
KANPHOS: A Database of Kinase-Associated Neural Protein Phosphorylation in the Brain

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. System Configuration and Application Software
2.2. Data Collection, Management and Analysis Tool
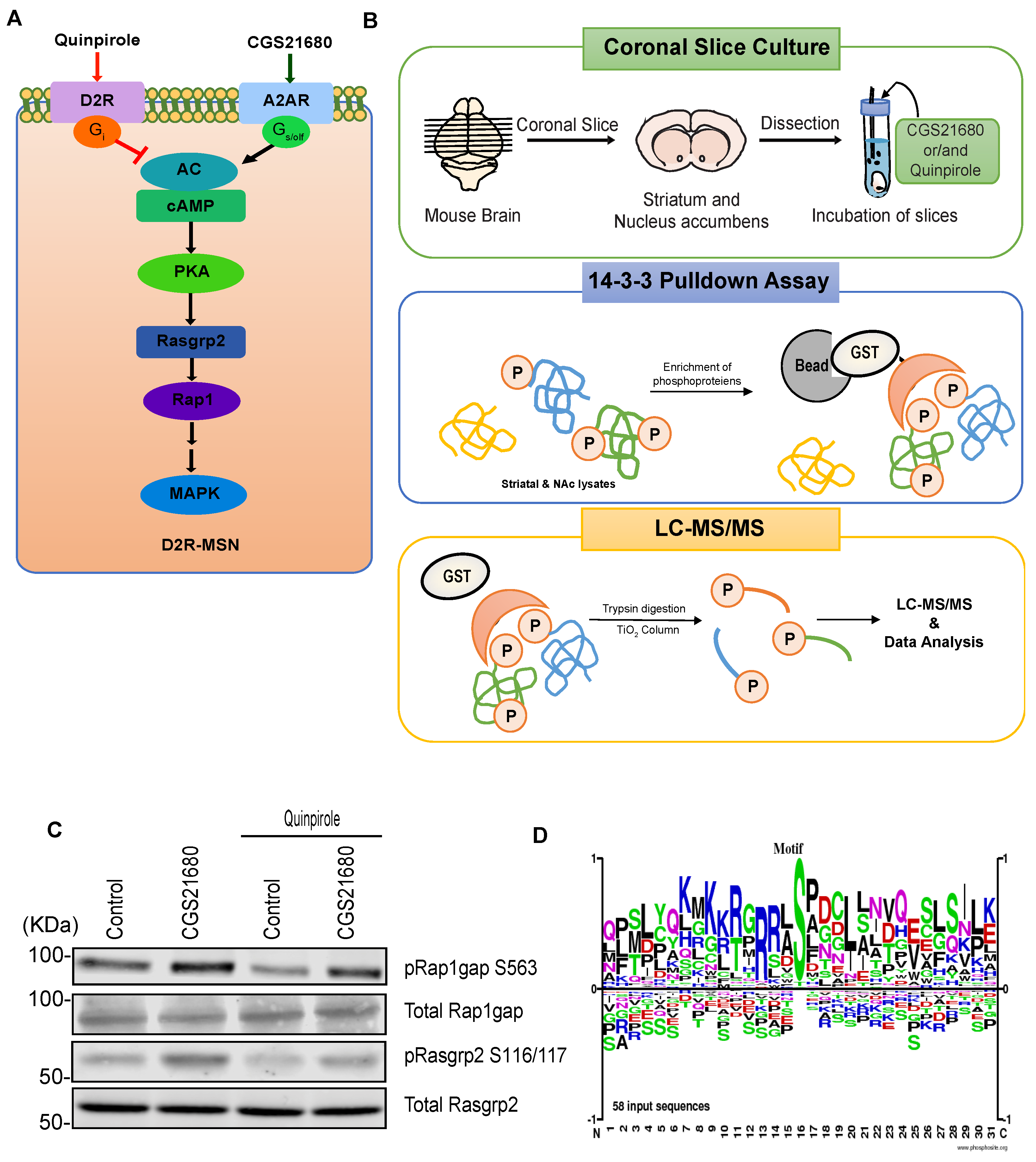
2.3. Preparation and Incubation of Coronal Slices
2.4. Mass Spectrometry
2.5. SDS-PAGE and Immunoblotting
3. Results
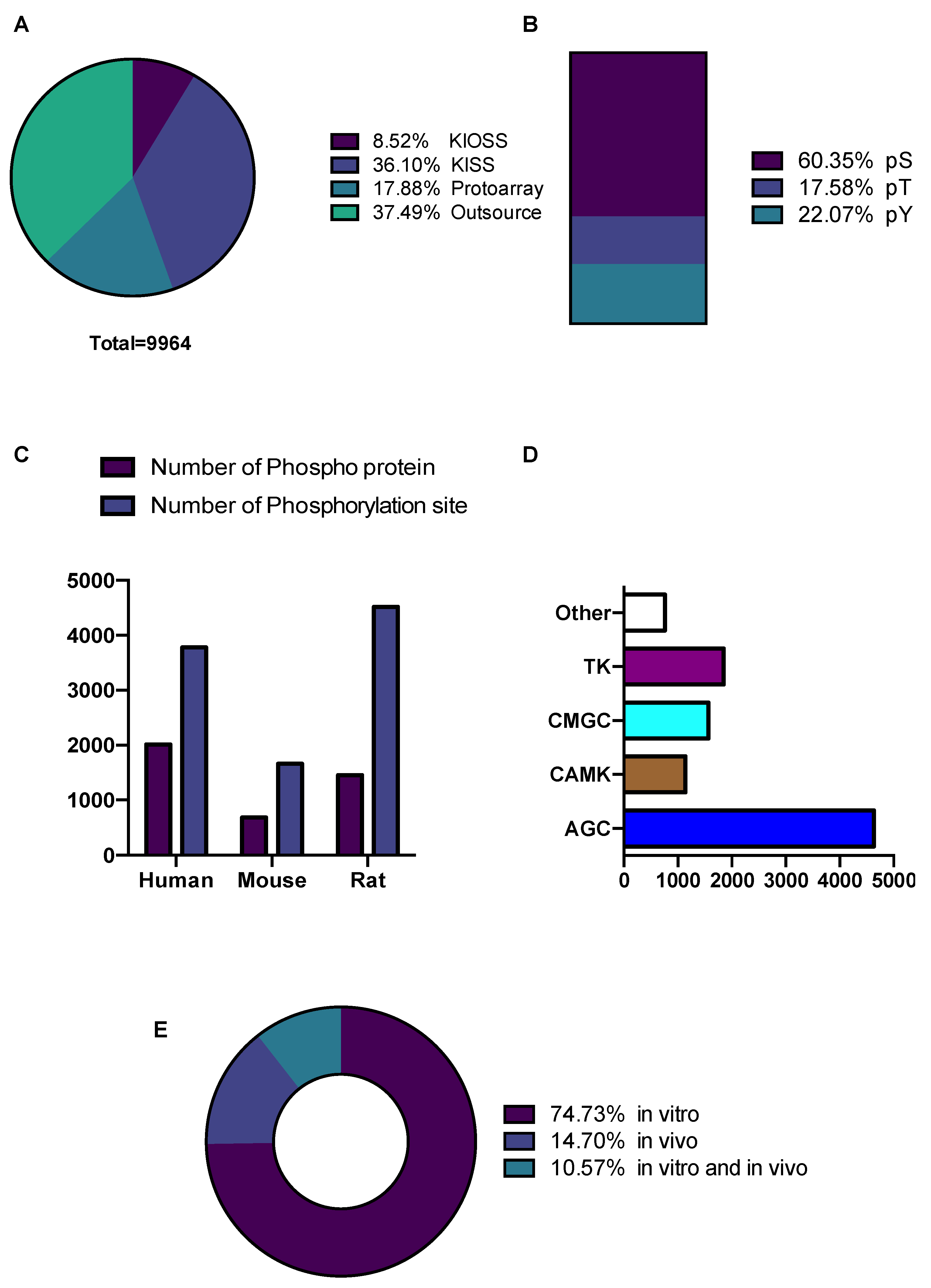
3.1. Construction and Content of the KANPHOS Database
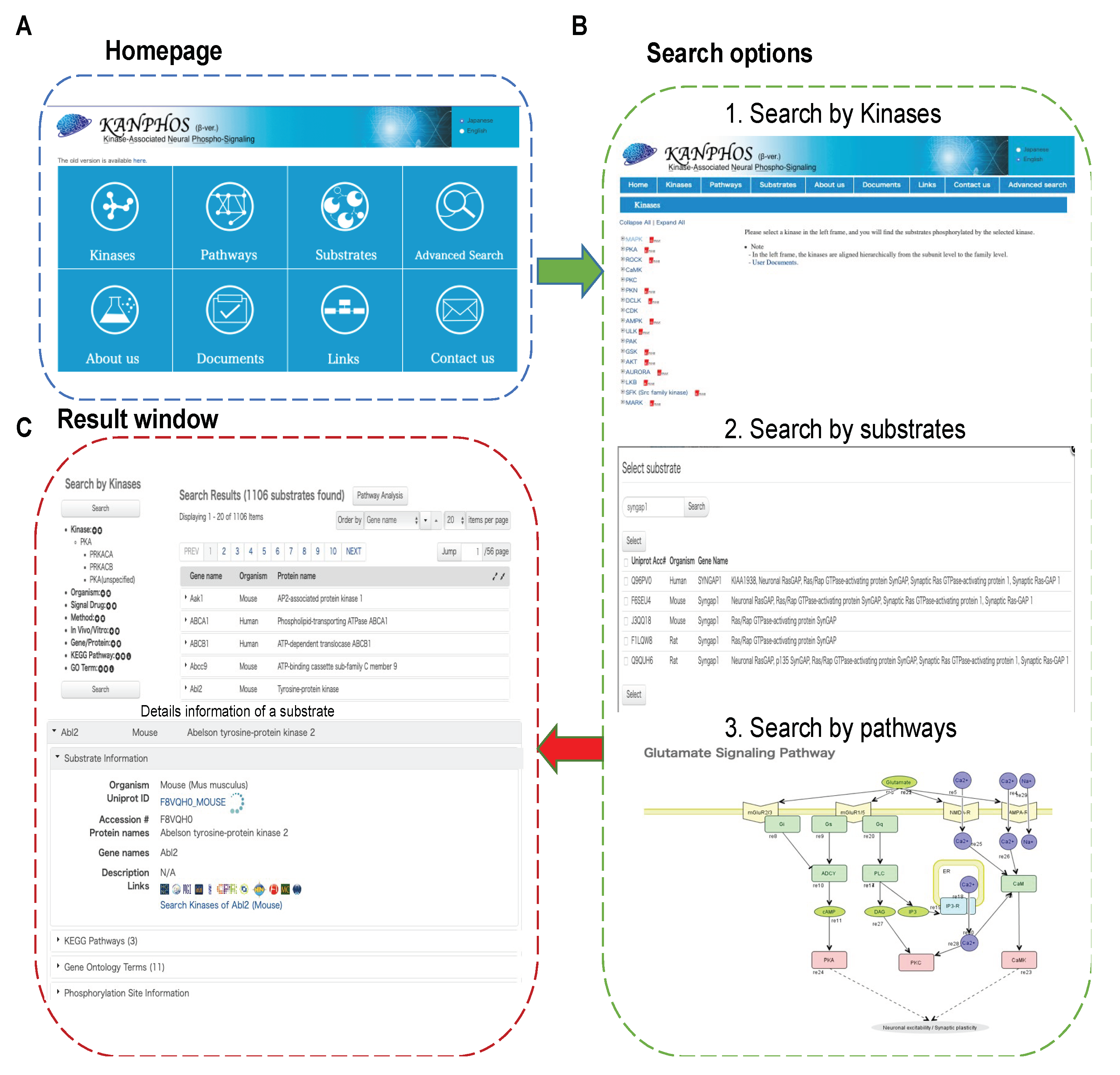
3.2. Data Accessibility and KANPHOS Workflow
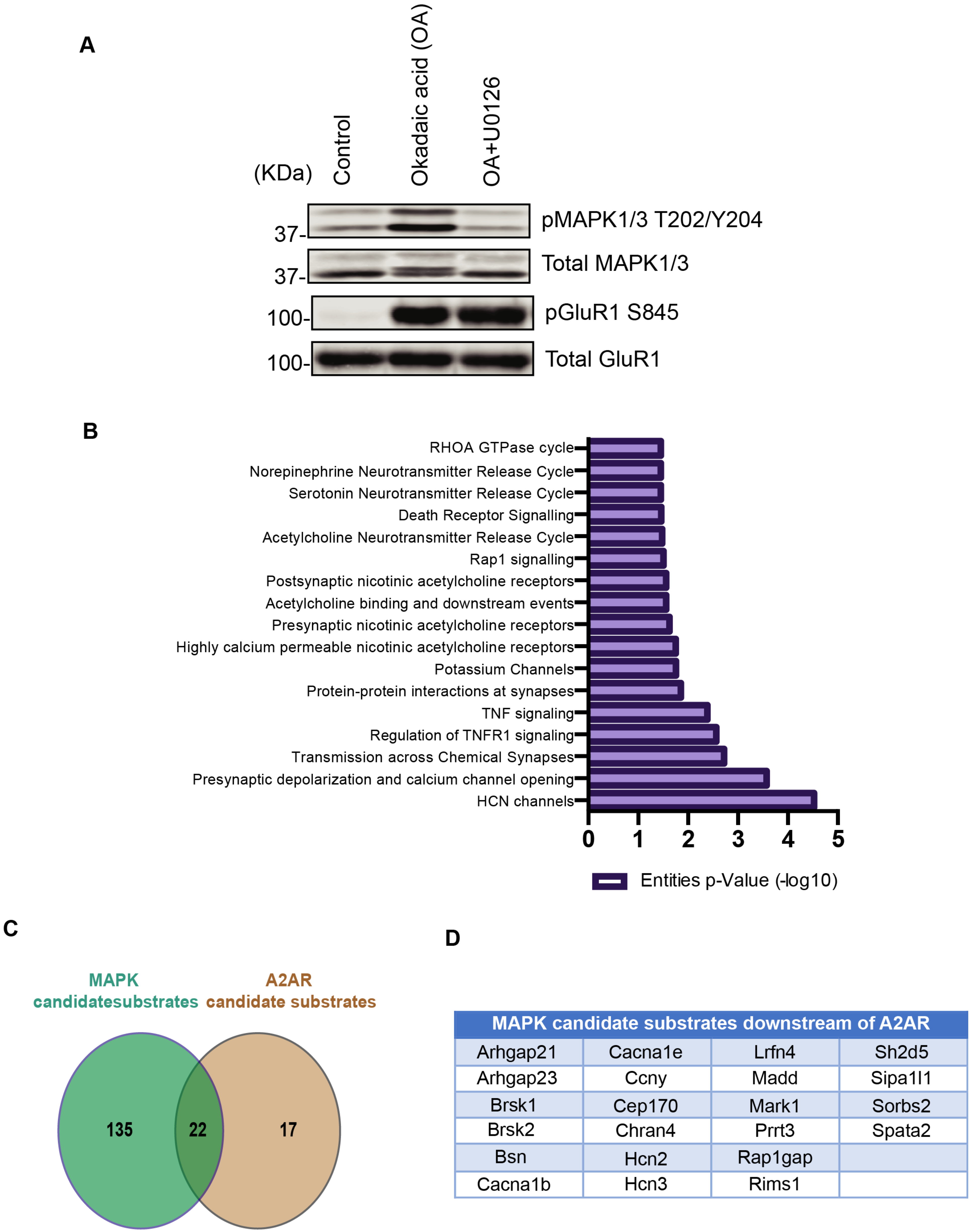
3.3. Case Study: Identification of Adenosine-A2A Receptor Signaling and MAPK-Mediated Signaling Molecules Using KANPHOS
3.4. Pathway Analysis Using KANPHOS Revealed That HCN and Calcium Channel Proteins Are Involved in the MEK-MAPK Pathway Downstream of A2AR
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cohen, P. The origins of protein phosphorylation. Nat. Cell. Biol. 2002, 4, E127–E130. [Google Scholar] [CrossRef] [PubMed]
- Pawson, T.; Scott, J.D. Protein phosphorylation in signaling—50 years and counting. Trends Biochem. Sci. 2005, 30, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.O.; Yu, L.; Coba, M.P.; Husi, H.; Campuzano, I.; Blackstock, W.P.; Choudhary, J.S.; Grant, S.G. Proteomic analysis of in vivo phosphorylated synaptic proteins. J. Biol. Chem. 2005, 280, 5972–5982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trinidad, J.C.; Specht, C.G.; Thalhammer, A.; Schoepfer, R.; Burlingame, A.L. Comprehensive identification of phosphorylation sites in postsynaptic density preparations. Mol. Cell. Proteom. 2006, 5, 914–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, P. The role of protein phosphorylation in human health and disease. The Sir Hans Krebs Medal Lecture. Eur. J. Biochem. 2001, 268, 5001–5010. [Google Scholar] [CrossRef]
- Manning, G.; Whyte, D.B.; Martinez, R.; Hunter, T.; Sudarsanam, S. The protein kinase complement of the human genome. Science 2002, 298, 1912–1934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nestler, E.J.; Greengard, P. Protein phosphorylation in the brain. Nature 1983, 305, 583–588. [Google Scholar] [CrossRef]
