
Regulation of Dynamic Cell Adhesion by Integrin-Integrin Crosstalk


Abstract
:
1. Overview of Cell Adhesion
2. Cells Adhere through Integrins by Binding to Cellular and Extracellular Ligands after Activation
3. The Integrin β-Chains Have a Pivotal Role in Integrin Regulation, Whereas Integrin α-Chains Are Important for the Formation of the Ligand Binding Sites
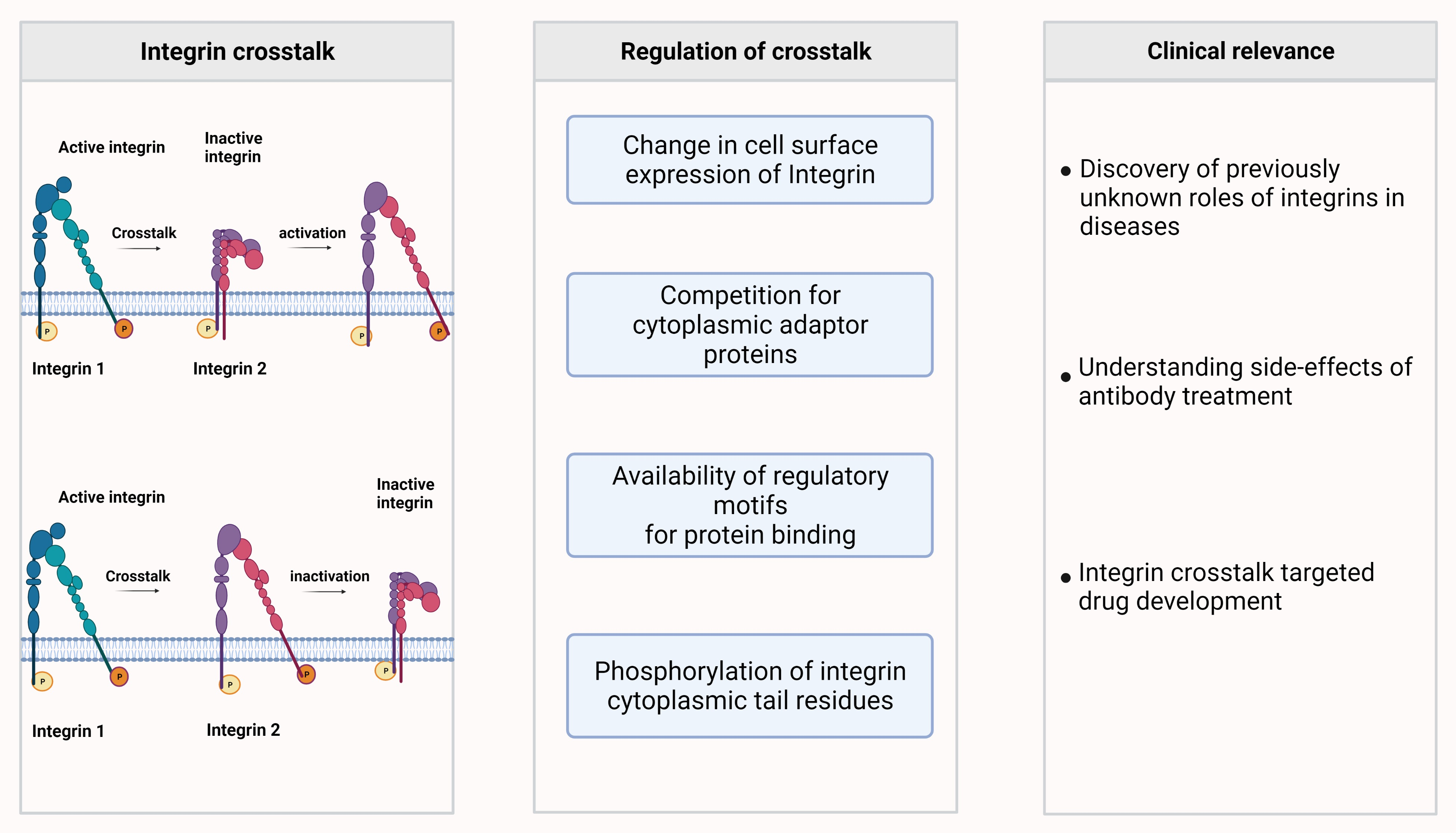
4. Integrin-Integrin Crosstalk Enables a Switch of Integrin Based Functions
5. Mechanisms of Integrin-Integrin Crosstalk
5.1. Changes in Integrin Expression or Integrin Chain Availability
5.2. Competition of Integrins for Cytoplasmic Adaptors
5.3. Important Regulatory Motifs in the Integrin β-Chains for Integrin Activity and Crosstalk
5.4. Integrin Phosphorylation Provides a Means for Rapid and Specific Trigger to Induce Integrin Activity and Crosstalk
6. Perspectives of the Role of Integrins in Crosstalk
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hynes, R.O. Integrins: Bidirectional, Allosteric Signaling Machines. Cell 2002, 110, 673–687. [Google Scholar] [CrossRef] [Green Version]
- Luo, B.-H.; Carman, C.V.; Springer, T.A. Structural Basis of Integrin Regulation and Signaling. Annu. Rev. Immunol. 2007, 25, 619–647. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Ye, F.; Ginsberg, M.H. Regulation of Integrin Activation. Annu. Rev. Cell Dev. Biol. 2011, 27, 321–345. [Google Scholar] [CrossRef]
- Gahmberg, C.G.; Grönholm, M.; Madhavan, S.; Jahan, F.; Mikkola, E.; Viazmina, L.; Koivunen, E. Regulation of cell adhesion: A collaborative effort of integrins, their ligands, cytoplasmic actors, and phosphorylation. Q. Rev. Biophys. 2019, 52, e10. [Google Scholar] [CrossRef]
- Gahmberg, C.G.; Grönholm, M. How integrin phosphorylations regulate cell adhesion and signaling. Trends Biochem. Sci. 2021, 47, 265–278. [Google Scholar] [CrossRef]
- Gonzalez, A.M.; Bhattacharya, R.; Dehart, G.W.; Jones, J.C. Transdominant regulation of integrin function: Mechanisms of crosstalk. Cell. Signal. 2010, 22, 578–583. [Google Scholar] [CrossRef] [Green Version]
- Samaržija, I.; Dekanić, A.; Humphries, J.D.; Paradžik, M.; Stojanović, N.; Humphries, M.J.; Ambriović-Ristov, A. Integrin Crosstalk Contributes to the Complexity of Signalling and Unpredictable Cancer Cell Fates. Cancers 2020, 12, 1910. [Google Scholar] [CrossRef]
- Eliceiri, B.P. Integrin and Growth Factor Receptor Crosstalk. Circ. Res. 2001, 89, 1104–1110. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, M.A.; Ginsberg, M.H. Networks and crosstalk: Integrin signalling spreads. Nat. Cell Biol. 2002, 4, E65–E68. [Google Scholar] [CrossRef]
- Kliche, J.; Kuss, H.; Ali, M.; Ivarsson, Y. Cytoplasmic short linear motifs in ACE2 and integrin β3 link SARS-CoV-2 host cell receptors to mediators of endocytosis and autophagy. Sci. Sign. 2021, 14, eabf1117. [Google Scholar] [CrossRef]
