
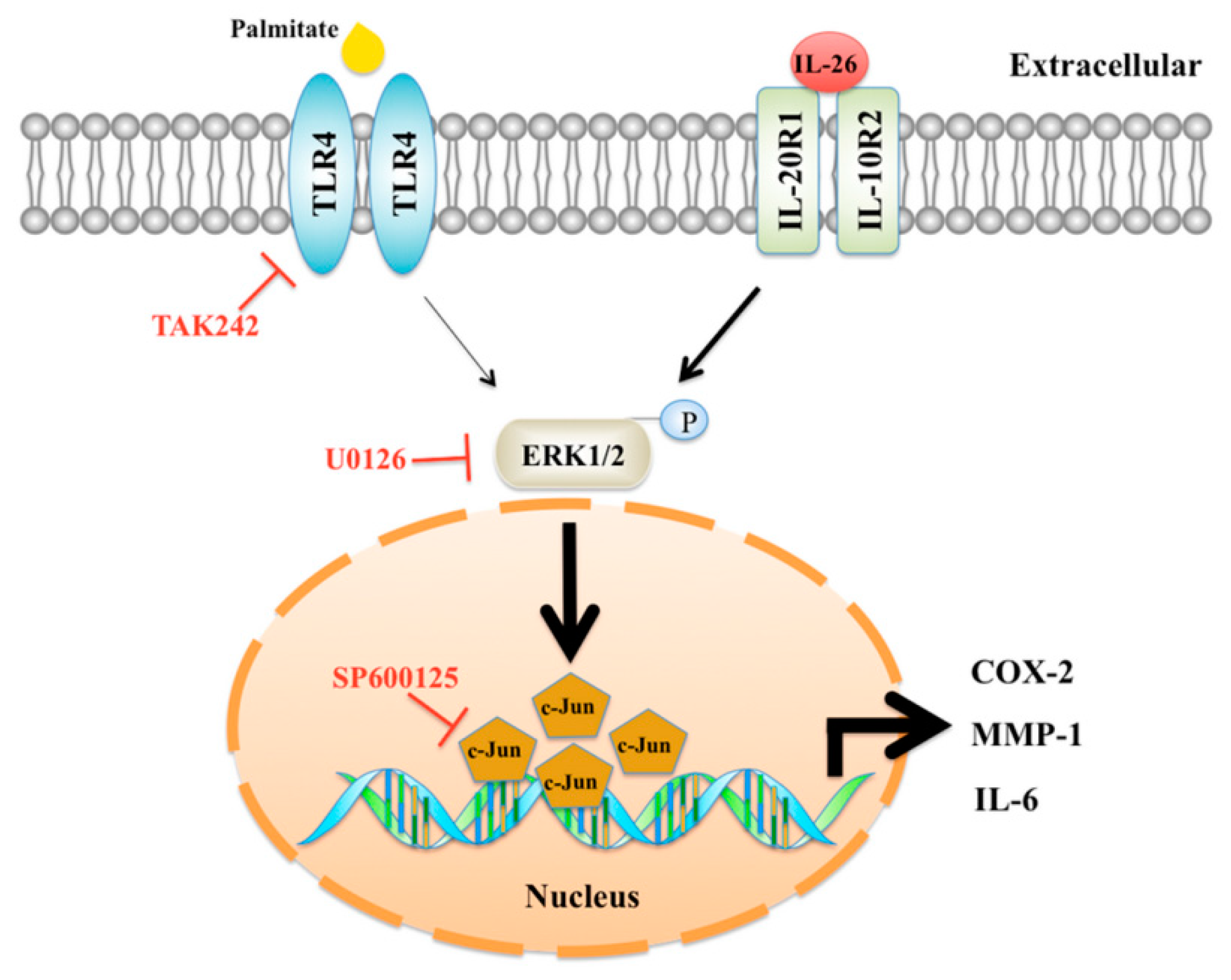
Interleukin-26 Has Synergistic Catabolic Effects with Palmitate in Human Articular Chondrocytes via the TLR4-ERK1/2-c-Jun Signaling Pathway

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Isolation and Culture of HACs
2.3. Palmitate/BSA Complex Solution
2.4. Cell Viability
2.5. Cell Stimulation and Immunoblotting
2.6. Extraction of Cytoplasmic and Nuclear Protein
2.7. Preparation of Cartilage Explants
2.8. Staining with Safranin O and 1,9-Dimethylmethylene Blue for Analysis of Cartilage Degradation
2.9. Statistical Analysis
3. Results
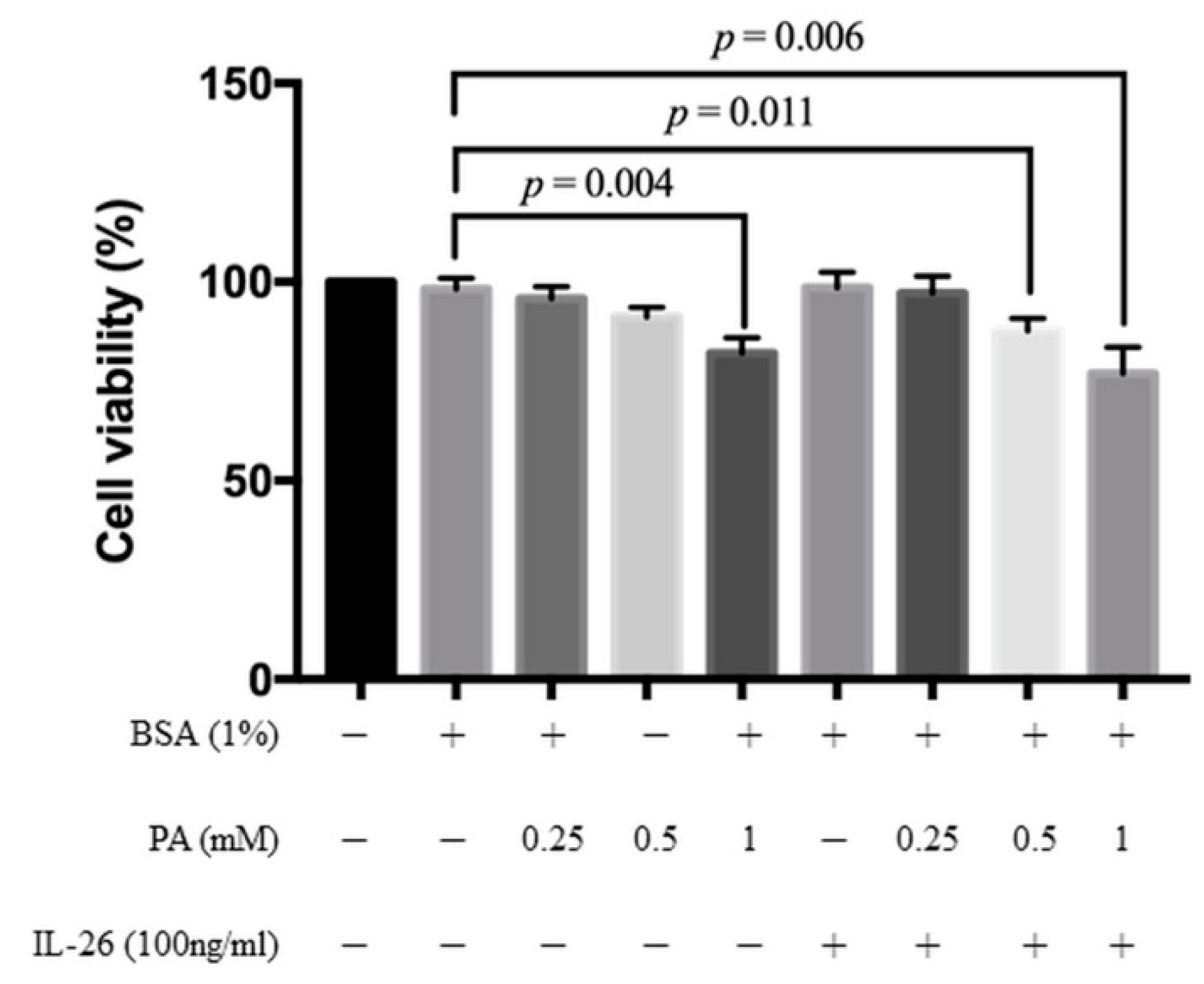
3.1. Cytotoxicity of IL-26 and Palmitate in HACs
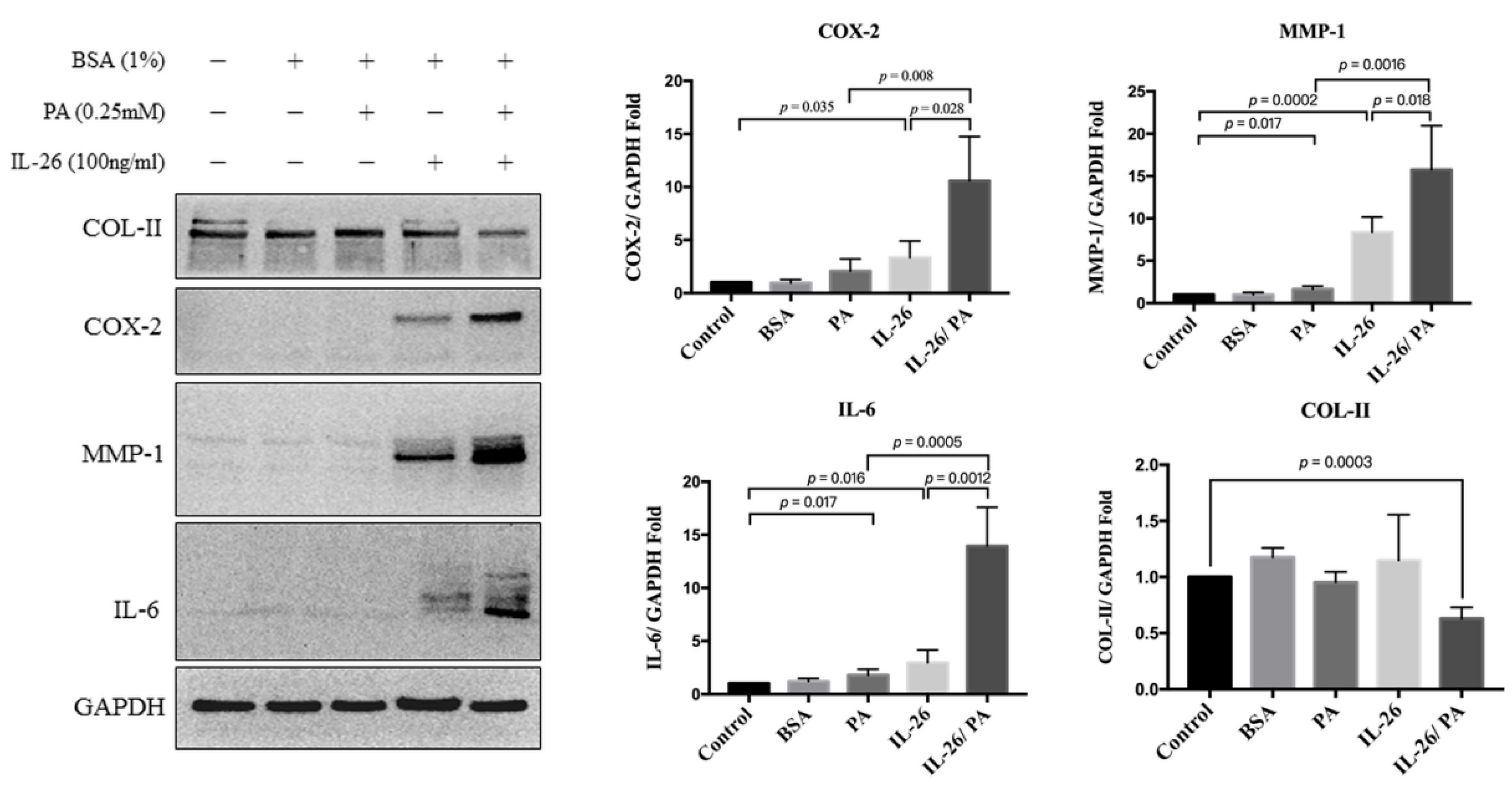
3.2. Effects of IL-26 and Palmitate on HACs
3.3. Effects of IL-26 and Palmitate on Signaling Pathways in HACs
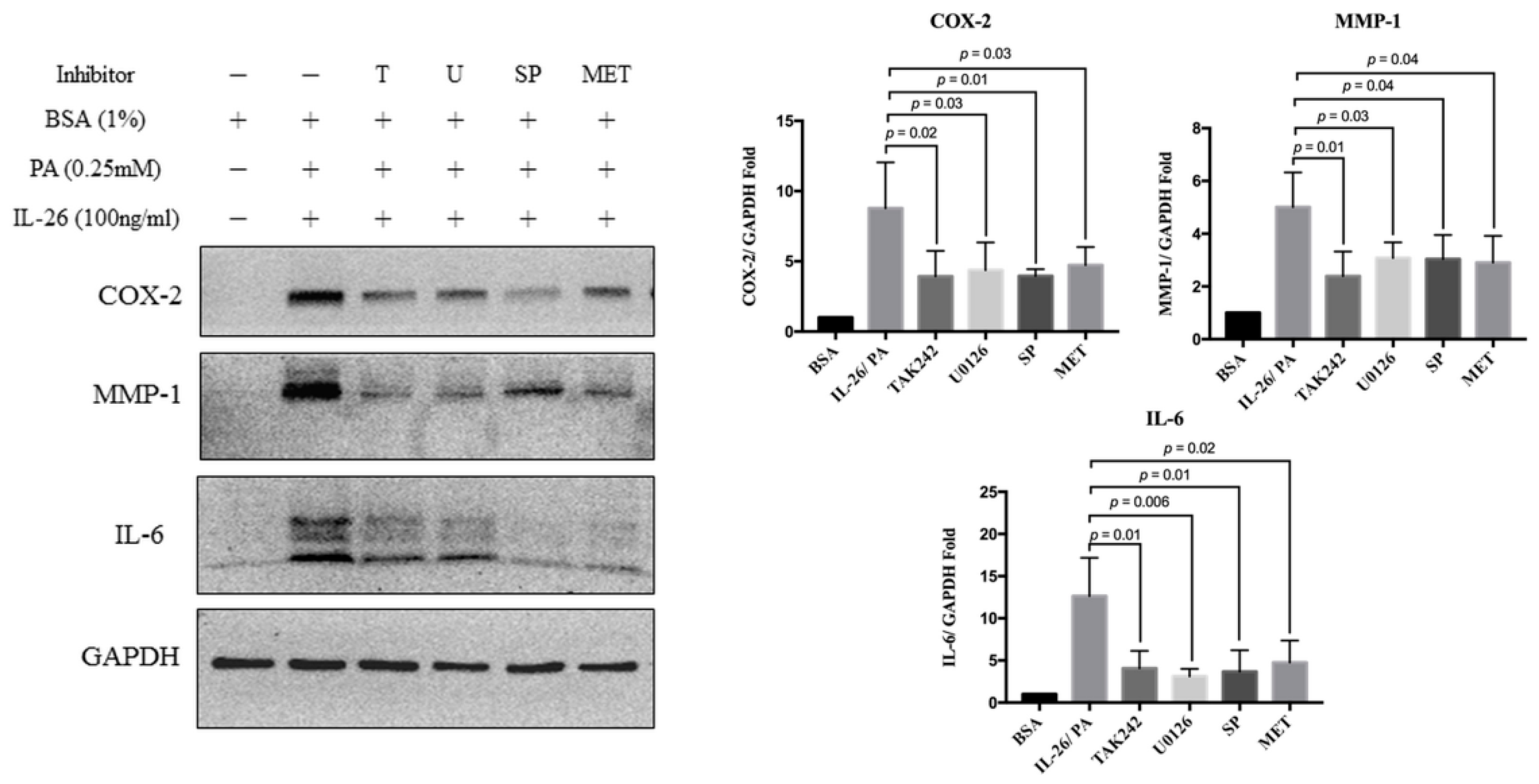
3.4. Synergistic Catabolic Effects of IL-26 and Palmitate via the TLR4/ERK1/2-c-Jun Signaling Pathway
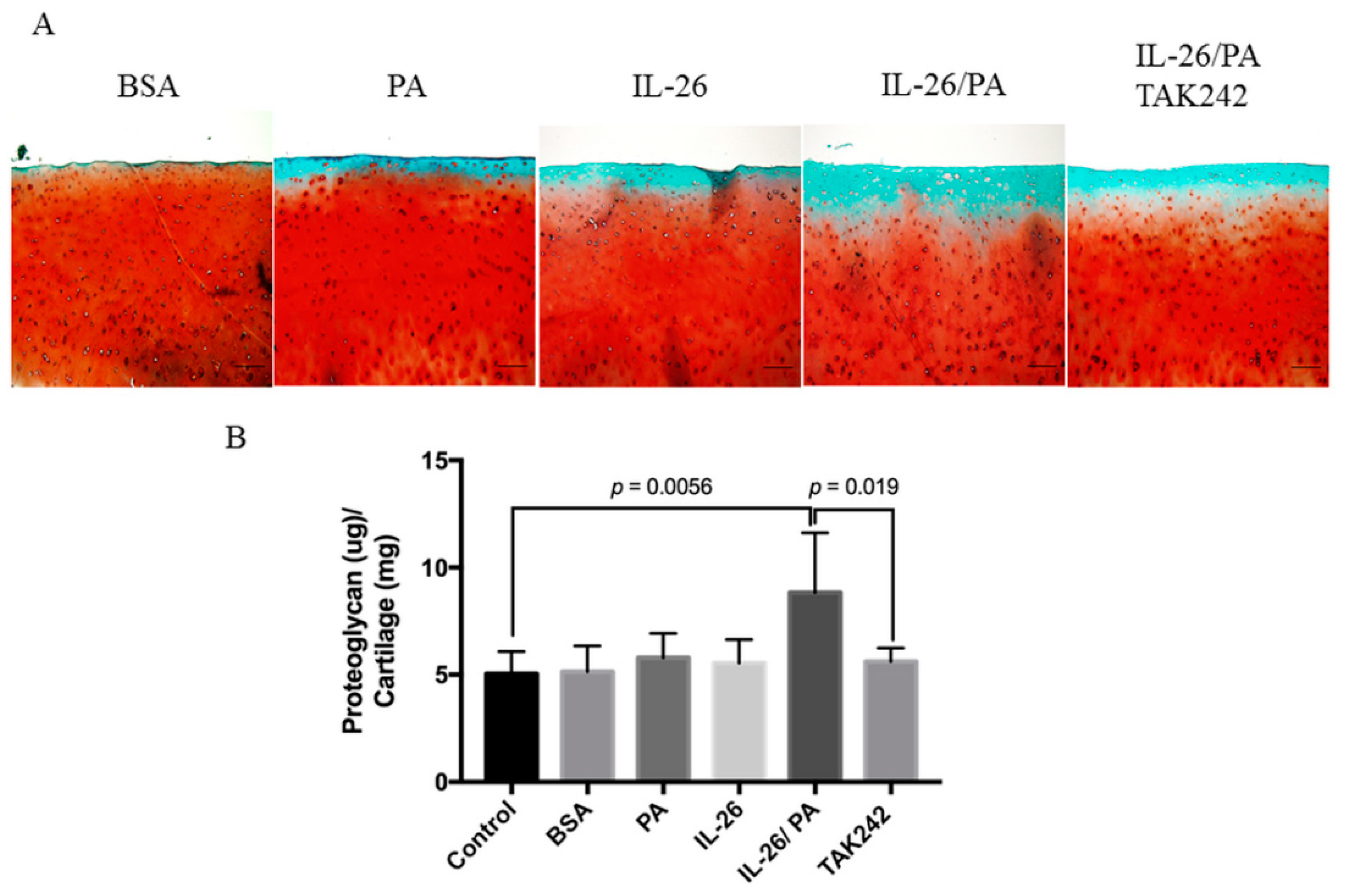
3.5. Attenuation of Palmitate- and IL-26-Induced Matrix Degradation of Human Cartilage Explants by TAK242
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Knappe, A.; Hör, S.; Wittmann, S.; Fickenscher, H. Induction of a novel cellular homolog of interleukin-10, ak155, by transformation of t lymphocytes with herpesvirus saimiri. J. Virol. 2000, 74, 3881–3887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheikh, F.; Baurin, V.V.; Lewis-Antes, A.; Shah, N.K.; Smirnov, S.V.; Anantha, S.; Dickensheets, H.; Dumoutier, L.; Renauld, J.-C.; Zdanov, A. Cutting edge: Il-26 signals through a novel receptor complex composed of il-20 receptor 1 and il-10 receptor 2. J. Immunol. 2004, 172, 2006–2010. [Google Scholar] [CrossRef] [Green Version]