- Ghanizadeh, A.; Sahraeizadeh, A.; Berk, M. A head-to-head comparison of aripiprazole and risperidone for safety and treating autistic disorders, a randomized double blind clinical trial. Child Psychiatry Hum. Dev. 2014, 45, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Posey, D.J.; Stigler, K.A.; Erickson, C.A.; McDougle, C.J. Antipsychotics in the treatment of autism. J. Clin. Investig. 2008, 118, 6–14. [Google Scholar] [CrossRef]
- Woods, S.W. Chlorpromazine equivalent doses for the newer atypical antipsychotics. J. Clin. Psychiatry 2003, 64, 663–667. [Google Scholar] [CrossRef]
- Leucht, S.; Cipriani, A.; Spineli, L.; Mavridis, D.; Orey, D.; Richter, F.; Samara, M.; Barbui, C.; Engel, R.R.; Geddes, J.R.; et al. Comparative efficacy and tolerability of 15 antipsychotic drugs in schizophrenia: A multiple-treatments meta-analysis. Lancet 2013, 382, 951–962. [Google Scholar] [CrossRef]
- Meltzer, H.Y. The role of serotonin in antipsychotic drug action. Neuropsychopharmacology 1999, 21 (Suppl. S2), 106S–115S. [Google Scholar] [CrossRef]
- Millan, M.J. Improving the treatment of schizophrenia: Focus on serotonin (5-HT)(1A) receptors. J. Pharmacol. Exp. Ther. 2000, 295, 853–861. [Google Scholar]
- Mailman, R.B.; Murthy, V. Third generation antipsychotic drugs: Partial agonism or receptor functional selectivity? Curr. Pharm. Des. 2010, 16, 488–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- UniProt, C. UniProt: A hub for protein information. Nucleic Acids Res. 2015, 43, D204–D212. [Google Scholar]
- Diella, F.; Gould, C.M.; Chica, C.; Via, A.; Gibson, T.J. Phospho.ELM: A database of phosphorylation sites—Update 2008. Nucleic Acids Res. 2008, 36, D240–D244. [Google Scholar] [CrossRef]
- Hornbeck, P.V.; Kornhauser, J.M.; Tkachev, S.; Zhang, B.; Skrzypek, E.; Murray, B.; Latham, V.; Sullivan, M. PhosphoSitePlus: A comprehensive resource for investigating the structure and function of experimentally determined post-translational modifications in man and mouse. Nucleic Acids Res. 2012, 40, D261–D270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gnad, F.; Gunawardena, J.; Mann, M. PHOSIDA 2011: The posttranslational modification database. Nucleic Acids Res. 2011, 39, D253–D260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Xing, X.; Ding, G.; Li, Q.; Wang, C.; Xie, L.; Zeng, R.; Li, Y. SysPTM: A systematic resource for proteomic research on post-translational modifications. Mol. Cell. Proteom. 2009, 8, 1839–1849. [Google Scholar] [CrossRef] [Green Version]