- Mészaros, B.; Samano-Sanchez, H.; Alvarado-Valverde, J.; Calyseva, J.; Martinez-Perez, E.; Alves, R.; Shields, D.C.; Kumar, M.; Rippmann, F.; Chemes, L.B.; et al. Short linear motif candidates in the cell entry system used by SARS-CoV-2 and their potential therapeutic implications. Sci. Sign. 2021, 14, eabd0334. [Google Scholar] [CrossRef]
- Pierschbacher, M.D.; Ruoslahti, E. Cell attachment activity of fibronectin can be duplicated by small synthetic fragments of the molecule. Nature 1984, 309, 30–33. [Google Scholar] [CrossRef]
- Park, E.; Myint, P.; Appiah, M.; Darkwah, S.; Caidengbate, S.; Ito, A.; Matsuo, E.; Kawamoto, E.; Gaowa, A.; Shimaoka, M. The Spike Glycoprotein of SARS-CoV-2 Binds to β1 Integrins Expressed on the Surface of Lung Epithelial Cells. Viruses 2021, 13, 645. [Google Scholar] [CrossRef]
- Simons, P.; Rinaldi, D.A.; Bondu, V.; Kell, A.M.; Bradfute, S.; Lidke, D.S.; Buranda, T. Integrin activation is an essential component of SARS-CoV-2 infection. Sci. Rep. 2021, 11, 20398. [Google Scholar] [CrossRef]
- Gahmberg, C.G. Leukocyte adhesion: CD11/CD18 integrins and intercellular adhesion molecules. Curr. Opin. Cell Biol. 1997, 9, 643–650. [Google Scholar] [CrossRef]
- Jakubowski, A.; Rosa, M.D.; Bixler, S.; Lobb, R.; Burkly, L.C. Vascular Cell Adhesion Molecule (VCAM)-Ig Fusion Protein Defines Distinct Affinity States of the Very Late Antigen-4 (VLA-4) Receptor. Cell Adhes. Commun. 1995, 3, 131–142. [Google Scholar] [CrossRef]
- Fan, Z.; McArdle, S.; Mark, G.; Mikulski, Z.; Gutierrez, E.; Engelhardt, B.; Deutsch, U.; Ginsberg, M.; Groisman, A.; Ley, K. Neutrophil recruitment limited by high-affinity bent β2 integrin binding ligand in cis. Nat. Commun. 2016, 7, 12658. [Google Scholar] [CrossRef] [Green Version]
- Fan, Z.; Kiosses, W.B.; Sun, H.; Orecchioni, M.; Ghosheh, Y.; Zajonc, D.M.; Arnaout, M.A.; Gutierrez, E.; Groisman, A.; Ginsberg, M.H.; et al. High-Affinity Bent β2-Integrin Molecules in Arresting Neutrophils Face Each Other through Binding to ICAMs In cis. Cell Rep. 2019, 26, 119–130.e5. [Google Scholar] [CrossRef] [Green Version]
- Dustin, M.; Springer, T.A. T-cell receptor cross-linking transiently stimulates adhesiveness through LFA-1. Nature 1989, 341, 619–624. [Google Scholar] [CrossRef]
- Van Kooyk, Y.; Kemenade, P.V.D.W.-V.; Weder, P.; Kuijpers, T.W.; Figdor, C.G. Enhancement of LFA-1-mediated cell adhesion by triggering through CD2 or CD3 on T lymphocytes. Nature 1989, 342, 811–813. [Google Scholar] [CrossRef] [Green Version]
