
Protocols for Generating Surfaces and Measuring 3D Organelle Morphology Using Amira

 , , , , , , add
Show full author list
, , , , , , add
Show full author list
Abstract
:
1. Introduction
2. Protocol
2.1. Installing and Preparing Amira
- Purchase and download Amira software from ThermoScientific: https://www.thermofisher.com/us/en/home/industrial/electron-microscopy/electron-microscopy-instruments-workflow-solutions/3d-visualization-analysis-software/amira-life-sciences-biomedical.html (accessed on 13 June 2021).
- Open Amira software.
- Select Project View > Open Data and import the image files to be analyzed from a specific folder.
- Select Read Complete Volume into Memory to ensure all images have been transferred.
- 5.
- Under Image Read Parameters, specify Voxel Size (nm) of the images according to the size of the files.
2.2. Three-Dimensional Segmentation in Amira
- Under the Project subsection, select an ortho slice to analyze.
- Under the Segmentation subsection, select the Brush tool and brush size appropriate for the analysis; brush sizes 2 and 3 are commonly used.
- 3.
- Activate the Wacom tablet device.
- 4.
- Calibrate the Wacom Pen
- 5.
- To calibrate, open the Wacom Tablet Properties application on the Wacom tablet.
- 6.
- Select the name of the stylus being used and then select Calibrate.
- 7.
- In each corner of the Wacom screen, a cross will appear. Using the stylus, select the middle of the cross.
- 8.
- Drag the Amira software window until it is visible onto the Wacom screen.
- 9.
- Using a stylus and the Wacom tablet, outline all instances of one type of structure (e.g., mitochondria or blood vessels) in the ortho slice.
- 10.
- When all structures of one type have been segmented, press F to segment the areas of the outlined structures.
- 11.
- Select a material color for the organelle type.
- 12.
- To change the shading of an area for better visibility, press D to change the settings between full color, outline, or gradient.
- 13.
- Under Segmentation > Selection, select the + icon to add the selection.
- 14.
- Repeat this process for each unique structure type on the ortho slice.
- 15.
- Repeat this process for each ortho slice ensuring that all structure types consistently use the same material color.
- 16.
- Using the Materials menu, colors can be adjusted, locked, or made invisible by clicking on the color panel, the 2D or 3D checkmarks, and/or the lock button.
- 17.
- Segmentation.
- 18.
- This segmentation protocol is based on the type of organelle; however, it can be based on individual organelles (i.e., segmentation of each mitochondrion separately).
- 19.
- To segment organelles individually, repeat Step 2.4 to create a new material for each individual organelle versus each type of structure.
2.3. Three-Dimensional Reconstruction of Segmented Structures in Amira
- Go to the Project Menu.
- Click on Selection Labels > Generate Surface.
- Select Apply.
- Rename the newly generated selection cell with the “. surf” suffix and click Surface View.
- The 3D structures should appear over an ortho slice.
- Disable the overlaid ortho image. Toggle off to hide the image by clicking on the orange rectangle labeled Ortho Slice and selecting the toggle button (blue square) under the Properties menu.
- 7.
- Make scale bars.
- 8.
- Right-click in the gray area under the Project subsection and select the option Scalebars.
- 9.
- Leave only the frame x-axis active.
- 10.
- Set the line width and font to something easy to read; 4 px and Arial 18 pt. font is recommended.
- 11.
- Set the position and size to the bottom right corner; the recommended scale is 500 nm.
2.4. Creating a Video/Animation in Amira
- 1.
- Right-click in the gray area under the Project subsection and select Camera Orbit.
- 2.
- Click on the newly created Camera Orbit box and select Movie Maker.
- 3.
- With Camera Orbit selected, select a parameter under Action, such as the Up Direction to create a simple animation.
- 4.
- Alternatively, right-click in the gray area under the Project subsection and select Camera Path.
- 5.
- Camera Path > Camera Path Editor will pop up a new viewer.
