
Mitochondrial DNA Haplogroup Related to the Prevalence of Helicobacter pylori

 ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects and Sample Collection
2.2. Preparation of H. pylori
2.3. mtDNA Sequencing by Miseq
2.4. Infection of Cultured Fibroblasts with H. pylori
2.5. Assessment of Apoptosis
2.6. Assessment of Mitochondrial Respiratory Functions
2.7. Statistics
3. Results
3.1. Relationship between mtDNA Haplogroups and H. pylori Prevalence
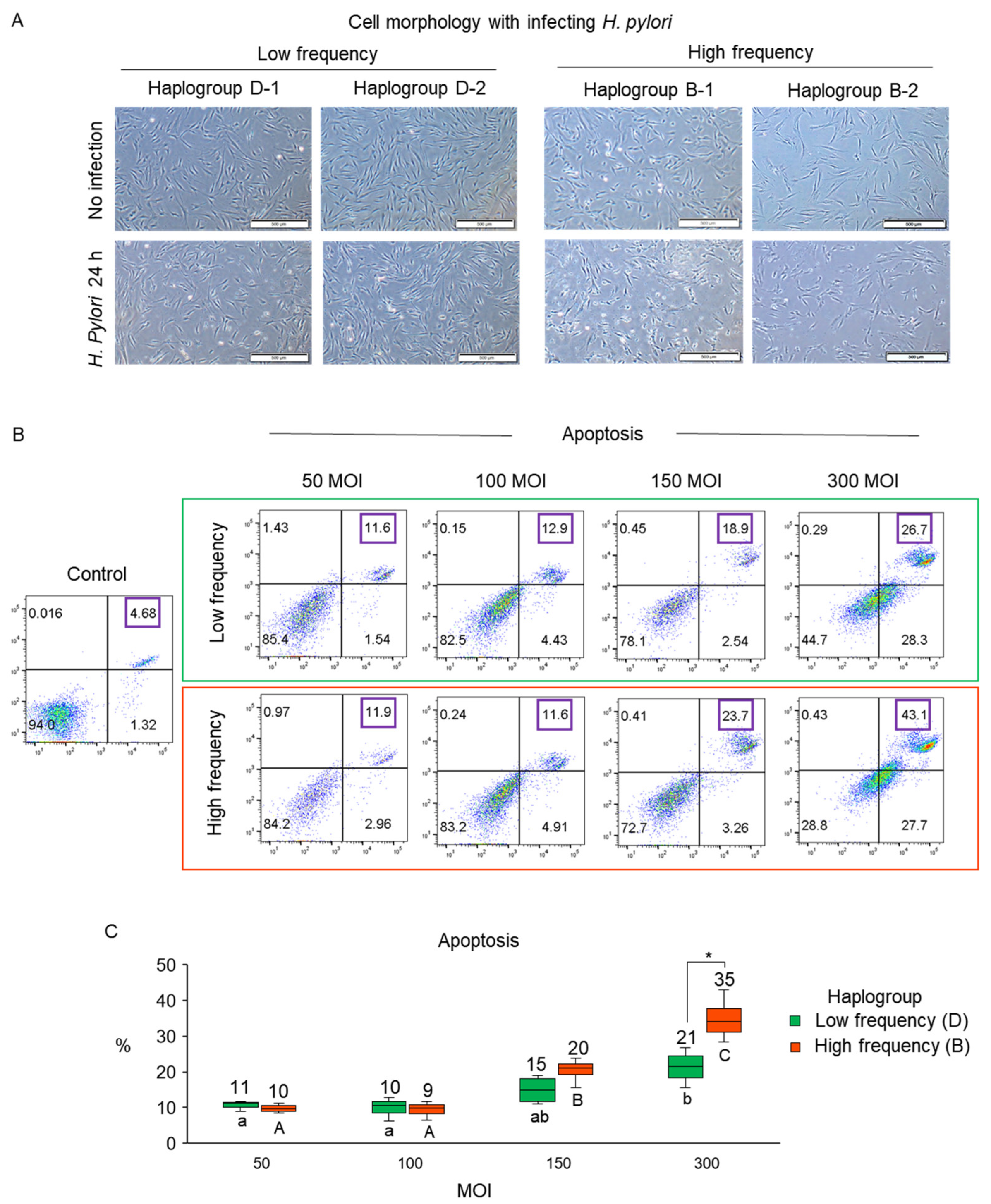
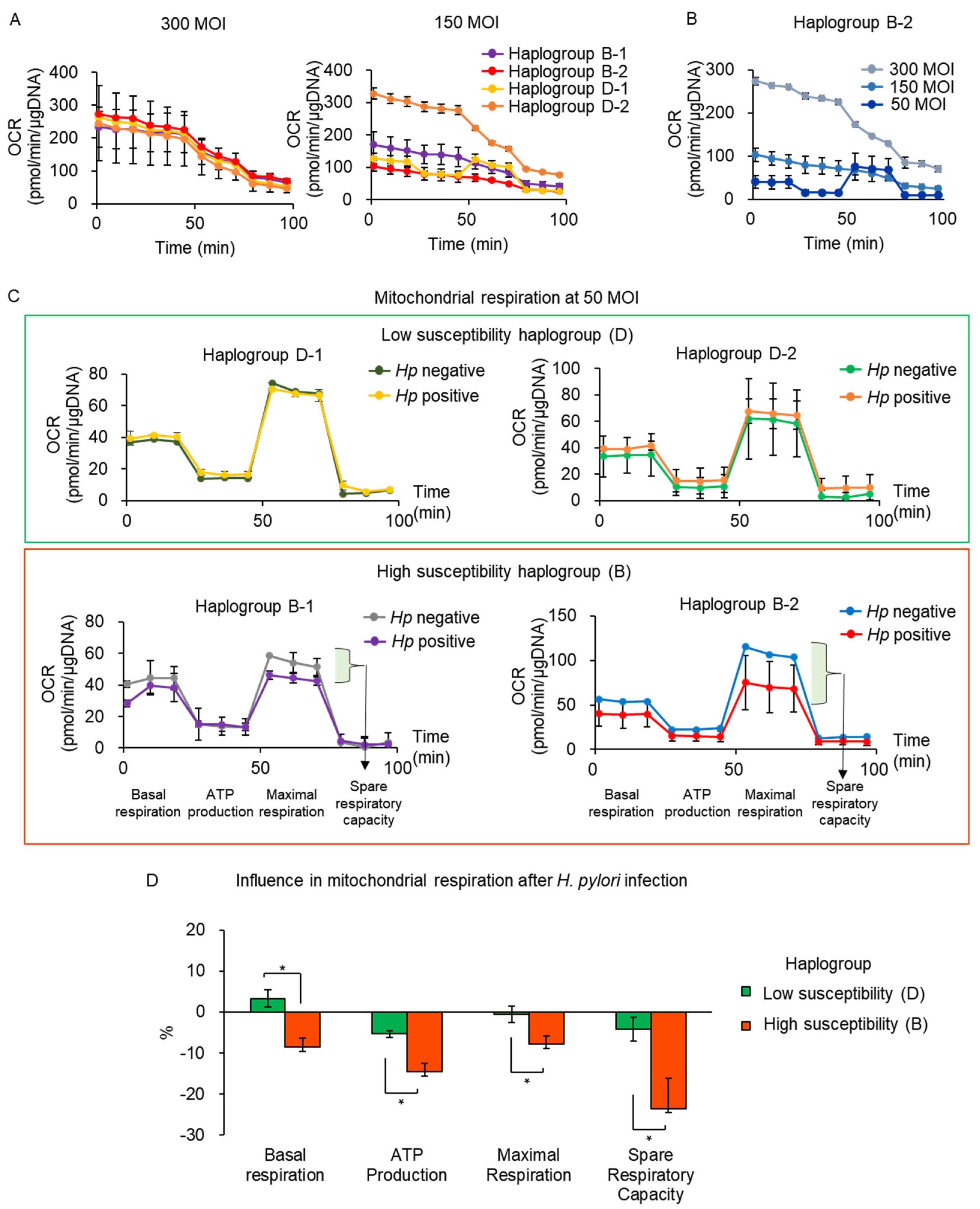
3.2. Validation of Susceptibility to H. pylori Infection Associated with mtDNA Haplogroup Prevalence
3.3. mtDNA Mutations with H. pylori Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Machado, A.M.; Desler, C.; Boggild, S.; Strickertsson, J.A.; Friis-Hansen, L.; Figueiredo, C.; Seruca, R.; Rasmussen, L.J. Helicobacter pylori infection affects mitochondrial function and DNA repair, thus, mediating genetic instability in gastric cells. Mech. Ageing Dev. 2013, 134, 460–466. [Google Scholar] [CrossRef]
- Figueiredo, C.; Machado, J.C.; Pharoah, P.; Seruca, R.; Sousa, S.; Carvalho, R.; Capelinha, A.F.; Quint, W.; Caldas, C.; van Doorn, L.J.; et al. Helicobacter pylori and interleukin 1 genotyping: An opportunity to identify high-risk individuals for gastric carcinoma. J. Natl. Cancer Inst. 2002, 94, 1680–1687. [Google Scholar] [CrossRef] [Green Version]
- Sjomina, O.; Heluwaert, F.; Moussata, D.; Leja, M. Helicobacter pylori infection and nonmalignant diseases. Helicobacter 2017, 22 (Suppl. 1). [Google Scholar] [CrossRef] [PubMed]
- Hooi, J.K.Y.; Lai, W.Y.; Ng, W.K.; Suen, M.M.Y.; Underwood, F.E.; Tanyingoh, D.; Malfertheiner, P.; Graham, D.Y.; Wong, V.W.S.; Wu, J.C.Y.; et al. Global Prevalence of Helicobacter pylori Infection: Systematic Review and Meta-Analysis. Gastroenterology 2017, 153, 420–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamani, M.; Ebrahimtabar, F.; Zamani, V.; Miller, W.H.; Alizadeh-Navaei, R.; Shokri-Shirvani, J.; Derakhshan, M.H. Systematic review with meta-analysis: The worldwide prevalence of Helicobacter pylori infection. Aliment. Pharmacol. Ther. 2018, 47, 868–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.H.; Kim, N.; Kwon, J.W.; Kim, S.E.; Baik, G.H.; Lee, J.Y.; Park, K.S.; Shin, J.E.; Song, H.J.; Myung, D.S.; et al. Trends in the seroprevalence of Helicobacter pylori infection and its putative eradication rate over 18 years in Korea: A cross-sectional nationwide multicenter study. PLoS ONE 2018, 13, e0204762. [Google Scholar] [CrossRef] [Green Version]