- Patarroyo, M.; Beatty, P.G.; Fabre, J.W.; Gahmberg, C.G. Identification of a Cell Surface Protein Complex Mediating Phorbol Ester-Induced Adhesion (Binding) among Human Mononuclear Leukocytes. Scand. J. Immunol. 1985, 22, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Winograd-Katz, S.E.; Fässler, R.; Geiger, B.; Legate, K.R. The integrin adhesome: From genes and proteins to human disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Fagerholm, S.C.; Hilden, T.J.; Nurmi, S.M.; Gahmberg, C.G. Specific integrin α and β chain phosphorylations regulate LFA-1 activation through affinity-dependent and -independent mechanisms. J. Cell Biol. 2005, 171, 705–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, J.-P.; Mahalingham, B.; Alonso, J.L.; Borrelli, L.A.; Rui, X.; Anand, S.; Hyman, T.R.; Rysiok, T.; Muller-Pompalla, D.; Goodman, S.L.; et al. Crystal structure of the complete integrin αVβ3 ectodomain plus an α/β transmembrane fragment. J. Cell Biol. 2009, 186, 589–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abram, C.L.; Lowell, C.A. The Ins and Outs of Leukocyte Integrin Signaling. Annu. Rev. Immunol. 2009, 27, 339–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogg, N.; Patzak, I.; Willenbrock, F. The insider’s guide to leukocyte integrin signalling and function. Nat. Rev. Immunol. 2011, 11, 416–426. [Google Scholar] [CrossRef]
- Durrant, T.N.; van den Bosch, M.T.; Hers, I. Integrin αIIbβ3 outside-in signaling. Blood 2017, 130, 1607–1619. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Costell, M.; Fässler, R. Integrin activation by talin, kindlin and mechanical forces. Nat. Cell Biol. 2019, 21, 25–31. [Google Scholar] [CrossRef]
- Bialkowska, K.; Byzova, T.V.; Plow, E.F. Site-specific phosphorylation of kindlin-3 protein regulates its capacity to control cellular responses mediated by integrin αIIbβ3. J. Biol. Chem. 2015, 290, 6226–6242. [Google Scholar] [CrossRef] [Green Version]
- Bialkowska, K.; Sossey-Alaoui, K.; Pluskota, E.; Izem, L.; Qin, J.; Plow, E.F. Site-specific phosphorylation regulates the functions of kindlin-3 in a variety of cells. Life Sci. Alliance 2019, 3, e201900594. [Google Scholar] [CrossRef]
- Margraf, A.; Germena, G.; Drexler, H.C.A.; Rossaint, J.; Ludwing, N.; Prystai, B.; Mersamann, S.; Thomas, K.; Block, H.; Gottschilch, W.; et al. The integrin linked kinase is required for chemokine-triggered high affinity conformation of neutrophil beta2-integrin LFA-1. Blood 2020, 136, 2200–2205. [Google Scholar] [CrossRef] [PubMed]
- Bialkowska, K.; Qin, J.; Plow, E. Phosphorylation of Kindlins and the Control of Integrin Function. Cells 2021, 10, 825. [Google Scholar] [CrossRef] [PubMed]
- van Kooyk, Y.; Kemenade, E.V.D.W.-V.; Weder, P.; Huijbens, R.J.; Figdor, C. Lymphocyte function-associated antigen 1 dominates very late antigen 4 in binding of activated T cells to endothelium. J. Exp. Med. 1993, 177, 185–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blystone, S.D.; Graham, I.I.; Lindberg, F.P.; Brown, E.J. Integrin αvβ1 differentially regulates adhesive and phagocytic functions of the fibronectin receptor α5β1. J. Cell Biol. 1994, 127, 1129–1137. [Google Scholar] [CrossRef] [Green Version]