- Dambacher, J.; Beigel, F.; Zitzmann, K.; De Toni, E.; Göke, B.; Diepolder, H.M.; Auernhammer, C.J.; Brand, S. The role of the novel th17 cytokine il-26 in intestinal inflammation. Gut 2009, 58, 1207–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savchenko, L.; Mykytiuk, M.; Cinato, M.; Tronchere, H.; Kunduzova, O.; Kaidashev, I. Il-26 in the induced sputum is associated with the level of systemic inflammation, lung functions and body weight in copd patients. Int. J. Chron. Obstruct. Pulmon. Dis. 2018, 13, 2569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, H.; Lai, X.; Li, J.; Huang, L.; Liu, Y.; Cao, J. Interleukin-26 is overexpressed in human sepsis and contributes to inflammation, organ injury, and mortality in murine sepsis. Crit. Care 2019, 23, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Corvaisier, M.; Delneste, Y.; Jeanvoine, H.; Preisser, L.; Blanchard, S.; Garo, E.; Hoppe, E.; Barré, B.; Audran, M.; Bouvard, B. Il-26 is overexpressed in rheumatoid arthritis and induces proinflammatory cytokine production and th17 cell generation. PLoS Biol. 2012, 10, e1001395. [Google Scholar] [CrossRef]
- Khaodhiar, L.; McCowen, K.C.; Blackburn, G.L. Obesity and its comorbid conditions. Clin. Cornerstone 1999, 2, 17–31. [Google Scholar] [CrossRef]