- Duchen, M.R. Mitochondria and calcium: From cell signalling to cell death. J. Physiol. 2000, 529 Pt 1, 57–68. [Google Scholar] [CrossRef]
- Susin, S.A.; Lorenzo, H.K.; Zamzami, N.; Marzo, I.; Snow, B.E.; Brothers, G.M.; Mangion, J.; Jacotot, E.; Costantini, P.; Loeffler, M.; et al. Molecular characterization of mitochondrial apoptosis-inducing factor. Nature 1999, 397, 441–446. [Google Scholar] [CrossRef]
- Huang, X.W.; Luo, R.H.; Zhao, Q.; Shen, Z.Z.; Huang, L.L.; An, X.Y.; Zhao, L.J.; Wang, J.; Huang, Y.Z. Helicobacter pylori induces mitochondrial DNA mutation and reactive oxygen species level in AGS cells. Int. J. Med. Sci. 2011, 8, 56–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, A.M.; Figueiredo, C.; Touati, E.; Maximo, V.; Sousa, S.; Michel, V.; Carneiro, F.; Nielsen, F.C.; Seruca, R.; Rasmussen, L.J. Helicobacter pylori infection induces genetic instability of nuclear and mitochondrial DNA in gastric cells. Clin. Cancer Res. 2009, 15, 2995–3002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zambon, C.F.; Basso, D.; Navaglia, F.; Belluco, C.; Falda, A.; Fogar, P.; Greco, E.; Gallo, N.; Rugge, M.; Di Mario, F.; et al. Pro- and anti-inflammatory cytokines gene polymorphisms and Helicobacter pylori infection: Interactions influence outcome. Cytokine 2005, 29, 141–152. [Google Scholar] [CrossRef]
- Achyut, B.R.; Tripathi, P.; Ghoshal, U.C.; Moorchung, N.; Mittal, B. Interleukin-10 (-819 C/T) and tumor necrosis factor-alpha (-308 G/A) gene variants influence gastritis and lymphoid follicle development. Dig. Dis. Sci. 2008, 53, 622–629. [Google Scholar] [CrossRef]
- Kusters, J.G.; van Vliet, A.H.; Kuipers, E.J. Pathogenesis of Helicobacter pylori infection. Clin. Microbiol. Rev. 2006, 19, 449–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marginean, M.O.; Marginean, C.O.; Melit, L.E.; Voidazan, S.; Moldovan, V.; Banescu, C. The impact of host’s genetic susceptibility on Helicobacter pylori infection in children. Medicine 2017, 96, e7612. [Google Scholar] [CrossRef]
- Mitchell, S.L.; Goodloe, R.; Brown-Gentry, K.; Pendergrass, S.A.; Murdock, D.G.; Crawford, D.C. Characterization of mitochondrial haplogroups in a large population-based sample from the United States. Hum. Genet. 2014, 133, 861–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Walt, J.M.; Dementieva, Y.A.; Martin, E.R.; Scott, W.K.; Nicodemus, K.K.; Kroner, C.C.; Welsh-Bohmer, K.A.; Saunders, A.M.; Roses, A.D.; Small, G.W.; et al. Analysis of European mitochondrial haplogroups with Alzheimer disease risk. Neurosci. Lett. 2004, 365, 28–32. [Google Scholar] [CrossRef]
