaman, H.; Illes, P.; Fischer, M.B.; Eibl, M.M.; Walter, J.E.; Wolf, H.M. Antigen-Specific CD4+ T-Cell Activation in Primary Antibody Deficiency After BNT162b2 mRNA COVID-19 Vaccination. Front. Immunol. 2022, 13, 827048. [Google Scholar] [CrossRef]
- Malphettes, M.; Gérard, L.; Carmagnat, M.; Mouillot, G.; Vince, N.; Boutboul, D.; Bérezné, A.; Nove-Josserand, R.; Lemoing, V.; Tetu, L.; et al. Late-onset combined immune deficiency: A subset of common variable immunodeficiency with severe T cell defect. Clin. Infect. Dis. 2009, 9, 1329–1338. [Google Scholar] [CrossRef]
- Bonilla, F.A.; Barlan, I.; Chapel, H.; Costa-Carvalho, B.T.; Cunningham-Rundles, C.; de la Morena, M.T.; Espinosa-Rosales, F.J.; Hammarström, L.; Nonoyama, S.; Quinti, I.; et al. International Consensus Document (ICON): Common Variable Immunodeficiency Disorders. J. Allergy Clin. Immunol. Pract. 2016, 4, 38–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conley, M.E.; Notarangelo, L.D.; Etzioni, A. Diagnostic Criteria for Primary Immunodeficiencies. Representing PAGID (Pan-American Group for Immunodeficiency) and ESID (European Society for Immunodeficiencies). Clin. Immunol. 1999, 3, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Seidel, M.G.; Kindle, G.; Gathmann, B.; Quinti, I.; Buckland, M.; van Montfrans, J. The European Society for Immunodeficiencies (ESID) Registry Working Definitions for the Clinical Diagnosis of Inborn Errors of Immunity. J. Allergy Clin. Immunol. Pract. 2019, 6, 1763–1770. [Google Scholar] [CrossRef] [PubMed]
- Ameratunga, R.; Ahn, Y.; Jordan, A.; Lehnert, K.; Brothers, S.; Woon, S.T. Keeping it in the family: The case for considering late-onset combined immunodeficiency a subset of common variable immunodeficiency disorders. Expert Rev. Clin. Immunol. 2018, 14, 549–556. [Google Scholar] [CrossRef]
- Pulvirenti, F.; Milito, C.; Cavaliere, F.M.; Mezzaroma, I.; Cinetto, F.; Quinti, I. IGA Antibody Induced by Immunization with Pneumococcal Polysaccharides Is a Prognostic Tool in Common Variable Immune Deficiencies. Front. Immunol. 2020, 11, 283. [Google Scholar] [CrossRef]
- Delmonte, O.M.; Bergerson, J.R.E.; Burbelo, P.D.; Durkee-Shock, J.R.; Dobbs, K.; Bosticardo, M.; Keller, M.D.; McDermott, D.H.; Rao, V.K.; Dimitrova, D.; et al. Antibody responses to the SARS-CoV-2 vaccine in individuals with various inborn errors of immunity. J. Allergy Clin. Immunol. 2021, 148, 1192–1197. [Google Scholar] [CrossRef]