- Blystone, S.D.; Lindberg, F.P.; LaFlamme, S.E.; Brown, E.J. Integrin β3 cytoplasmic tail is necessary and sufficient for regulation of α5 phagocytosis by αvβ3 and integrin-associated protein. J. Cell Biol. 1995, 130, 745–754. [Google Scholar] [CrossRef]
- Díaz-González, F.; Forsyth, J.; Steiner, B.; Ginsberg, M.H. Trans-dominant inhibition of integrin function. Mol. Biol. Cell 1996, 7, 1939–1951. [Google Scholar] [CrossRef] [Green Version]
- Porter, J.C.; Hogg, N. Integrin cross talk: Activation of Lymphocyte function-associated antigen-1 on human T cells alters α4β1- and α5β1-mediated function. J. Cell Biol. 1997, 138, 1437–1447. [Google Scholar] [CrossRef]
- Chan, J.R.; Hyduk, S.J.; Cybulsky, M.H. α4β1 integrin/VCAM-1 interaction activates αLβ2 integrin-mediated adhesion to ICAM-1 in human T cells. J. Immunol. 2000, 164, 746–753. [Google Scholar] [CrossRef] [Green Version]
- Borza, C.M.; Pozzi, A.; Borza, D.-B.; Pedchenko, V.; Hellmark, T.; Hudson, B.G.; Zent, R. Integrin α3β1, a novel receptor for α3(IV) noncollagenous domain and a trans-dominant inhibitor for integrin αvβ3. J. Biol. Chem. 2006, 281, 20932–20939. [Google Scholar] [CrossRef] [Green Version]
- Bharadwaj, M.; Strohmeyer, N.; Colo, G.P.; Helenius, J.; Beerenwinkel, N.; Schiller, H.B.; Fässler, R.; Müller, D.J. αV-class integrins exert dual roles on α5β1 integrins to strengthen adhesion to fibronectin. Nat. Commun. 2017, 8, 14348. [Google Scholar] [CrossRef]
- Grönholm, M.; Jahan, F.; Bryushkova, E.A.; Madhavan, S.; Aglialoro, F.; Hinojosa, L.S.; Uotila, L.; Gahmberg, C.G. LFA-1 integrin antibodies inhibit leukocyte α4β1-mediated adhesion by intracellular signalling. Blood 2016, 128, 1270–1281. [Google Scholar] [CrossRef] [PubMed]
- Steiner, O.; Coisne, C.; Cecchelli, R.; Boscacci, R.; Deusch, U.; Engelhardt, B.; Lyck, R. Differential roles for endothelial ICAM-1, ICAM-2, and VCAM-1 in sheer-resistant T cell arrest, polarization, and directed crawling on blood-brain barrier endothelium. J. Immunol. 2010, 185, 4846–4855. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, G.A.; Anderson, N.; Hammer, D.A. The direction of migration of T-lymphocytes under flow depends upon which adhesion receptors are engaged. Integr. Biol. 2015, 7, 345–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valignat, M.-P.; Theodoly, O.; Gucciardi, A.; Hogg, N.; Lellouch, A.C. T Lymphocytes Orient against the Direction of Fluid Flow during LFA-1-Mediated Migration. Biophys. J. 2013, 104, 322–331. [Google Scholar] [CrossRef] [Green Version]
- Hornung, A.; Sbarrato, T.; Garcia-Seyda, N.; Aoun, L.; Luo, X.; Biarnes-Pelicot, M.; Theodoly, O.; Valignat, M.-P. A Bistable Mechanism Mediated by Integrins Controls Mechanotaxis of Leukocytes. Biophys. J. 2019, 118, 565–577. [Google Scholar] [CrossRef]
- Kim, S.H.J.; Hammer, D.A. Integrin crosstalk allows CD4+ T lymphocytes to continue migrating in the upstream direction after flow. Integr. Biol. 2019, 11, 384–393. [Google Scholar] [CrossRef]