- Moriya, Y.; Kawano, S.; Okuda, S.; Watanabe, Y.; Matsumoto, M.; Takami, T.; Kobayashi, D.; Yamanouchi, Y.; Araki, N.; Yoshizawa, A.C.; et al. The jPOST environment: An integrated proteomics data repository and database. Nucleic Acids Res. 2019, 47, D1218–D1224. [Google Scholar] [CrossRef] [PubMed]
- Peri, S.; Navarro, J.D.; Kristiansen, T.Z.; Amanchy, R.; Surendranath, V.; Muthusamy, B.; Gandhi, T.K.; Chandrika, K.N.; Deshpande, N.; Suresh, S.; et al. Human protein reference database as a discovery resource for proteomics. Nucleic Acids Res. 2004, 32, D497–D501. [Google Scholar] [CrossRef] [PubMed]
- Su, M.G.; Lee, T.Y. Incorporating substrate sequence motifs and spatial amino acid composition to identify kinase-specific phosphorylation sites on protein three-dimensional structures. BMC Bioinform. 2013, 14 (Suppl. S16), S2. [Google Scholar] [PubMed] [Green Version]
- Linding, R.; Jensen, L.J.; Ostheimer, G.J.; van Vugt, M.A.; Jorgensen, C.; Miron, I.M.; Diella, F.; Colwill, K.; Taylor, L.; Elder, K.; et al. Systematic discovery of in vivo phosphorylation networks. Cell 2007, 129, 1415–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.D.; Lee, T.Y.; Tzeng, S.W.; Wu, L.C.; Horng, J.T.; Tsou, A.P.; Huang, K.T. Incorporating hidden Markov models for identifying protein kinase-specific phosphorylation sites. J. Comput. Chem. 2005, 26, 1032–1041. [Google Scholar] [CrossRef] [PubMed]
- Amano, M.; Hamaguchi, T.; Shohag, M.H.; Kozawa, K.; Kato, K.; Zhang, X.; Yura, Y.; Matsuura, Y.; Kataoka, C.; Nishioka, T.; et al. Kinase-interacting substrate screening is a novel method to identify kinase substrates. J. Cell. Biol. 2015, 209, 895–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagai, T.; Nakamuta, S.; Kuroda, K.; Nakauchi, S.; Nishioka, T.; Takano, T.; Zhang, X.; Tsuboi, D.; Funahashi, Y.; Nakano, T.; et al. Phosphoproteomics of the Dopamine Pathway Enables Discovery of Rap1 Activation as a Reward Signal In Vivo. Neuron 2016, 89, 550–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishioka, T.; Nakayama, M.; Amano, M.; Kaibuchi, K. Proteomic screening for Rho-kinase substrates by combining kinase and phosphatase inhibitors with 14-3-3zeta affinity chromatography. Cell Struct. Funct. 2012, 37, 39–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funahashi, Y.; Watanabe, T.; Kaibuchi, K. Advances in defining signaling networks for the establishment of neuronal polarity. Curr. Opin. Cell Biol. 2020, 63, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Yamaji, K.; Sakai, H.; Okumura, Y.; Usui, S. Customizable neuroinformatics database system: XooNIps and its application to the pupil platform. Comput. Biol. Med. 2007, 37, 1036–1041. [Google Scholar] [CrossRef]