- van Leeuwen, L.P.M.; GeurtsvanKessel, C.H.; Ellerbroek, P.M.; de Bree, G.J.; Potjewijd, J.; Rutgers, A.; Jolink, H.; van de Veerdonk, F.; van Gorp, E.C.M.; de Wilt, F.; et al. Immunogenicity of the mRNA-1273 COVID-19 vaccine in adult patients with inborn errors of immunity. J. Allergy Clin. Immunol. 2022, 149, 1949–1957. [Google Scholar] [CrossRef]
- Ameratunga, R.; Longhurst, H.; Steele, R.; Lehnert, K.; Leung, E.; Brooks, A.E.S.; Woon, S.T. Common Variable Immunodeficiency Disorders, T-Cell Responses to SARS-CoV-2 Vaccines, and the Risk of Chronic COVID-19. J. Allergy Clin. Immunol. Pract. 2021, 10, 3575–3583. [Google Scholar] [CrossRef]
- D’Amelio, R.; Asero, R.; Cassatella, M.A.; Laganà, B.; Lunardi, C.; Migliorini, P.; Nisini, R.; Parronchi, P.; Quinti, I.; Racanelli, V.; et al. Anti-COVID-19 Vaccination in Patients with Autoimmune-Autoinflammatory Disorders and Primary/Secondary Immunodeficiencies: The Position of the Task Force on Behalf of the Italian Immunological Societies. Biomedicines 2021, 9, 1163. [Google Scholar] [CrossRef] [PubMed]
- Hagin, D.; Freund, T.; Navon, M.; Halperin, T.; Adir, D.; Marom, R. Immunogenicity of Pfizer-BioNTech COVID-19 vaccine in patients with inborn errors of immunity. J. Allergy Clin. Immunol. 2021, 148, 739–749. [Google Scholar] [CrossRef]
- Pham, M.N.; Murugesan, K.; Banaei, N.; Pinsky, B.A.; Tang, M.; Hoyte, E.; Lewis, D.B.; Gernez, Y. Immunogenicity and tolerability of COVID-19 messenger RNA vaccines in primary immunodeficiency patients with functional B-cell defects. J. Allergy Clin. Immunol. 2022, 149, 907–911. [Google Scholar] [CrossRef] [PubMed]
- Pasparakis, M.; Alexopoulou, L.; Episkopou, V.; Kollias, G. Immune and inflammatory responses in TNF alpha-deficient mice: A critical requirement for TNF alpha in the formation of primary B cell follicles, follicular dendritic cell networks and germinal centers, and in the maturation of the humoral immune response. J. Exp. Med. 1996, 184, 1397–1411. [Google Scholar] [CrossRef] [PubMed]
- Opata, M.M.; Ye, Z.; Hollifield, M.; Garvy, B.A. B cell production of tumor necrosis factor in response to Pneumocystis murina infection in mice. Infect. Immun. 2013, 81, 4252–4260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, E.N.; Devlin, J.C.; Buus, T.B.; Koide, A.; Shwetar, J.; Cornelius, A. SARS-CoV-2 mRNA vaccine elicits a potent adaptive immune response in the absence of IFN-mediated inflammation observed in COVID-19. medRxiv 2021. [Google Scholar] [CrossRef]