- Kim, S.H.J.; Hammer, D.A. Integrin cross-talk modulates stiffness-independent motility of CD4+ T lymphocytes. Mol. Biol. Cell 2021, 32, 1749–1757. [Google Scholar] [CrossRef]
- Koistinen, P.; Heino, J. The selective regulation of αVβ1 integrin expression is based on the hierarchical formation of αV-containing heterodimers. J. Biol. Chem. 2002, 277, 24835–24841. [Google Scholar] [CrossRef] [Green Version]
- Parvani, J.G.; Galliher-Beckley, A.J.; Schiemann, B.J.; Schiemann, W.P. Targeted inactivation of β1 integrin induces β3 integrin switching, which drives breast cancer metastasis by TGF-β. Mol. Biol. Cell 2013, 24, 3449–3459. [Google Scholar] [CrossRef]
- Calderwood, D.A.; Tai, V.; Di Paolo, G.; De Camilli, P.; Ginsberg, M.H. Competition for Talin Results in Trans-dominant Inhibition of Integrin Activation. J. Biol. Chem. 2004, 279, 28889–28895. [Google Scholar] [CrossRef] [Green Version]
- Malinin, N.L.; Zhang, L.; Choi, J.; Ciocea, A.; Razorenova, O.; Ma, Y.-Q.; Podrez, E.A.; Tosi, M.; Lennon, D.P.; Caplan, A.I.; et al. A point mutation in KINDLIN3 ablates activation of three integrin subfamilies in humans. Nat. Med. 2009, 15, 313–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moser, M.; Bauer, M.; Schmid, S.; Ruppert, R.; Schmidt, S.; Sixt, M.; Wang, H.-V.; Sperandio, M.; Fässler, R. Kindlin-3 is required for β2 integrin–mediated leukocyte adhesion to endothelial cells. Nat. Med. 2009, 15, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Svensson, L.; Howarth, K.; McDowall, A.; Patzak, I.; Evans, R.; Ussar, S.; Moser, M.; Metin, A.; Fried, M.; Tomlinson, I.; et al. Leukocyte adhesion deficiency-III is caused by mutations in KINDLIN3 affecting integrin activation. Nat. Med. 2009, 15, 306–312. [Google Scholar] [CrossRef] [Green Version]
- Klapproth, S.; Moretti, F.A.; Zeiler, M.; Ruppert, R.; Breithaupt, U.; Mueller, S.; Haas, R.; Mann, M.; Sperandio, M.; Fässler, R.; et al. Minimal amounts of kindlin-3 suffice for basal platelet and leukocyte functions in mice. Blood 2015, 126, 2592–2600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stossel, T.P.; Condeelis, J.; Cooley, L.; Hartwig, J.H.; Noegel, A.; Schleicher, M.; Shapiro, S.S. Filamins as integrators of cell mechanics and signalling. Nat. Rev. Mol. Cell Biol. 2001, 2, 138–145. [Google Scholar] [CrossRef]
- Huang, J.; Zhou, Y.; Su, X.; Lyu, Y.; Tao, L.; Shi, X.; Liu, P.; Long, Z.; Ruan, Z.; Xiao, B.; et al. Roles of integrin β3 cytoplasmic tail in bidirectional signal transduction in a trans-dominant inhibition model. Front. Med. 2016, 10, 311–319. [Google Scholar] [CrossRef]
- Ruoslahti, E. Fibronectin and its receptors. Annu. Rev. Biochem. 1988, 57, 375–413. [Google Scholar] [CrossRef]
- van der Bijl, I.; Nawaz, K.; Kazlauskaite, U.; van Stalborch, A.-M.; Tol, S.; Orgaz, A.J.; Bout, I.V.D.; Reinhard, N.R.; Sonnenberg, A.; Margadant, C. Reciprocal integrin/integrin antagonism through kindlin-2 and Rho GTPases regulates cell cohesion and collective migration. Matrix Biol. 2020, 93, 60–78. [Google Scholar] [CrossRef]