- Nishi, A.; Snyder, G.L.; Greengard, P. Bidirectional regulation of DARPP-32 phosphorylation by dopamine. J. Neurosci. 1997, 17, 8147–8155. [Google Scholar] [CrossRef] [Green Version]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Nagai, T.; Ahammad, R.U.; Kuroda, K.; Nakamuta, S.; Nakano, T.; Yukinawa, N.; Funahashi, Y.; Yamahashi, Y.; Amano, M.; et al. Balance between dopamine and adenosine signals regulates the PKA/Rap1 pathway in striatal medium spiny neurons. Neurochem. Int. 2019, 122, 8–18. [Google Scholar] [CrossRef]
- Amano, M.; Nishioka, T.; Yura, Y.; Kaibuchi, K. Identification of Protein Kinase Substrates by the Kinase-Interacting Substrate Screening (KISS) Approach. Curr. Protoc. Cell Biol. 2016, 72, 11–12. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, T.; Amano, M.; Funahashi, Y.; Tsuboi, D.; Yamahashi, Y.; Kaibuchi, K. In Vivo Identification of Protein Kinase Substrates by Kinase-Oriented Substrate Screening (KIOSS). Curr. Protoc. Chem. Biol. 2019, 11, e60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takano, T.; Wu, M.; Nakamuta, S.; Naoki, H.; Ishizawa, N.; Namba, T.; Watanabe, T.; Xu, C.; Hamaguchi, T.; Yura, Y.; et al. Discovery of long-range inhibitory signaling to ensure single axon formation. Nat. Commun. 2017, 8, 33. [Google Scholar] [CrossRef] [PubMed]
- Koike, R.; Amano, M.; Kaibuchi, K.; Ota, M. Protein kinases phosphorylate long disordered regions in intrinsically disordered proteins. Protein Sci. 2020, 29, 564–571. [Google Scholar] [CrossRef]
- Shohag, M.H.; Nishioka, T.; Ahammad, R.U.; Nakamuta, S.; Yura, Y.; Hamaguchi, T.; Kaibuchi, K.; Amano, M. Phosphoproteomic Analysis Using the WW and FHA Domains as Biological Filters. Cell Struct. Funct. 2015, 40, 95–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mok, J.; Im, H.; Snyder, M. Global identification of protein kinase substrates by protein microarray analysis. Nat. Protoc. 2009, 4, 1820–1827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, L.; Michaud, G.A.; Merkel, J.S.; Zhou, F.; Huang, J.; Mattoon, D.R.; Schweitzer, B. Protein kinase substrate identification on functional protein arrays. BMC Biotechnol. 2008, 8, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wardas, J. Neuroprotective role of adenosine in the CNS. Pol. J. Pharmacol. 2002, 54, 313–326. [Google Scholar] [PubMed]