- Maglione, P.J.; Cols, M.; Cunningham-Rundles, C. Dysregulation of Innate Lymphoid Cells in Common Variable Immunodeficiency. Curr. Allergy Asthma Rep. 2017, 11, 77. [Google Scholar] [CrossRef]
- Romberg, N.; Le Coz, C.; Glauzy, S.; Schickel, J.N.; Trofa, M.; Nolan, B.E. Patients with common variable immunodeficiency with autoimmune cytopenias exhibit hyperplastic yet inefficient germinal center responses. J. Allergy Clin. Immunol. 2019, 143, 258–265. [Google Scholar] [CrossRef] [Green Version]
- Del Pino-Molina, L.; López-Granados, E.; Lecrevisse, Q.; Torres Canizales, J.; Pérez-Andrés, M.; Blanco, E.; Wentink, M.; Bonroy, C.; Nechvatalova, J.; Milota, T.; et al. Dissection of the Pre-Germinal Center B-Cell Maturation Pathway in Common Variable Immunodeficiency Based on Standardized Flow Cytometric EuroFlow Tools. Front. Immunol. 2021, 11, 603972. [Google Scholar] [CrossRef]
- Sette, A.; Crotty, S. Adaptive immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef]
- Gupta, S.; Su, H.; Narsai, T.; Agrawal, S. SARS-CoV-2-Associated T-Cell Responses in the Presence of Humoral Immunodeficiency. Int. Arch. Allergy Immunol. 2021, 3, 195–209. [Google Scholar] [CrossRef]
- Friedmann, D.; Goldacker, S.; Peter, H.H.; Warnatz, K. Preserved Cellular Immunity Upon Influenza Vaccination in Most Patients with Common Variable Immunodeficiency. J. Allergy Clin. Immunol. Pract. 2020, 7, 2332–2340. [Google Scholar] [CrossRef]
- Pegu, A.; O’Connell, S.E.; Schmidt, S.D.; O’Dell, S.; Talana, C.A.; Lai, L. Durability of mRNA-1273 Vaccine-Induced Antibodies Against SARS-CoV-2 Variants. Science 2021, 373, 1372–1377. [Google Scholar] [CrossRef] [PubMed]
- Pulvirenti, F.; Milito, C.; Cinetto, F.; Fernandez Salinas, A.; Terreri, S.; Piano Mortari, E.; Auria, S.; Soccodato, V.; Miriam, L. Severe Acute Respiratory Syndrome Coronavirus 2 Monoclonal Antibody Combination Therapy in Patients With Coronavirus Disease 2019 and Primary Antibody Deficiency. J. Infect. Dis. 2022, 225, 820–824. [Google Scholar] [CrossRef] [PubMed]
- Milito, C.; Cinetto, F.; Palladino, A.; Garzi, G.; Punziano, A.; Lagnese, G.; Scarpa, R.; Rattazzi, M.; Pesce, A.M.; Pulvirenti, F.; et al. Mortality in Severe Antibody Deficiencies Patients during the First Two Years of the COVID-19 Pandemic: Vaccination and Monoclonal Antibodies Efficacy. Biomedicines 2022, 10, 1026. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| p Value | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| NR n = 17 | S1 IgG-R n = 18 | S1 IgG/IgA-R n = 12 | NR vs. IgG | NR vs. IgG/IgA-R | IgG vs. IgG/IgA | ||||