- Wennerberg, K.; Fässler, R.; Wärmegård, B.; Johansson, S. Mutational analysis of the potential phosphorylation sites in the cytoplasmic domain of integrin β1A. J. Cell Sci. 1998, 111, 1117–1126. [Google Scholar] [CrossRef]
- Nilsson, S.; Kaniowska, D.; Brakebusch, C.; Fässler, R.; Johansson, S. Threonine 788 in integrin subunit β1 regulates integrin activation. Exp. Cell Res. 2006, 312, 844–853. [Google Scholar] [CrossRef]
- Gonzalez, A.M.; Claiborne, J.; Jones, J.C.R. Integrin Cross-talk in Endothelial Cells Is Regulated by Protein Kinase A and Protein Phosphatase 1. J. Biol. Chem. 2008, 283, 31849–31860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimm, T.M.; Dierdorf, N.I.; Betz, K.; Paone, C.; Hauck, C.R. PPM1F controls integrin activity via a conserved phospho-switch. J. Cell Biol. 2020, 219, e202001057. [Google Scholar] [CrossRef] [PubMed]
- Böttcher, R.T.; Strohmeyer, N.; Aretz, J.; Fässler, R. New insights into the phosphorylation of the threonine motif of the β1 integrin cytoplasmic domain. Life Sci. Alliance 2022, 5, e202101301. [Google Scholar] [CrossRef] [PubMed]
- Mulrooney, J.P.; Hong, T.; Grabel, L.B. Serine 785 phosphorylation of the β1 cytoplasmic domain modulates β1A-integrin-dependent functions. J. Cell Sci. 2001, 114, 2525–2533. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, H.; Lundin, L.; Thullberg, M.; Liu, Y.; Wang, Y.; Claesson-Welsh, L.; Strömblad, S. p21-activated Kinase 4 Phosphorylation of Integrin β5 Ser-759 and Ser-762 Regulates Cell Migration. J. Biol. Chem. 2010, 285, 23699–23710. [Google Scholar] [CrossRef] [Green Version]
- Fagerholm, S.C.; Varis, M.; Stefanidakis, M.; Hilden, T.J.; Gahmberg, C.G. α-Chain integrin activation to bind ICAMs and leukocyte extravasation in vivo. Blood 2006, 108, 3379–3386. [Google Scholar] [CrossRef] [Green Version]
- Uotila, L.M.; Aatonen, M.; Gahmberg, C.G. Integrin CD11c/CD18 α-Chain Phosphorylation Is Functionally Important. J. Biol. Chem. 2013, 288, 33494–33499. [Google Scholar] [CrossRef] [Green Version]
- Valmu, L.; Gahmberg, C.G. Treatment with okadaic acid reveals strong threonine phosphorylation of CD18 after activation of CD11/CD18 leukocyte integrin with phorbol esters or CD3 antibodies. J. Immunol. 1995, 155, 1175–1183. [Google Scholar]
- Fagerholm, S.; Morrice, N.; Gahmberg, C.; Cohen, P. Phosphorylation of the Cytoplasmic Domain of the Integrin CD18 Chain by Protein Kinase C Isoforms in Leukocytes. J. Biol. Chem. 2002, 277, 1728–1738. [Google Scholar] [CrossRef] [Green Version]
- Jahan, F.; Madhavan, S.; Rolova, T.; Viazmina, L.; Grönholm, M.; Gahmberg, C.G. Phosphorylation of the α-chain in the integrin LFA-1 enables β2-chain phosphorylation and α-actinin binding required for cell adhesion. J. Biol. Chem. 2018, 293, 12318–12330. [Google Scholar] [CrossRef] [Green Version]