- 6.
- Camera Path > Add will add new keyframes in the viewer. The positions of the keyframes can be altered in the new viewer by moving the cuboidal objects. Add as many keyframes as desired and use the cursor to adjust the path between the keyframes.
- 7.
- Adjust the time between the keyframes in the Camera Path menu. This will allow more complex and specific animation movements.
- 8.
- With the Movie Maker label selected, choose the movie parameters, such as File Format (MPEG) and Size (1080P optional).
- 9.
- Click Apply at the bottom left.
- 10.
- Save video. See (Supplementary Table S5, Supplementary Figure S1)
2.5. Quantification
- Three-dimensional length and other length measurements can be set by selecting the ruler icon under the tool icon when viewing the Project view.
- Ensure a scale has been set in Amira.
- Select the Line measurement tool and click on the desired surface to measure.
- Click on the two points to be measured. For 3D length, this should be the Feret’s diameter.
- The length will be calculated automatically. Units may be changed by clicking on the number that appears on the line measurement.
- For other measurements, right-click in the gray area under the Project subsection and select Label Analysis.
- Click on the newly created Label Analysis box and ensure that the properties are visible at the bottom left of the screen.
- Under the Data and Intensity Image sections, ensure that the desired data are chosen for analysis, and ensure for all 3D measurements that 3D is selected under the Interpretation section.
- Under the Measures section, click on the “...” to choose quantifications.
- In the newly opened Measures window, click the ruler icon next to User Measures, and name the new group of measurements.
- Add all relevant measurements to the new measurement group.
- Area3d, Volume3d, and perimeter measurements, used for quantification in this study, are default measurements in Amira. Select them from the Native Measurements box and click the ruler icon in the middle to add to the measurement group.
- Sphericity is a custom measurement that can be added in the Measure Editor menu that opens when creating a new measurement group. In the output menu, type in the equation for sphericity according to Amira convention: (π**(1/3) *(6*Volume3d) **(2/3)/Area3d.