- Kenney, M.C.; Hertzog, D.; Chak, G.; Atilano, S.R.; Khatibi, N.; Soe, K.; Nobe, A.; Yang, E.; Chwa, M.; Zhu, F.; et al. Mitochondrial DNA haplogroups confer differences in risk for age-related macular degeneration: A case control study. BMC Med. Genet. 2013, 14, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinnery, P.F.; Mowbray, C.; Patel, S.K.; Elson, J.L.; Sampson, M.; Hitman, G.A.; McCarthy, M.I.; Hattersley, A.T.; Walker, M. Mitochondrial DNA haplogroups and type 2 diabetes: A study of 897 cases and 1010 controls. J. Med. Genet. 2007, 44, e80. [Google Scholar] [CrossRef]
- Fang, H.; Hu, N.; Zhao, Q.; Wang, B.; Zhou, H.; Fu, Q.; Shen, L.; Chen, X.; Shen, F.; Lyu, J. mtDNA Haplogroup N9a Increases the Risk of Type 2 Diabetes by Altering Mitochondrial Function and Intracellular Mitochondrial Signals. Diabetes 2018, 67, 1441–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita-Daitoku, R.; Ogura, Y.; Kiga, K.; Maruyama, F.; Kondo, T.; Nakagawa, I.; Hayashi, T.; Mimuro, H. Complete Genome Sequence of Helicobacter pylori Strain ATCC 43504, a Type Strain That Can Infect Gerbils. Microbiol. Resour. Announc. 2020, 9. [Google Scholar] [CrossRef]
- Kang, E.; Wang, X.; Tippner-Hedges, R.; Ma, H.; Folmes, C.D.; Gutierrez, N.M.; Lee, Y.; Van Dyken, C.; Ahmed, R.; Li, Y.; et al. Age-Related Accumulation of Somatic Mitochondrial DNA Mutations in Adult-Derived Human iPSCs. Cell Stem Cell 2016, 18, 625–636. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.J.; Tyler-Smith, C.; Kim, W. The peopling of Korea revealed by analyses of mitochondrial DNA and Y-chromosomal markers. PLoS ONE 2009, 4, e4210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonciarz, W.; Krupa, A.; Hinc, K.; Obuchowski, M.; Moran, A.P.; Gajewski, A.; Chmiela, M. The effect of Helicobacter pylori infection and different H. pylori components on the proliferation and apoptosis of gastric epithelial cells and fibroblasts. PLoS ONE 2019, 14, e0220636. [Google Scholar]
- He, Y.; Wang, C.; Zhang, X.; Lu, X.; Xing, J.; Lv, J.; Guo, M.; Huo, X.; Liu, X.; Lu, J.; et al. Sustained Exposure to Helicobacter pylori Lysate Inhibits Apoptosis and Autophagy of Gastric Epithelial Cells. Front. Oncol. 2020, 10, 581364. [Google Scholar] [CrossRef] [PubMed]