- Grönholm, M.; Jahan, F.; Marchesan, S.; Karvonen, U.; Aatonen, M.; Narumanchi, S.; Gahmberg, C.G. TCR-Induced Activation of LFA-1 Involves Signaling through Tiam1. J. Immunol. 2011, 187, 3613–3619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takala, H.; Nurminen, E.; Nurmi, S.M.; Aatonen, M.; Strandin, T.; Takatalo, M.; Kiema, T.; Gahmberg, C.G.; Ylänne, J.; Fagerholm, S.C.; et al. β2 integrin phosphorylation on Thr758 acts as a molecular switch to regulate 14-3-3 and filamin binding. Blood 2008, 112, 1853–1862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uotila, L.M.; Jahan, F.; Hinojosa, L.S.; Melandri, E.; Grönholm, M.; Gahmberg, C.G. Specific Phosphorylations Transmit Signals from Leukocyte β2 to β1 Integrins and Regulate Adhesion. J. Biol. Chem. 2014, 289, 32230–32242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldfinger, L.E.; Tzima, E.; Stockton, R.; Kiosses, W.B.; Kinbara, K.; Tkachenko, E.; Gutierrez, E.; Groisman, A.; Nguyen, P.; Chien, S.; et al. Localized α4 Integrin Phosphorylation Directs Shear Stress–Induced Endothelial Cell Alignment. Circ. Res. 2008, 103, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Jung, O.; Beauvais, D.M.; Adams, K.M.; Rapraeger, A.C. VLA-4 phosphorylation during tumor and immune cell migration relies on its coupling to VEGFR2 and CXCR4 by syndecan-1. J. Cell Sci. 2019, 132, jcs.232645. [Google Scholar] [CrossRef]
- Hyduk, S.J.; Oh, J.; Xiao, H.; Chen, M.; Cybulsky, M.I. Paxillin selectively associates with constitutive and chemoattractant-induced high-affinity α4β1 integrins: Implications for integrin signaling. Blood 2004, 104, 2818–2824. [Google Scholar] [CrossRef]
- Rose, D.M.; Liu, S.; Woodside, D.G.; Han, J.; Schlaepfer, D.D.; Ginsberg, M.H. Paxillin Binding to the α4Integrin Subunit Stimulates LFA-1 (Integrin αLβ2)-Dependent T Cell Migration by Augmenting the Activation of Focal Adhesion Kinase/Proline-Rich Tyrosine Kinase-2. J. Immunol. 2003, 170, 5912–5918. [Google Scholar] [CrossRef] [Green Version]
- Roy, N.H.; Kim, S.H.J.; Buffone, A.; Blumenthal, D.; Huang, B.; Agarwal, S.; Schwartzberg, P.L.; Hammer, D.A.; Burkhardt, J.K. LFA-1 signals to promote actin polymerization and upstream migration in T cells. J. Cell Sci. 2020, 133, jcs248328. [Google Scholar] [CrossRef]
- Das, M.; Ithychanda, S.S.; Qin, J.; Plow, E.F. Mechanisms of talin-dependent integrin signaling and crosstalk. Biochim. Biophys. Acta (BBA)—Biomembr. 2013, 1838, 579–588. [Google Scholar] [CrossRef] [Green Version]
- Arias-Salgado, E.G.; Lizano, S.; Sarkar, S.; Brugge, J.S.; Ginsberg, M.H.; Shattil, S.J. Src kinase activation by direct interaction with the integrin beta cytoplasmic domain. Proc. Natl. Acad. Sci. USA 2003, 100, 13298–13302. [Google Scholar] [CrossRef] [Green Version]