- Other quantifications are available as default measurements or can be added using an equation via the method presented in step 5.7.2.
- Once all measurements are added, click the Ok button to save this new group for present and future analysis.
- In the bottom left, select the newly created measurement group under the Measures section.
- Click the Apply button at the bottom left of the properties screen. Data can then be copied to create graphs in other software, such as GraphPad. See (Supplementary Table S6, Supplementary Figure S1).
3. Results
3.1. SBF-SEM Reveals Mitochondrial Changes in OPA1 smKO-Derived Skeletal Muscle
3.2. Three-Dimensional Reconstruction Allows for Identification of Mitochondria-ER Contact Sites (MERCs)
3.3. Three-Dimensional Reconstruction Shows Organelle Morphology Changes upon Knockdown of MFN-2
4. Discussion
5. Limitations
Online Methods

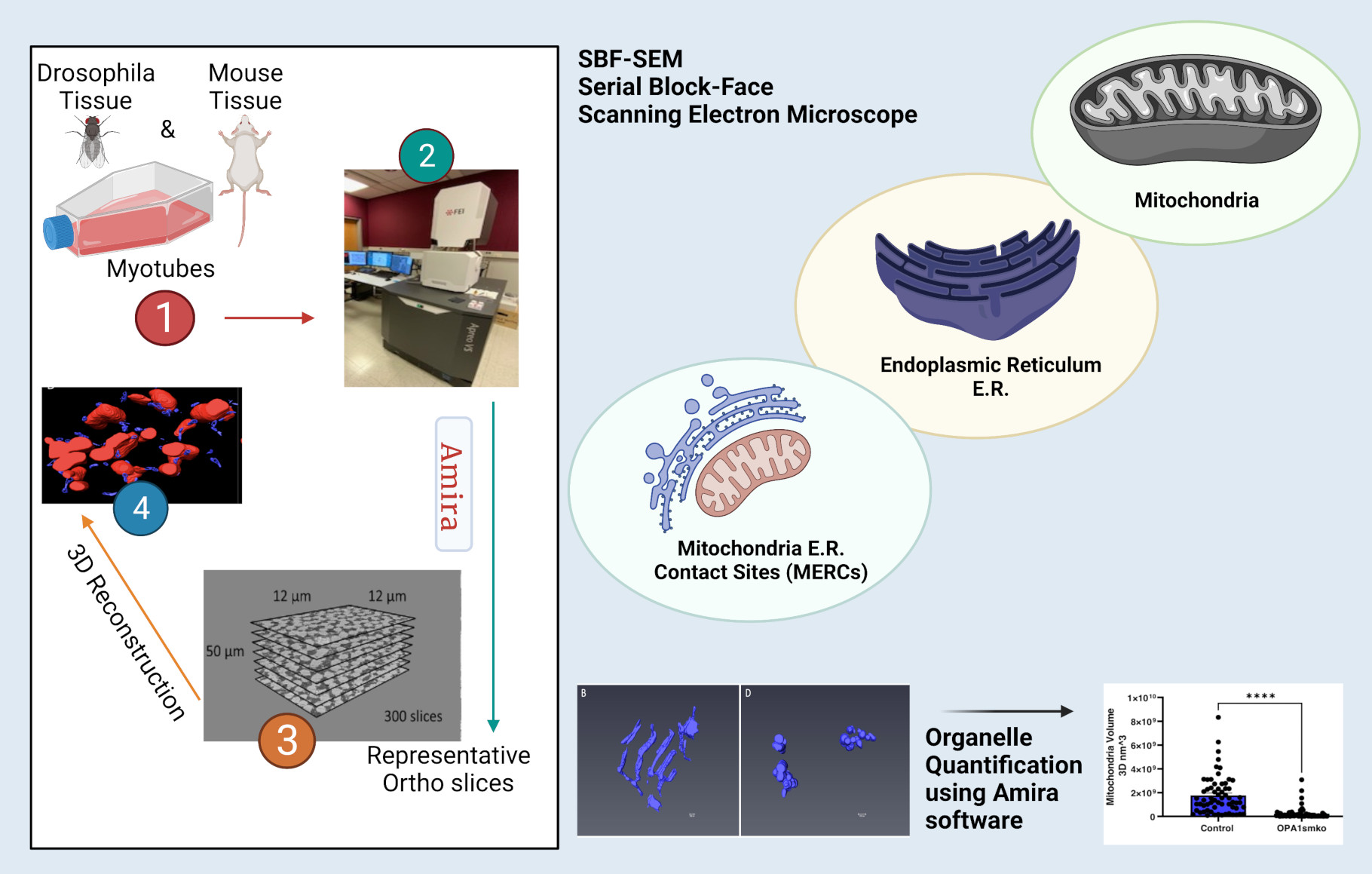{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent or Resource | Source | Identifier |
|---|---|---|
| Chemicals, peptides, and recombinant proteins | ||
| 2% glutaraldehyde in 0.1 M cacodylate buffer | N/A | N/A |
| 3% potassium ferrocyanide | Sigma | Cat# P3289 |
| 0.1% thiocarbohydrazide | Electron Microscopy Sciences | Cat# 21900 |
| 2% osmium tetroxide | Electron Microscopy Sciences | Cat# 19112 |
| 1% uranyl acetate | Electron Microscopy Sciences | Cat# 22400-2 |
| 0.6% lead aspartate solution | MP Biomedicals | Cat# 155180 |
| HPLC grade acetone | N/A | N/A |
| Epoxy 812 hard resin | Electron Microscopy Sciences | Cat# 14900 |
| Experimental Models: Organisms/strains | ||
| Mouse: C57B16 | Mayo Clinic | N/A |
| Drosophila: Mef2 Gal4 | VDRC (Vienna) Drosophila stock center and Bloomington Drosophila stock center. | BS# 27390 |
| Drosophila: W1118 | VDRC (Vienna) Drosophila stock center and Bloomington Drosophila stock center. | N/A |
| Drosophila: Opa-1-like | VDRC (Vienna) Drosophila stock center and Bloomington Drosophila stock center. | N/A |
| Software and Algorithms | ||
| Amira Software | Thermo Scientific/Amira [61] | RRID: SCR_007353 |
| Image J | Schneider et al. (2012) [62] | https://imagej.nih.gov/ij/ (assessed on 3 June 2021) |
| Ilastik | Berg et al. (2019) [63] | https://www.ilastik.org/publications.html (assessed on 3 June 2021) |
6. Resource Availability
6.1. Lead Contact