- Karpe, F.; Dickmann, J.R.; Frayn, K.N. Fatty acids, obesity, and insulin resistance: Time for a reevaluation. Diabetes 2011, 60, 2441–2449. [Google Scholar] [CrossRef] [Green Version]
- Suganami, T.; Tanimoto-Koyama, K.; Nishida, J.; Itoh, M.; Yuan, X.; Mizuarai, S.; Kotani, H.; Yamaoka, S.; Miyake, K.; Aoe, S. Role of the toll-like receptor 4/nf-κb pathway in saturated fatty acid–induced inflammatory changes in the interaction between adipocytes and macrophages. Arter. Thromb. Vasc. Biol. 2007, 27, 84–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, H.-J.; Kim, H.-S.; Hwang, D.H.; Quon, M.J.; Kim, J.-a. Toll-like receptor 2 mediates high-fat diet-induced impairment of vasodilator actions of insulin. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E1077–E1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasu, M.R.; Jialal, I. Free fatty acids in the presence of high glucose amplify monocyte inflammation via toll-like receptors. Am. J. Physiol. Endocrinol. Metab. 2011, 300, E145–E154. [Google Scholar] [CrossRef] [Green Version]
- Colmegna, I.; Hitchon, C.A.; Bardales, M.C.B.; Puri, L.; Bartlett, S.J. High rates of obesity and greater associated disability among people with rheumatoid arthritis in canada. Clin. Rheumatol. 2016, 35, 457–460. [Google Scholar] [CrossRef]
- de Resende Guimaraes, M.F.B.; Rodrigues, C.E.M.; Gomes, K.W.P.; Machado, C.J.; Brenol, C.V.; Krampe, S.F.; de Andrade, N.P.B.; Kakehasi, A.M. High prevalence of obesity in rheumatoid arthritis patients: Association with disease activity, hypertension, dyslipidemia and diabetes, a multi-center study. Adv. Rheumatol. 2019, 59, 44. [Google Scholar] [CrossRef] [PubMed]