- Yakoob, J.; Abbas, Z.; Jafri, W.; Khan, R.; Salim, S.A.; Awan, S.; Abid, S.; Hamid, S.; Ahmad, Z. Helicobacter pylori outer membrane protein and virulence marker differences in expatriate patients. Epidemiol. Infect. 2016, 144, 2200–2208. [Google Scholar] [CrossRef] [Green Version]
- Matsunaga, S.; Nishiumi, S.; Tagawa, R.; Yoshida, M. Alterations in metabolic pathways in gastric epithelial cells infected with Helicobacter pylori. Microb. Pathog. 2018, 124, 122–129. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Lee, P.; Galbiati, F.; Kitsis, R.N.; Lisanti, M.P. Caveolin-1 expression sensitizes fibroblastic and epithelial cells to apoptotic stimulation. Am. J. Physiol. Cell Physiol. 2001, 280, 823. [Google Scholar] [CrossRef]
- Kohda, K.; Tanaka, K.; Aiba, Y.; Yasuda, M.; Miwa, T.; Koga, Y. Role of apoptosis induced by Helicobacter pylori infection in the development of duodenal ulcer. Gut 1999, 44, 456–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, T.; Nakajima, N.; Kuwayama, H.; Ito, Y.; Iwasaki, A.; Arakawa, Y. Gastric epithelial cell proliferation and apoptosis in Helicobacter pylori-infected mice. Aliment. Pharmacol. Ther. 2000, 14 (Suppl. 1), 68–73. [Google Scholar] [CrossRef] [PubMed]
- Domek, M.J.; Netzer, P.; Prins, B.; Nguyen, T.; Liang, D.; Wyle, F.A.; Warner, A. Helicobacter pylori induces apoptosis in human epithelial gastric cells by stress activated protein kinase pathway. Helicobacter 2001, 6, 110–115. [Google Scholar] [CrossRef]
- Assis, S.; Marques, C.R.; Silva, T.M.; Costa, R.S.; Alcantara-Neves, N.M.; Barreto, M.L.; Barnes, K.C.; Figueiredo, C.A. IL10 single nucleotide polymorphisms are related to upregulation of constitutive IL-10 production and susceptibility to Helicobacter pylori infection. Helicobacter 2014, 19, 168–173. [Google Scholar] [CrossRef]
- Shirin, H.; Sordillo, E.M.; Oh, S.H.; Yamamoto, H.; Delohery, T.; Weinstein, I.B.; Moss, S.F. Helicobacter pylori inhibits the G1 to S transition in AGS gastric epithelial cells. Cancer Res. 1999, 59, 2277–2281. [Google Scholar]
- Mookerjee, S.A.; Brand, M.D. Measurement and Analysis of Extracellular Acid Production to Determine Glycolytic Rate. J. Vis. Exp. 2015, e53464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McInnes, J. Mitochondrial-associated metabolic disorders: Foundations, pathologies and recent progress. Nutr. Metab. 