6.2. Materials Availability
6.3. Data and Code Availability
7. Experimental Model and Subject Details
7.1. Animal Care and Tissue Isolation
7.2. Fly Strains and Genetics
7.3. Serial Block Facing-Scanning Electron Microscopy (SBF-SEM) Processing of Drosophila and Mouse Muscle Fibers
7.4. Quantification
7.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Friederich, M.; Hansell, P.; Palm, F. Diabetes, Oxidative Stress, Nitric Oxide and Mitochondria Function. Curr. Diabetes Rev. 2009, 5, 120–144. [Google Scholar] [CrossRef]
- Bratic, A.; Larsson, N.-G. The Role of Mitochondria in Aging. J. Clin. Investig. 2013, 123, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Barja, G. The Mitochondrial Free Radical Theory of Aging. Prog. Mol. Biol. Transl. Sci. 2014, 127, 1–27. [Google Scholar] [PubMed]
- Olichon, A.; Baricault, L.; Gas, N.; Guillou, E.; Valette, A.; Belenguer, P.; Lenaers, G. Loss of OPA1 Perturbates the Mitochondrial Inner Membrane Structure and Integrity, Leading to Cytochrome c Release and Apoptosis. J. Biol. Chem. 2003, 278, 7743–7746. [Google Scholar] [CrossRef]
- Otera, H.; Miyata, N.; Kuge, O.; Mihara, K. Drp1-Dependent Mitochondrial Fission via MiD49/51 Is Essential for Apoptotic Cristae Remodeling. J. Cell Biol. 2016, 212, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Gunter, T.E.; Yule, D.I.; Gunter, K.K.; Eliseev, R.A.; Salter, J.D. Calcium and Mitochondria. FEBS Lett. 2004, 567, 96–102. [Google Scholar] [CrossRef]
- Dias, N.; Bailly, C. Drugs Targeting Mitochondrial Functions to Control Tumor Cell Growth. Biochem. Pharmacol. 2005, 70, 1–12. [Google Scholar] [CrossRef]
- Pinton, P.; Giorgi, C.; Siviero, R.; Zecchini, E.; Rizzuto, R. Calcium and Apoptosis: ER-Mitochondria Ca2+ Transfer in the Control of Apoptosis. Oncogene 2008, 27, 6407–6418. [Google Scholar] [CrossRef]
- Nicholls, D.G. Mitochondria and Calcium Signaling. Cell Calcium 2005, 38, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, A.; Wojtczak, L. Mitochondria as a Pharmacological Target. Pharmacol. Rev. 2002, 54, 101–127. [Google Scholar] [CrossRef] [PubMed]
- Glancy, B. Visualizing Mitochondrial Form and Function within the Cell. Trends Mol. Med. 2020, 26, 58–70. [Google Scholar] [CrossRef]
- Transmission Electron Microscopy vs.Scanning Electron Microscopy; ThermoScientific: Bangalore, India, 2019.