- Durcan, L.; Wilson, F.; Conway, R.; Cunnane, G.; O’Shea, F.D. Increased body mass index in ankylosing spondylitis is associated with greater burden of symptoms and poor perceptions of the benefits of exercise. J. Rheumatol. 2012, 39, 2310–2314. [Google Scholar] [CrossRef]
- Peng, Y.-J.; Lee, C.-H.; Wang, C.-C.; Salter, D.M.; Lee, H.-S. Pycnogenol attenuates the inflammatory and nitrosative stress on joint inflammation induced by urate crystals. Free Radic. Biol. Med. 2012, 52, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, G.I.; Langley, K.G.; Berglund, N.A.; Kammoun, H.L.; Reibe, S.; Estevez, E.; Weir, J.; Mellett, N.A.; Pernes, G.; Conway, J.R. Evidence that tlr4 is not a receptor for saturated fatty acids but mediates lipid-induced inflammation by reprogramming macrophage metabolism. Cell Metab. 2018, 27, 1096–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritzker, K.P.; Gay, S.; Jimenez, S.; Ostergaard, K.; Pelletier, J.-P.; Revell, P.; Salter, D.; Van den Berg, W. Osteoarthritis cartilage histopathology: Grading and staging. Osteoarthr. Cartil. 2006, 14, 13–29. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.-C.; Hung, L.-F.; Wu, W.-L.; Chang, D.-M.; Huang, C.-Y.; Lai, J.-H.; Ho, L.-J. Chondroprotective effects and mechanisms of resveratrol in advanced glycation end products-stimulated chondrocytes. Arthritis Res. Ther. 2010, 12, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.-J.; Wang, C.-Y.; Lin, Y.-H.; Lin, G.-J.; Huang, S.-H.; Shyu, J.-F.; Sytwu, H.-K.; Cheng, C.