| Age (year), median (IQR) | 56 | (43–82) | 46 | (34–80) | 59 | (34–74) | 0.166 | 0.562 | 0.166 |
| Sex (female), n (%) | 10 | (59) | 11 | (61) | 6 | (50) | 0.485 | 1.000 | 0.421 |
| IgG (mg/dL), median (IQR) | 670 | (600–800) | 600 | (510–951) | 780 | (630–1130) | 0.023 | 0.028 | 0.023 |
| IgA (mg/dL), median (IQR) | 2 | (0–21) | 3 | (2–44) | 29 | (0–181) | 0.071 | 0.068 | 0.071 |
| IgM (mg/dL), median (IQR) | 3 | (0–142) | 13 | (2–75) | 66 | (1–117) | 0.026 | 0.906 | 0.026 |
| Lymphocytes (cells/mm3), median (IQR) | 54.5 | (20.5–74.1) | 50.9 | (27.9–73.8) | 49.8 | (1.53–70.7) | 0.954 | 0.471 | 0.47 |
| CD19+ (cells/mm3), median (IQR) | 19.8 | (1.63–83.66) | 47.1 | (11.9–161.5) | 86.86 | (22.5–208.9) | 0.0083 | <0.0001 | 0.134 |
| MBC (cells/mm3), median (IQR) | 1.52 | (0–7.5) | 6.27 | (2.4–52.2) | 37.93 | (6.45–92.18) | 0.0016 | <0.0001 | 0.0245 |
| IgM MBC (cells/mm3), median (IQR) | 1.49 | (0–7.5) | 6.08 | (2.3–44.7) | 26.1 | (3.89–73.56) | 0.0021 | <0.0001 | 0.0741 |
| Switched MBC (cells/mm3), median (IQR) | 0.01 | (0.00–0.27) | 0.15 | (0.1–5.8) | 5.36 | (1.93–16.54) | 0.0038 | <0.0001 | 0.0004 |
| CVID (All) n = 47 | NR n = 17 | S1 IgG R n = 19 | S1 IgG/IgA R n = 11 | HD n = 9 | |
|---|---|---|---|---|---|
| Lymphocytes (cells/mm3) | 1400 (1070–1930) | 1170 (525–1375) | 1870 (970–2470) | 1550 (1420–1770) | 2530 (2438–2775) |
| CD3+ (cells/mm3) | 1112 (758–1617) | 863 (451–1198) | 1280 (738–1930) | 1112 (966–1314) | 1960 (1767–2119) |
| CD3+CD4+ (cells/mm3) | 641 (470–876) | 490 (243–636) | 641 (470–998) | 876 (749–1127) | 1518 (1311–1664) |
| CD4+ naive (cells/mm3) | 107 (33–229) | 32 (14–194) | 107 (53–201) | 241 (87–395) | 791 (600–997) |
| CD4+ central memory (cells/mm3) | 146 (89–201) | 109 (55–161) | 140 (121–216) | 173 (64–291) | 376 (238–418) |
| CD4+ naïve/memory ratio | 0.76 (0.41–1.05 | 0.56 (0.305–1.055) | 0.78 (0.62–1.15) | 0.83(0.55–1.05) | 3.07 (2.855–5.07) |
| CD40L expression in CD45RO+CD4+ cells (%) | 23.4 (14–36.7) | 24.7 (12.8–33.95) | 20.1 (9.9–26.9) | 42.8 (23.4–51.8) | 58.8 (50.9–62.4) |
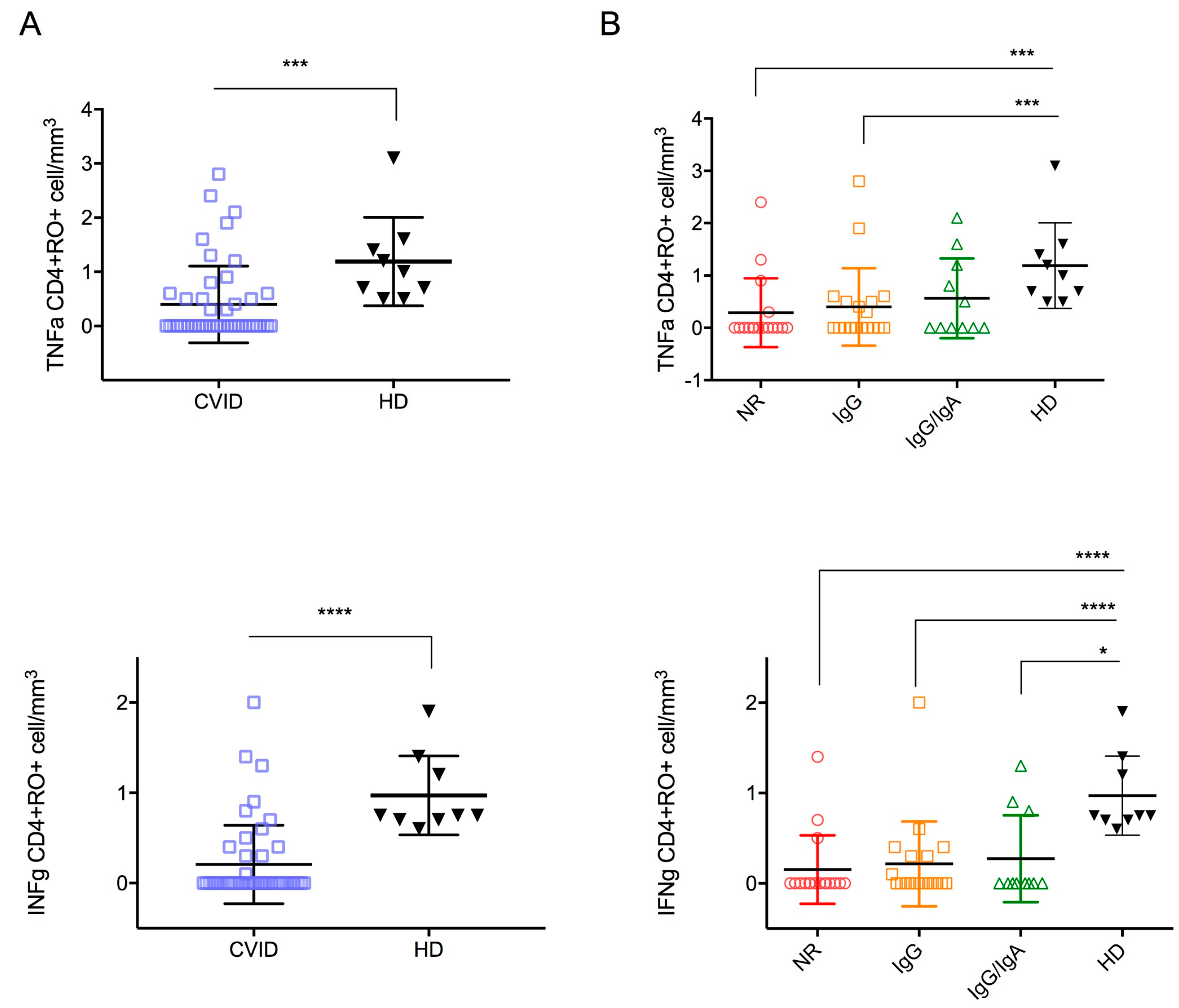
| TNFα CD45RO+CD4+ (cells/mm3) | 68.5 (37.2–98.4) | 66.8 (30.5–98) | 55.4 (31.3–77) | 75.5 (66.2–117.9) | 152.8 (91.7–233.5) |
| IFNγ CD45RO+CD4+ (cells/mm3) | 27 (8.5–51.3) | 29 (9.8–59.7) | 10.1 (5.6–35.3) | 34.3 (15.7–52.3) | 48 (23–113.6) |
| CD3+CD8+ (cells/mm3) |
542 (333–804) | 482 (210.5–693.5) | 736 (333–1106) | 461 (382–579) | 778 (730–1033) |