- Kemp, J.M.; Powell, T.P. The structure of the caudate nucleus of the cat: Light and electron microscopy. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1971, 262, 383–401. [Google Scholar] [PubMed]
- Gerfen, C.R.; Engber, T.M.; Mahan, L.C.; Susel, Z.; Chase, T.N.; Monsma, F.J., Jr.; Sibley, D.R. D1 and D2 dopamine receptor-regulated gene expression of striatonigral and striatopallidal neurons. Science 1990, 250, 1429–1432. [Google Scholar] [CrossRef] [PubMed]
- Fink, J.S.; Weaver, D.R.; Rivkees, S.A.; Peterfreund, R.A.; Pollack, A.E.; Adler, E.M.; Reppert, S.M. Molecular cloning of the rat A2 adenosine receptor: Selective co-expression with D2 dopamine receptors in rat striatum. Brain Res. Mol. Brain Res. 1992, 14, 186–195. [Google Scholar] [CrossRef]
- Ferre, S.; O’Connor, W.T.; Svenningsson, P.; Bjorklund, L.; Lindberg, J.; Tinner, B.; Stromberg, I.; Goldstein, M.; Ogren, S.O.; Ungerstedt, U.; et al. Dopamine D1 receptor-mediated facilitation of GABAergic neurotransmission in the rat strioentopenduncular pathway and its modulation by adenosine A1 receptor-mediated mechanisms. Eur. J. Neurosci. 1996, 8, 1545–1553. [Google Scholar] [CrossRef] [PubMed]
- Kull, B.; Svenningsson, P.; Fredholm, B.B. Adenosine A(2A) receptors are colocalized with and activate g(olf) in rat striatum. Mol. Pharmacol. 2000, 58, 771–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAvoy, T.; Zhou, M.M.; Greengard, P.; Nairn, A.C. Phosphorylation of Rap1GAP, a striatally enriched protein, by protein kinase A controls Rap1 activity and dendritic spine morphology. Proc. Natl. Acad. Sci. USA 2009, 106, 3531–3536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snyder, G.L.; Allen, P.B.; Fienberg, A.A.; Valle, C.G.; Huganir, R.L.; Nairn, A.C.; Greengard, P. Regulation of phosphorylation of the GluR1 AMPA receptor in the neostriatum by dopamine and psychostimulants in vivo. J. Neurosci. 2000, 20, 4480–4488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.H.; Yamahashi, Y.; Kuroda, K.; Faruk, M.O.; Zhang, X.; Yamada, K.; Yamanaka, A.; Nagai, T.; Kaibuchi, K. Accumbal D2R-medium spiny neurons regulate aversive behaviors through PKA-Rap1 pathway. Neurochem. Int. 2021, 143, 104935. [Google Scholar] [CrossRef] [PubMed]