-P. Interleukin 26 suppresses receptor activator of nuclear factor κb ligand induced osteoclastogenesis via down-regulation of nuclear factor of activated t-cells, cytoplasmic 1 and nuclear factor κb activity. Rheumatology 2016, 55, 2074–2083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sindhu, S.; Al-Roub, A.; Koshy, M.; Thomas, R.; Ahmad, R. Palmitate-induced mmp-9 expression in the human monocytic cells is mediated through the tlr4-myd88 dependent mechanism. Cell Physiol. Biochem. 2016, 39, 889–900. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qian, Y.; Fang, Q.; Zhong, P.; Li, W.; Wang, L.; Fu, W.; Zhang, Y.; Xu, Z.; Li, X. Saturated palmitic acid induces myocardial inflammatory injuries through direct binding to tlr4 accessory protein md2. Nat. Commun. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Hör, S.; Pirzer, H.; Dumoutier, L.; Bauer, F.; Wittmann, S.; Sticht, H.; Renauld, J.-C.; de Waal Malefyt, R.; Fickenscher, H. The t-cell lymphokine interleukin-26 targets epithelial cells through the interleukin-20 receptor 1 and interleukin-10 receptor 2 chains. J. Biol. Chem. 2004, 279, 33343–33351. [Google Scholar] [CrossRef] [Green Version]
- Bailey, C.J. Metformin: Historical overview. Diabetologia 2017, 60, 1566–1576. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Hussain, S.M.; Wluka, A.E.; Lim, Y.Z.; Abram, F.; Pelletier, J.