- Bledzka, K.; Bialkowska, K.; Nie, H.; Qin, J.; Byzova, T.; Wu, C.; Plow, E.F.; Ma, Y.-Q. Tyrosine Phosphorylation of Integrin β3 Regulates Kindlin-2 Binding and Integrin Activation. J. Biol. Chem. 2010, 285, 30370–30374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blystone, S.D.; Slater, S.E.; Williams, M.P.; Crow, M.T.; Brown, E.J. A molecular mechanism of integrin crosstalk: AlphaVbeta3 suppression of calcium/calmodulin-dependent protein kinase II regulates alpha5beta1 function. J. Cell Biol. 1999, 145, 889–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirk, R.I.; Sanderson, M.R.; Lerea, K.M. Threonine phosphorylation of the β3 integrin cytoplasmic tail, at a site recognized by PDK1 and Akt/PKB in vitro, regulates Shc binding. J. Biol. Chem. 2000, 275, 30901–30906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K. The linkage between beta1 integrin and the actin cytoskeleton is differentially regulated by tyrosine and serine/threonine phosphorylation of beta1 integrin in normal and cancerous human breast cells. BMC Cell Biol. 2001, 2, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehberg, K.; Kliche, S.; Madencioglu, D.A.; Thiere, M.; Müller, B.; Meineke, B.M.; Freund, C.; Budinger, E.; Stork, O. The Serine/Threonine Kinase Ndr2 Controls Integrin Trafficking and Integrin-Dependent Neurite Growth. J. Neurosci. 2014, 34, 5342–5354. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.M.; Kwon, M.S.; Park, C.S.; Choi, K.R.; Chun, I.S.; Ahn, J.; Song, W.K. Modulation of thr phosphorylation of integrin beta1 during muscle differentiation. J. Biol. Chem. 2004, 279, 7082–7090. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Integrin Causing Crosstalk | Affected Integrin | Effect | Mechanism | References |
|---|---|---|---|---|
| α3β1, α6β1 | αVβ3 | INHIBITORY | PKA/PP1 β3 chain required β3 phospho-S752 | [61] |
| αVβ3 | α5β1 | INHIBITORY | CAMKII β3 chain required β3 phospho-S752 | [34,35,83] |
| αIIbβ3 | α2β1, α5β1 | INHIBITORY | β3 chain required β3 phospho-S752 | [36] |
| αLβ2 | α4β1 | INHIBITORY | β2 chain required β2 phospho-T758 | [37,41] |
| β3-integrin | α5β1 | INHIBITION | Competition for talin | [50] |
| αLβ2 | α5β1 | INHIBITORY | Not known | [37] |
| α4β1 | αLβ2 | ACTIVATING | Not known | [38] |
| α3β1 | αVβ3 | INHIBITORY | Not known | [39] |
| αV-integrins | α5β1 | ACTIVATING | Not known | [40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gahmberg, C.G.; Grönholm, M.; Madhavan, S. Regulation of Dynamic Cell Adhesion by Integrin-Integrin Crosstalk. Cells 2022, 11, 1685. https://doi.org/10.3390/cells11101685
Gahmberg CG, Grönholm M, Madhavan S. Regulation of Dynamic Cell Adhesion by Integrin-Integrin Crosstalk. Cells. 2022; 11(10):1685. https://doi.org/10.3390/cells11101685
Chicago/Turabian StyleGahmberg, Carl G., Mikaela Grönholm, and Sudarrshan Madhavan. 2022. "Regulation of Dynamic Cell Adhesion by Integrin-Integrin Crosstalk" Cells 11, no. 10: 1685. https://doi.org/10.3390/cells11101685