- Arborgh, B.; Bell, P.; Brunk, U.; Collins, V. The Osmotic Effect of Glutaraldehyde during Fixation. A Transmission Electron Microscopy, Scanning Electron Microscopy and Cytochemical Study. J. Ultrastruct. Res. 1976, 56, 339–350. [Google Scholar] [CrossRef]
- Courson, J.A.; Landry, P.T.; Spehlmann, E.; Lafontant, P.; Patel, N.; Rumbaut, R.; Burns, A. Serial Block-Face Scanning Electron Microscopy (SBF-SEM) of Biological Tissue Samples. J. Vis. Exp. Jove 2021. [Google Scholar] [CrossRef]
- Goggin, P.; Ho, E.M.; Gnaegi, H.; Searle, S.; Oreffo, R.O.; Schneider, P. Development of Protocols for the First Serial Block-Face Scanning Electron Microscopy (SBF SEM) Studies of Bone Tissue. Bone 2020, 131, 115107. [Google Scholar] [CrossRef] [PubMed]
- Baena, V.; Schalek, R.L.; Lichtman, J.W.; Terasaki, M. Serial-Section Electron Microscopy Using Automated Tape-Collecting Ultramicrotome (ATUM). Methods Cell Biol. 2019, 152, 41–67. [Google Scholar]
- Denk, W.; Horstmann, H.; Harris, K.M. Serial Block-Face Scanning Electron Microscopy to Reconstruct Three-Dimensional Tissue Nanostructure. PLoS Biol. 2004, 2, e329. [Google Scholar] [CrossRef]
- Lippens, S.; Kremer, A.; Borghgraef, P.; Guérin, C.J. Serial Block Face-Scanning Electron Microscopy for Volume Electron Microscopy. Methods Cell Biol. 2019, 152, 69–85. [Google Scholar] [PubMed]
- Hughes, L.; Hawes, C.; Monteith, S.; Vaughan, S. Serial Block Face Scanning Electron Microscopy—The Future of Cell Ultrastructure Imaging. Protoplasma 2014, 251, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, K.; Clark, H.R.; Chavan, V.; Benson, E.K.; Kidd, G.J.; Srivastava, S. Analysis of Brain Mitochondria Using Serial Block-Face Scanning Electron Microscopy. J. Vis. Exp. JoVE 2016, 54214. [Google Scholar] [CrossRef] [PubMed]
- Howard, V.; Reed, M. Unbiased Stereology: Three-Dimensional Measurement in Microscopy; Garland Science: New York, NY, USA, 2004; ISBN 1-135-33168-5. [Google Scholar]
- Lam, J.; Katti, P.; Biete, M.; Mungai, M.; AshShareef, S.; Neikirk, K.; Lopez, E.G.; Vue, Z.; Christensen, T.A.; Beasley, H.K. A Universal Approach to Analyzing Transmission Electron Microscopy with ImageJ. bioRxiv 2021, 10, 2177. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.E.; White, K.; Davey, T.; Philips, J.; Ogden, R.T.; Lawless, C.; Warren, C.; Hall, M.G.; Ng, Y.S.; Falkous, G. Quantitative 3D Mapping of the Human Skeletal Muscle Mitochondrial Network. Cell Rep. 2019, 26, 996–1009. [Google Scholar] [CrossRef] [PubMed]
- Cogliati, S.; Enriquez, J.A.; Scorrano, L. Mitochondrial Cristae: Where Beauty Meets Functionality. Trends Biochem. Sci. 2016, 41, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Kühlbrandt, W. Structure and Function of Mitochondrial Membrane Protein Complexes. BMC Biol. 2015, 13, 89. [Google Scholar] [CrossRef]
- Wolf, S.G.; Mutsafi, Y.; Dadosh, T.; Ilani, T.; Lansky, Z.; Horowitz, B.; Rubin, S.; Elbaum, M.; Fass, D. 3D Visualization of Mitochondrial Solid-Phase Calcium Stores in Whole Cells. Elife 2017, 6, e29929. [Google Scholar] [CrossRef] [PubMed]