2013, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, N.; Tay, A.C.Y.; Marshall, B.J.; Jain, U. Helicobacter pylori VacA, a distinct toxin exerts diverse functionalities in numerous cells: An overview. Helicobacter 2018, 24, e12544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moss, S.F.; Calam, J.; Agarwal, B.; Wang, S.; Holt, P.R. Induction of gastric epithelial apoptosis by Helicobacter pylori. Gut 1996, 38, 498–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handa, O.; Naito, Y.; Yoshikawa, T. Redox biology and gastric carcinogenesis: The role of Helicobacter pylori. Redox Rep. 2011, 16, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.K. Mitochondria damage checkpoint in apoptosis and genome stability. FEMS Yeast Res. 2004, 5, 127–132. [Google Scholar] [CrossRef] [Green Version]
- Capparelli, R.; Iannelli, D. Genetics of Host Protection against Helicobacter pylori Infections. Int. J. Mol. Sci. 2021, 22, 3192. [Google Scholar] [CrossRef]
- Kim, N.; Choi, W.R.; Song, C.H.; Sheen, D.H.; Yang, S.S.; Lee, J.Y.; Han, Y.J.; Lim, S.H.; Lee, K.H.; Choi, S.E. Difference in the distribution pattern of Helicobacter pylori and grade of gastritis in the antrum and in the body between duodenal ulcer and benign gastric ulcer patients. Korean J. Intern. Med. 2000, 15, 32–36. [Google Scholar]
- Naumann, M.; Sokolova, O.; Tegtmeyer, N.; Backert, S. Helicobacter pylori: A Paradigm Pathogen for Subverting Host Cell Signal Transmission. Trends Microbiol. 2017, 25, 316–328. [Google Scholar] [CrossRef] [PubMed]
- De Bont, R.; van Larebeke, N. Endogenous DNA damage in humans: A review of quantitative data. Mutagenesis 2004, 19, 169–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatre, L.; Fernandes, J.; Michel, V.; Fiette, L.; Ave, P.; Arena, G.; Jain, U.; Haas, R.; Wang, T.C.; Ricchetti, M.; et al. Helicobacter pylori targets mitochondrial import and components of mitochondrial DNA replication machinery through an alternative VacA-dependent and a VacA-independent mechanisms. Sci. Rep. 2017, 7, 15901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefano, G.B.; Bjenning, C.; Wang, F.; Wang, N.; Kream, R.M. Mitochondrial Heteroplasmy. Adv. Exp. Med. Biol. 2017, 982, 577–594. [Google Scholar] [PubMed]
- Locke, A.E.; Kahali, B.; Berndt, S.I.; Justice, A.E.; Pers, T.H.; Day, F.R.; Powell, C.; Vedantam, S.; Buchkovich, M.L.; Yang, J.; et al. Genetic studies of body mass index yield new insights for obesity biology. Nature 2015, 518, 197–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, A.V. Evolution, revolution and heresy in the genetics of infectious disease susceptibility. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 840–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H. pylori-Positive Subjects | H. pylori-Negative Subjects | ||||||