- Poolos, N.P.; Bullis, J.B.; Roth, M.K. Modulation of h-channels in hippocampal pyramidal neurons by p38 mitogen-activated protein kinase. J. Neurosci. 2006, 26, 7995–8003. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Bullis, J.B.; Lau, I.H.; Jones, T.D.; Warner, L.N.; Poolos, N.P. Downregulation of dendritic HCN channel gating in epilepsy is mediated by altered phosphorylation signaling. J. Neurosci. 2010, 30, 6678–6688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzgerald, E.M. Regulation of voltage-dependent calcium channels in rat sensory neurones involves a Ras-mitogen-activated protein kinase pathway. J. Physiol. 2000, 527, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Cole, A.R.; Knebel, A.; Morrice, N.A.; Robertson, L.A.; Irving, A.J.; Connolly, C.N.; Sutherland, C. GSK-3 phosphorylation of the Alzheimer epitope within collapsin response mediator proteins regulates axon elongation in primary neurons. J. Biol. Chem. 2004, 279, 50176–50180. [Google Scholar] [CrossRef] [Green Version]
- Uchida, Y.; Ohshima, T.; Sasaki, Y.; Suzuki, H.; Yanai, S.; Yamashita, N.; Nakamura, F.; Takei, K.; Ihara, Y.; Mikoshiba, K.; et al. Semaphorin3A signalling is mediated via sequential Cdk5 and GSK3beta phosphorylation of CRMP2: Implication of common phosphorylating mechanism underlying axon guidance and Alzheimer’s disease. Genes Cells 2005, 10, 165–179. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, T.; Kawano, Y.; Arimura, N.; Kawabata, S.; Kikuchi, A.; Kaibuchi, K. GSK-3beta regulates phosphorylation of CRMP-2 and neuronal polarity. Cell 2005, 120, 137–149. [Google Scholar] [CrossRef] [Green Version]
- Cole, A.R.; Noble, W.; van Aalten, L.; Plattner, F.; Meimaridou, R.; Hogan, D.; Taylor, M.; LaFrancois, J.; Gunn-Moore, F.; Verkhratsky, A.; et al. Collapsin response mediator protein-2 hyperphosphorylation is an early event in Alzheimer’s disease progression. J. Neurochem. 2007, 103, 1132–1144. [Google Scholar] [CrossRef]
- Castillon, C.; Gonzalez, L.; Domenichini, F.; Guyon, S.; Da Silva, K.; Durand, C.; Lestaevel, P.; Vaillend, C.; Laroche, S.; Barnier, J.V.; et al. The intellectual disability PAK3 R67C mutation impacts cognitive functions and adult hippocampal neurogenesis. Hum. Mol. Genet. 2020, 29, 1950–1968. [Google Scholar] [CrossRef] [PubMed]