| CD8+ naive (cells/mm3) |
64 (31–117) | 28 (12–56.5) | 85 (34–117) | 106 (68–134) | 431 (313.5–496) |
| CD8+ central memory (cells/mm3) | 19 (10–33) | 17 (9.5–30.5) | 24 (13–38) | 19 (10–36) | 19 (11.5–27) |
| CD8+ naïve/memory ratio | 0.76 (0.41–1.05) | 0.11 (0.06–0.24) | 0.23 (0.11–0.3) | 0.47 (0.27–0.63) | 1.23 (0.71–2.61) |
| CD40L expression in CD45RO+CD8+ cells (%) | 2.6 (0.9–6.1) | 3.4 (1.35–7.5) | 3.0 (1.5–12.9) | 13.5 (4.1–18.3) | 7.2 (4.8–10.5) |
| TNFα CD45RO+CD8+ (cells/mm3) | 42 (24.7–79.8) | 59.7 (22.75–94.25) | 48.1 (34.5–82.6) | 24.8 (20.7–49.8) | 15.1 (11.05–26.25) |
| IFNγ CD45RO+CD8+ (cells/mm3) |
42.7 (22.8–97) |
63.4 (23.95–133.0) |
42.4 (33.4–91.1) | 22.8 (12.2–59.7) | 24.4 (8.85–33.7) |
| Peripheral blood Lymphocyte counts | Reduced |
| Circulating B cells | Reduced |
| Switched memory B cells | Reduced |
| Circulating CD4 T cells | Reduced |
| Circulating Naive CD4 and CD8 T cells | Reduced |
| CD40L expression in stimulated CD4+CD45+RO+ | Reduced |
| TNFα and IFNγ expressing CD8+ cells | Increased |
| IFNγ release by SARS-CoV-2-induced CD4+ T-cells | Reduced |
| Chronic Lung disease, Bronchiectasis | Increased |
| Autoimmune cytopenias | Increased |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pulvirenti, F.; Di Cecca, S.; Sinibaldi, M.; Piano Mortari, E.; Terreri, S.; Albano, C.; Guercio, M.; Sculco, E.; Milito, C.; Ferrari, S.; et al. T-Cell Defects Associated to Lack of Spike-Specific Antibodies after BNT162b2 Full Immunization Followed by a Booster Dose in Patients with Common Variable Immune Deficiencies. Cells 2022, 11, 1918. https://doi.org/10.3390/cells11121918
Pulvirenti F, Di Cecca S, Sinibaldi M, Piano Mortari E, Terreri S, Albano C, Guercio M, Sculco E, Milito C, Ferrari S, et al. T-Cell Defects Associated to Lack of Spike-Specific Antibodies after BNT162b2 Full Immunization Followed by a Booster Dose in Patients with Common Variable Immune Deficiencies. Cells. 2022; 11(12):1918. https://doi.org/10.3390/cells11121918
Chicago/Turabian StylePulvirenti, Federica, Stefano Di Cecca, Matilde Sinibaldi, Eva Piano Mortari, Sara Terreri, Christian Albano, Marika Guercio, Eleonora Sculco, Cinzia Milito, Simona Ferrari, and et al. 2022. "T-Cell Defects Associated to Lack of Spike-Specific Antibodies after BNT162b2 Full Immunization Followed by a Booster Dose in Patients with Common Variable Immune Deficiencies" Cells 11, no. 12: 1918. https://doi.org/10.3390/cells11121918