- Frey, T.G.; Mannella, C.A. The Internal Structure of Mitochondria. Trends Biochem. Sci. 2000, 25, 319–324. [Google Scholar] [CrossRef]
- Jährling, N.; Becker, K.; Schönbauer, C.; Schnorrer, F.; Dodt, H.-U. Three-Dimensional Reconstruction and Segmentation of Intact Drosophila by Ultramicroscopy. Front. Syst. Neurosci. 2010, 4, 1. [Google Scholar] [CrossRef]
- Kopecky, B.; Duncan, J.; Elliott, K.; Fritzsch, B. Three-dimensional Reconstructions from Optical Sections of Thick Mouse Inner Ears Using Confocal Microscopy. J. Microsc. 2012, 248, 292–298. [Google Scholar] [CrossRef]
- Jährling, N.; Becker, K.; Dodt, H.-U. 3D-Reconstruction of Blood Vessels by Ultramicroscopy. Organogenesis 2009, 5, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Iwai, M.; Yokono, M.; Nakano, A. Visualizing Structural Dynamics of Thylakoid Membranes. Sci. Rep. 2014, 4, 3768. [Google Scholar] [CrossRef] [PubMed]
- Cocks, E.; Taggart, M.; Rind, F.; White, K. A Guide to Analysis and Reconstruction of Serial Block Face Scanning Electron Microscopy Data. J. Microsc. 2018, 270, 217–234. [Google Scholar] [CrossRef] [PubMed]
- Stalling, D.; Westerhoff, M.; Hege, H.-C. Amira: A Highly Interactive System for Visual Data Analysis. Vis. Handb. 2005, 38, 749–767. [Google Scholar]
- Suga, S.; Nakamura, K.; Humbel, B.M.; Kawai, H.; Hirabayashi, Y. An Interactive Deep Learning-Based Approach Reveals Mitochondrial Cristae Topologies. bioRxiv 2021. [Google Scholar] [CrossRef]
- Frezza, C.; Cipolat, S.; De Brito, O.M.; Micaroni, M.; Beznoussenko, G.V.; Rudka, T.; Bartoli, D.; Polishuck, R.S.; Danial, N.N.; De Strooper, B. OPA1 Controls Apoptotic Cristae Remodeling Independently from Mitochondrial Fusion. Cell 2006, 126, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Olichon, A.; Guillou, E.; Delettre, C.; Landes, T.; Arnauné-Pelloquin, L.; Emorine, L.J.; Mils, V.; Daloyau, M.; Hamel, C.; Amati-Bonneau, P. Mitochondrial Dynamics and Disease, OPA1. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2006, 1763, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, D.; Samakovlis, C.; Krasnow, M.A. Branchless Encodes a Drosophila FGF Homolog That Controls Tracheal Cell Migration and the Pattern of Branching. Cell 1996, 87, 1091–1101. [Google Scholar] [CrossRef]
- Sanz, A.; Stefanatos, R.K. The Mitochondrial Free Radical Theory of Aging: A Critical View. Curr. Aging Sci. 2008, 1, 10–21. [Google Scholar] [CrossRef]
- Rowland, A.A.; Voeltz, G.K. Endoplasmic Reticulum–Mitochondria Contacts: Function of the Junction. Nat. Rev. Mol. Cell Biol. 2012, 13, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Giacomello, M.; Pellegrini, L. The Coming of Age of the Mitochondria–ER Contact: A Matter of Thickness. Cell Death Differ. 2016, 23, 1417–1427. [Google Scholar] [CrossRef] [PubMed]
- Scorrano, L.; De Matteis, M.A.; Emr, S.; Giordano, F.; Hajnóczky, G.; Kornmann, B.; Lackner, L.L.; Levine, T.P.; Pellegrini, L.; Reinisch, K. Coming Together to Define Membrane Contact Sites. Nat. Commun. 2019, 10, 1287. [Google Scholar] [CrossRef]