- Pavlowsky, A.; Chelly, J.; Billuart, P. Emerging major synaptic signaling pathways involved in intellectual disability. Mol. Psychiatry 2012, 17, 682–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiwer, M.; Xu, Q.; Gale, J.P.; Lewis, M.; Campbell, A.J.; Schroeder, F.A.; Van de Bittner, G.C.; Walk, M.; Amaya, A.; Su, P.; et al. Functionally Biased D2R Antagonists: Targeting the beta-Arrestin Pathway to Improve Antipsychotic Treatment. ACS Chem. Biol. 2018, 13, 1038–1047. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, K.; Jeevakumar, V.; Nakao, K. Spatial and temporal boundaries of NMDA receptor hypofunction leading to schizophrenia. NPJ Schizophr. 2017, 3, 7. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method/Stimulation | Kinase | No. of Phos. Site |
|---|---|---|
| KIOSS (1433z)/Forskolin | PKA, MAPK and other | 278 |
| KIOSS (1433z)/OA&U0126 | MAPK and other | 324 |
| KIOSS (1433z)/D1R Agonist | PKA, MAPK and other | 185 |
| KIOSS (1433z)/A2AR Agonist | PKA, MAPK and other | 62 |
| KISS | CaMKI, CDK5, FYN, LYN, MAPK1, PAK7, PKA, PKN, ROCK2 | 3597 |
| Protoarray | AKT1, CaMK2A, CDK5, LRRK2, LYN, MAPK1, PRKCA, PRKACA, ROCK2 | 1782 |
| Outsource | AurA, AurB, CaMKI, CaMKII, CaMK4, CDK5, GSK3B, LYN, MAPK, MARK1, PKA, AMPK, PRKCA, PRKCE, ROCK, STK11(LKB1) | 3736 |
| Motif | Identified Phosphoprotein and Phosphorylation Site | |||||
|---|---|---|---|---|---|---|
| PKA | Arhgap21 | Arhgap21 | Arhgap21 | Arhgap23 | Brsk1 | Brsk2 |
| S472 | S851 | S875 | S401 | S324 | S383 | |
| Bsn | Bsn | Cacna1b | Ccdc177 | Ccny | Cdk17 | |
| S1987 | S2860 | S2219 | S303 | S301 | S180 | |
| Chrna4 | Fam126b | Gpr75 | Hcn2 | Kcnh7 | Kif21b | |
| S540 | S521 | S317 | S840 | S174 | S1150 | |
| Madd | Mark1 | Nedd4l | Nf1 | Prickle2 | Prrt3 | |
| S1038 | S394 | T353 | S2524 | S752 | S846 | |
| Rap1gap | Rims1 | Sik3 | Syn1 | Taf4b | Ttc32 | |
| S245 | S887 | S454 | S427 | S707 | S24 | |
| MAPK | Ablim1 | Arhgap21 | Bsn | Cacna1e | Ccdc177 | Cdk17 |
| S89 | S42 | Y3020 | S791 | S308 | T11 | |
| Chrna4 | Chrna4 | Hcn3 | Lrfn4 | Madd | Madd | |
| S540 | S543 | S633 | S627 | S1038 | T1045 | |
| Nedd4l | Prrt3 | Rap1gap | Sipa1l1 | Sorbs2 | Spata2 | |
| S329 | S834 | S213 | S1528 | S944 | S247 | |
| Syn1 | ||||||
| S427 | ||||||
| Others | Arhgap21 | Atp6v1h | Cep170 | Gpr75 | Kcnh2 | Khnyn |
| S856 | Y125 | S353 | T313 | S322 | S43 | |
| Madd | Mapk8 | Rsl1d1 | Rsl1d1 | Sh2d5 | Ttc32 | |
| S1089 | S210 | T314 | S316 | S126 | T18 | |
| Identified MAPK Candidate Substrates List | |||||||