-P.; Martel-Pelletier, J.; Cicuttini, F.M. Association between metformin use and disease progression in obese people with knee osteoarthritis: Data from the osteoarthritis initiative—A prospective cohort study. Arthritis Res. Ther. 2019, 21, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peixoto, L.G.; Teixeira, R.R.; Vilela, D.D.; Barbosa, L.N.; Caixeta, D.C.; Deconte, S.R.; de Araújo, F.d.A.; Sabino-Silva, R.; Espindola, F.S. Metformin attenuates the tlr4 inflammatory pathway in skeletal muscle of diabetic rats. Acta Diabetol. 2017, 54, 943–951. [Google Scholar] [CrossRef]
- Tengvall, S.; Che, K.F.; Lindén, A. Interleukin-26: An emerging player in host defense and inflammation. J. Innate Immun. 2016, 8, 15–22. [Google Scholar] [CrossRef]
- Meller, S.; Di Domizio, J.; Voo, K.S.; Friedrich, H.C.; Chamilos, G.; Ganguly, D.; Conrad, C.; Gregorio, J.; Le Roy, D.; Roger, T. Th 17 cells promote microbial killing and innate immune sensing of DNA via interleukin 26. Nat. Immunol. 2015, 16, 970–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larochette, V.; Miot, C.; Poli, C.; Beaumont, E.; Roingeard, P.; Fickenscher, H.; Jeannin, P.; Delneste, Y. Il-26, a cytokine with roles in extracellular DNA-induced inflammation and microbial defense. Front. Immunol. 2019, 10, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Che, K.F.; Tengvall, S.; Levänen, B.; Silverpil, E.; Smith, M.E.; Awad, M.; Vikström, M.; Palmberg, L.; Qvarfordt, I.; Sköld, M. Interleukin-26 in antibacterial host defense of human lungs. Effects on neutrophil mobilization. Am. J. Respir. Crit. Care Med. 2014, 190, 1022–1031. [Google Scholar] [CrossRef]
- Zhou, H.; Urso, C.; Jadeja, V. Saturated fatty acids in obesity-associated inflammation. J. Inflamm. Res. 2020, 13, 1. [Google Scholar] [CrossRef] [Green Version]