- Kim, Y.; Lindberg, E.; Bleck, C.K.; Glancy, B. Endothelial Cell Nanotube Insertions into Cardiac and Skeletal Myocytes during Coordinated Tissue Development. Cardiovasc. Res. 2020, 116, 260–261. [Google Scholar] [CrossRef]
- De Brito, O.M.; Scorrano, L. Mitofusin 2 Tethers Endoplasmic Reticulum to Mitochondria. Nature 2008, 456, 605–610. [Google Scholar] [CrossRef]
- Filadi, R.; Greotti, E.; Pizzo, P. Highlighting the Endoplasmic Reticulum-Mitochondria Connection: Focus on Mitofusin 2. Pharmacol. Res. 2018, 128, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Merkwirth, C.; Langer, T. Mitofusin 2 Builds a Bridge between ER and Mitochondria. Cell 2008, 135, 1165–1167. [Google Scholar] [CrossRef] [PubMed]
- Ngoh, G.A.; Papanicolaou, K.N.; Walsh, K. Loss of Mitofusin 2 Promotes Endoplasmic Reticulum Stress. J. Biol. Chem. 2012, 287, 20321–20332. [Google Scholar] [CrossRef] [PubMed]
- Yasukawa, K.; Oshiumi, H.; Takeda, M.; Ishihara, N.; Yanagi, Y.; Seya, T.; Kawabata, S.; Koshiba, T. Mitofusin 2 Inhibits Mitochondrial Antiviral Signaling. Sci. Signal. 2009, 2, ra47. [Google Scholar] [CrossRef] [PubMed]
- Filadi, R.; Greotti, E.; Turacchio, G.; Luini, A.; Pozzan, T.; Pizzo, P. Mitofusin 2 Ablation Increases Endoplasmic Reticulum–Mitochondria Coupling. Proc. Natl. Acad. Sci. USA 2015, 112, E2174–E2181. [Google Scholar] [CrossRef]
- Naon, D.; Zaninello, M.; Giacomello, M.; Varanita, T.; Grespi, F.; Lakshminaranayan, S.; Serafini, A.; Semenzato, M.; Herkenne, S.; Hernández-Alvarez, M.I. Critical Reappraisal Confirms That Mitofusin 2 Is an Endoplasmic Reticulum–Mitochondria Tether. Proc. Natl. Acad. Sci. USA 2016, 113, 11249–11254. [Google Scholar] [CrossRef] [PubMed]
- Sebastián, D.; Hernández-Alvarez, M.I.; Segalés, J.; Sorianello, E.; Muñoz, J.P.; Sala, D.; Waget, A.; Liesa, M.; Paz, J.C.; Gopalacharyulu, P. Mitofusin 2 (Mfn2) Links Mitochondrial and Endoplasmic Reticulum Function with Insulin Signaling and Is Essential for Normal Glucose Homeostasis. Proc. Natl. Acad. Sci. USA 2012, 109, 5523–5528. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.G.; Williams, J.; Munoz-Pinedo, C.; Perkins, G.A.; Brown, J.M.; Ellisman, M.H.; Green, D.R.; Frey, T.G. Correlated Three-Dimensional Light and Electron Microscopy Reveals Transformation of Mitochondria during Apoptosis. Nat. Cell Biol. 2007, 9, 1057–1065. [Google Scholar] [CrossRef] [PubMed]
- Peddie, C.J.; Collinson, L.M. Exploring the Third Dimension: Volume Electron Microscopy Comes of Age. Micron 2014, 61, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Briggman, K.L.; Bock, D.D. Volume Electron Microscopy for Neuronal Circuit Reconstruction. Curr. Opin. Neurobiol. 2012, 22, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Lidke, D.S.; Lidke, K.A. Advances in High-Resolution Imaging–Techniques for Three-Dimensional Imaging of Cellular Structures. J. Cell Sci. 2012, 125, 2571–2580. [Google Scholar] [CrossRef] [PubMed]