|---|---|---|---|---|---|---|---|
| 2010300C02Rik | Atg9a | Cdk18 | Epb4.1l3 | Lppr3 | Osbpl6 | Rasgrp2 | Ssh2 |
| Aak1 | Bai1 | Cdkl5 | Epb49 | Lrfn4 | Pak7 | Rem2 | Stim1 |
| Abi1 | Baiap2 | Cep170 | Erc2 | Lrrc7 | Panx2 | Rims1 | Syn1 |
| Abi2 | Bcr | Chrna4 | Etl4 | Madd | Pclo | Rims2 | Syn2 |
| Agap2 | Begain | Clasp2 | Fam171a1 | Map3k5 | Pfkfb2 | Rph3a | Syn3 |
| Agfg1 | Brsk1 | Crtc2 | Fam171a2 | Map4 | Pip5k1c | Rrad | Synpo |
| Akap6 | Brsk2 | Ctnnd2 | Fam171b | Mark1 | Pitpnm3 | Sh2d5 | Syt7 |
| Als2 | Bsn | Cyth3 | Frmd4a | Mark3 | Plekha5 | Shank3 | Tanc2 |
| Ampd2 | C17orf59 | Dab2ip | Gab2 | Mast1 | Plekho2 | Shisa7 | Tpd52 |
| Ank2 | C2cd4c | Dennd1a | Git1 | Mast3 | Pola1 | Sipa1l1 | Tpd52l1 |
| Ankrd34a | Cacna1b | Dennd4c | Gm1568 | Mbp | Ppfia2 | Sipa1l2 | Tsc2 |
| Ankrd34b | Cacna1e | Dgkq | Gm15800 | Mink1 | Ppfia3 | Slc4a4 | Uhrf1bp1l |
| Anks1b | Camk2g | Dlgap2 | Gpr158 | Mllt4 | Ppfia4 | Sorbs1 | Ulk1 |
| Arhgap21 | Camkk1 | Dlgap3 | Hcn2 | Mprip | Prrt3 | Sorbs2 | Ulk2 |
| Arhgap23 | Camkk2 | Dos | Hcn3 | Nav1 | Psd | Spata2 | Usp31 |
| Arhgap32 | Camsap2 | Dpysl2 | Iqsec1 | Nav3 | Psd3 | Specc1 | Usp8 |
| Arhgap39 | Caskin1 | Dpysl5 | Kcnb1 | Ndel1 | Rab11fip2 | Speg | Wdr47 |
| Arhgef2 | Ccdc22 | Dstn | Kiaa0284 | Nelf | Ralgapa1 | Srcin1 | |
| Arhgef6 | Ccny | Dtna | Kiaa0528 | Nhsl2 | Rap1gap | Srgap3 | |
| Atat1 | Ccnyl1 | Eif3d | Kiaa1211 | Nyap2 | Rapgef2 | Ssfa2 | |
| Total 157 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahammad, R.U.; Nishioka, T.; Yoshimoto, J.; Kannon, T.; Amano, M.; Funahashi, Y.; Tsuboi, D.; Faruk, M.O.; Yamahashi, Y.; Yamada, K.; et al. KANPHOS: A Database of Kinase-Associated Neural Protein Phosphorylation in the Brain. Cells 2022, 11, 47. https://doi.org/10.3390/cells11010047
Ahammad RU, Nishioka T, Yoshimoto J, Kannon T, Amano M, Funahashi Y, Tsuboi D, Faruk MO, Yamahashi Y, Yamada K, et al. KANPHOS: A Database of Kinase-Associated Neural Protein Phosphorylation in the Brain. Cells. 2022; 11(1):47. https://doi.org/10.3390/cells11010047
Chicago/Turabian StyleAhammad, Rijwan Uddin, Tomoki Nishioka, Junichiro Yoshimoto, Takayuki Kannon, Mutsuki Amano, Yasuhiro Funahashi, Daisuke Tsuboi, Md. Omar Faruk, Yukie Yamahashi, Kiyofumi Yamada, and et al. 2022. "KANPHOS: A Database of Kinase-Associated Neural Protein Phosphorylation in the Brain" Cells 11, no. 1: 47. https://doi.org/10.3390/cells11010047