- Quan, J.; Liu, J.; Gao, X. Palmitate induces interleukin-8 expression in human aortic vascular smooth muscle cells via toll-like receptor 4/nf-κb pathway. J. Diabetes 2013. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, D.; Wang, F.; Liu, S.; Zhao, S.; Ling, E.-A.; Hao, A. Saturated fatty acids activate microglia via toll-like receptor 4/nf-κb signalling. Br. J. Nutr. 2012, 107, 229–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Sohn, K.H.; Rhee, S.H.; Hwang, D. Saturated fatty acids, but not unsaturated fatty acids, induce the expression of cyclooxygenase-2 mediated through toll-like receptor 4. J. Biol. Chem. 2001, 276, 16683–16689. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.E.; Gabler, N.K.; Walker-Daniels, J.; Spurlock, M.E. Tlr-4 deficiency selectively protects against obesity induced by diets high in saturated fat. Obesity 2008, 16, 1248–1255. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Garcia, O.; Rogers, N.H.; Smith, R.G.; Lotz, M.K. Palmitate has proapoptotic and proinflammatory effects on articular cartilage and synergizes with interleukin-1. Arthritis Rheumatol. 2014, 66, 1779–1788. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, R.; Al-Roub, A.; Kochumon, S.; Akther, N.; Thomas, R.; Kumari, M.; Koshy, M.S.; Tiss, A.; Hannun, Y.A.; Tuomilehto, J. The synergy between palmitate and tnf-α for ccl2 production is dependent on the trif/irf3 pathway: Implications for metabolic inflammation. J. Immunol. 2018, 200, 3599–3611. [Google Scholar] [CrossRef]