|---|---|---|---|---|---|---|---|
| Subject No. | Sex | Age | Mitochondrial Haplotype | Subject No. | Sex | Age | Mitochondrial Haplotype |
| 1 | M | 45 | M8a | 1 | M | 48 | G1a |
| 2 | M | 45 | N9a | 2 | M | 54 | F1a |
| 3 | M | 46 | B5b | 3 | M | 54 | D4i |
| 4 | M | 49 | D4i | 4 | M | 56 | M7b |
| 5 | F | 50 | M7c | 5 | F | 57 | D4a |
| 6 | M | 50 | D4b | 6 | M | 58 | D4g |
| 7 | F | 51 | N9a | 7 | M | 60 | D4b |
| 8 | M | 52 | G3a | 8 | M | 60 | D4j |
| 9 | M | 52 | B4c | 9 | M | 61 | C |
| 10 | M | 53 | F1b | 10 | F | 67 | B5b |
| 11 | F | 54 | G3a | 11 | M | 67 | G2a |
| 12 | M | 57 | D4a | 12 | M | 68 | M7c |
| 13 | M | 58 | B5a | 13 | M | 70 | D4f |
| 14 | F | 59 | D4j | 14 | F | 72 | B5a |
| 15 | F | 60 | A | 15 | F | 73 | M10a |
| 16 | M | 61 | M7b | ||||
| 17 | M | 64 | N9a | ||||
| 18 | M | 64 | M7b | ||||
| 19 | M | 67 | B4c | ||||
| 20 | F | 67 | M7b | ||||
| 21 | F | 67 | B5b | ||||
| 22 | M | 68 | M9a | ||||
| 23 | F | 69 | D4h | ||||
| 24 | M | 72 | B4a | ||||
| 25 | F | 73 | D5a | ||||
| 26 | F | 73 | F1a | ||||
| 27 | M | 78 | M10a | ||||
| 28 | M | 78 | B4b | ||||
| Group | No. of H. pylori-Positive (Mean Age ± SEM) | No. of H. pylori-Negative (Mean Age ± SEM) | |
|---|---|---|---|
| Age | <60 | 14 (52 ± 1) | 6 (55 ± 1) |
| 60≤ | 14 (69 ± 1) | 9 (66 ± 1) | |
| Sex | Male | 18 (59 ± 3) | 11 (60 ± 2) |
| Female | 10 (62 ± 3) | 4 (67 ± 4) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.; Lee, S.-M.; Choi, J.; Kang, S.; So, S.; Kim, D.; Ahn, J.-Y.; Jung, H.-Y.; Jeong, J.-Y.; Kang, E. Mitochondrial DNA Haplogroup Related to the Prevalence of Helicobacter pylori. Cells 2021, 10, 2482. https://doi.org/10.3390/cells10092482
Lee Y, Lee S-M, Choi J, Kang S, So S, Kim D, Ahn J-Y, Jung H-Y, Jeong J-Y, Kang E. Mitochondrial DNA Haplogroup Related to the Prevalence of Helicobacter pylori. Cells. 2021; 10(9):2482. https://doi.org/10.3390/cells10092482
Chicago/Turabian StyleLee, Yeonmi, Sun-Mi Lee, Jiwan Choi, Seoon Kang, Seongjun So, Deokhoon Kim, Ji-Yong Ahn, Hwoon-Yong Jung, Jin-Yong Jeong, and Eunju Kang. 2021. "Mitochondrial DNA Haplogroup Related to the Prevalence of Helicobacter pylori" Cells 10, no. 9: 2482. https://doi.org/10.3390/cells10092482