- Renken, C.W. The Structure of Mitochondria. Ph.D. Thesis, University of California, San Diego, CA, USA, 2004. [Google Scholar]
- Willingham, T.B.; Kim, Y.; Lindberg, E.; Bleck, C.K.; Glancy, B. The Unified Myofibrillar Matrix for Force Generation in Muscle. Nat. Commun. 2020, 11, 3722. [Google Scholar] [CrossRef] [PubMed]
- Bleck, C.K.; Kim, Y.; Willingham, T.B.; Glancy, B. Subcellular Connectomic Analyses of Energy Networks in Striated Muscle. Nat. Commun. 2018, 9, 5111. [Google Scholar] [CrossRef] [PubMed]
- Faitg, J.; Lacefield, C.; Davey, T.; White, K.; Laws, R.; Kosmidis, S.; Reeve, A.K.; Kandel, E.R.; Vincent, A.E.; Picard, M. 3D Neuronal Mitochondrial Morphology in Axons, Dendrites, and Somata of the Aging Mouse Hippocampus. bioRxiv 2021. [Google Scholar] [CrossRef] [PubMed]
- Soufan, A.T.; Ruijter, J.M.; van den Hoff, M.J.; de Boer, P.A.; Hagoort, J.; Moorman, A.F. Three-Dimensional Reconstruction of Gene Expression Patterns during Cardiac Development. Physiol. Genom. 2003, 13, 187–195. [Google Scholar] [CrossRef] [PubMed]
- De Boer, B.A.; Soufan, A.T.; Hagoort, J.; Mohun, T.J.; van den Hoff, M.J.; Hasman, A.; Voorbraak, F.P.; Moorman, A.F.; Ruijter, J.M. The Interactive Presentation of 3D Information Obtained from Reconstructed Datasets and 3D Placement of Single Histological Sections with the 3D Portable Document Format. Development 2011, 138, 159–167. [Google Scholar] [CrossRef]
- Lanika Solutions. In Amira Software For Life & Biomedical Sciences; ThermoScientific: Bangalore, India, 2019.
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Berg, S.; Kutra, D.; Kroeger, T.; Straehle, C.N.; Kausler, B.X.; Haubold, C.; Schiegg, M.; Ales, J.; Beier, T.; Rudy, M. Ilastik: Interactive Machine Learning for (Bio) Image Analysis. Nat. Methods 2019, 16, 1226–1232. [Google Scholar] [CrossRef]
- Mustafi, D.; Kikano, S.; Palczewski, K. Serial Block Face-Scanning Electron Microscopy: A Method to Study Retinal Degenerative Phenotypes. Curr. Protoc. Mouse Biol. 2014, 4, 197–204. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garza-Lopez, E.; Vue, Z.; Katti, P.; Neikirk, K.; Biete, M.; Lam, J.; Beasley, H.K.; Marshall, A.G.; Rodman, T.A.; Christensen, T.A.; et al. Protocols for Generating Surfaces and Measuring 3D Organelle Morphology Using Amira. Cells 2022, 11, 65. https://doi.org/10.3390/cells11010065
Garza-Lopez E, Vue Z, Katti P, Neikirk K, Biete M, Lam J, Beasley HK, Marshall AG, Rodman TA, Christensen TA, et al. Protocols for Generating Surfaces and Measuring 3D Organelle Morphology Using Amira. Cells. 2022; 11(1):65. https://doi.org/10.3390/cells11010065
Chicago/Turabian StyleGarza-Lopez, Edgar, Zer Vue, Prasanna Katti, Kit Neikirk, Michelle Biete, Jacob Lam, Heather K. Beasley, Andrea G. Marshall, Taylor A. Rodman, Trace A. Christensen, and et al. 2022. "Protocols for Generating Surfaces and Measuring 3D Organelle Morphology Using Amira" Cells 11, no. 1: 65. https://doi.org/10.3390/cells11010065