- Saisho, Y. Metformin and inflammation: Its potential beyond glucose-lowering effect. Endocr. Metab. Immune. Disord. Drug Targets 2015, 15, 196–205. [Google Scholar] [CrossRef]
- Yerevanian, A.; Soukas, A.A. Metformin: Mechanisms in human obesity and weight loss. Curr. Obes. Rep. 2019, 8, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.H.; Chung, C.H.; Lee, C.H.; Hsieh, C.H.; Hung, Y.J.; Lin, F.H.; Tsao, C.H.; Hsieh, P.S.; Chien, W.C. Combination cox-2 inhibitor and metformin attenuate rate of joint replacement in osteoarthritis with diabetes: A nationwide, retrospective, matched-cohort study in taiwan. PLoS ONE 2018, 13, e0191242. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-T.; Wang, C.-C.; Cheng, C.-P.; Liu, F.-C.; Lee, C.-H.; Lee, H.-S.; Peng, Y.-J. Interleukin-26 Has Synergistic Catabolic Effects with Palmitate in Human Articular Chondrocytes via the TLR4-ERK1/2-c-Jun Signaling Pathway. Cells 2021, 10, 2500. https://doi.org/10.3390/cells10092500
Chen Y-T, Wang C-C, Cheng C-P, Liu F-C, Lee C-H, Lee H-S, Peng Y-J. Interleukin-26 Has Synergistic Catabolic Effects with Palmitate in Human Articular Chondrocytes via the TLR4-ERK1/2-c-Jun Signaling Pathway. Cells. 2021; 10(9):2500. https://doi.org/10.3390/cells10092500
Chicago/Turabian StyleChen, Yi-Ting, Chih-Chien Wang, Chia-Pi Cheng, Feng-Cheng Liu, Chian-Her Lee, Herng-Sheng Lee, and Yi-Jen Peng. 2021. "Interleukin-26 Has Synergistic Catabolic Effects with Palmitate in Human Articular Chondrocytes via the TLR4-ERK1/2-c-Jun Signaling Pathway" Cells 10, no. 9: 2500. https://doi.org/10.3390/cells10092500