
The Role of Stem Cells and Their Derived Extracellular Vesicles in Restoring Female and Male Fertility

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
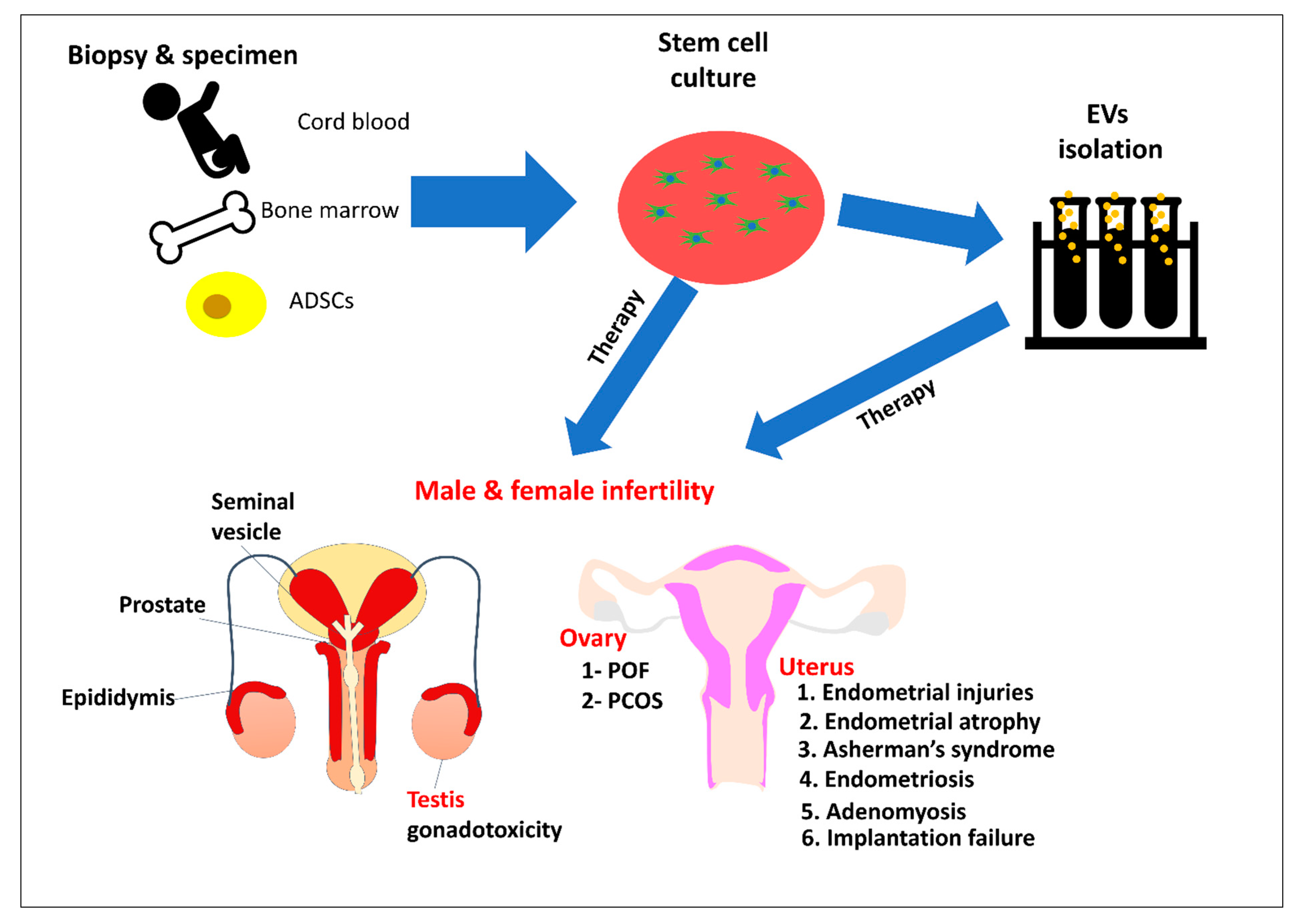
2. Stem Cell for Therapy
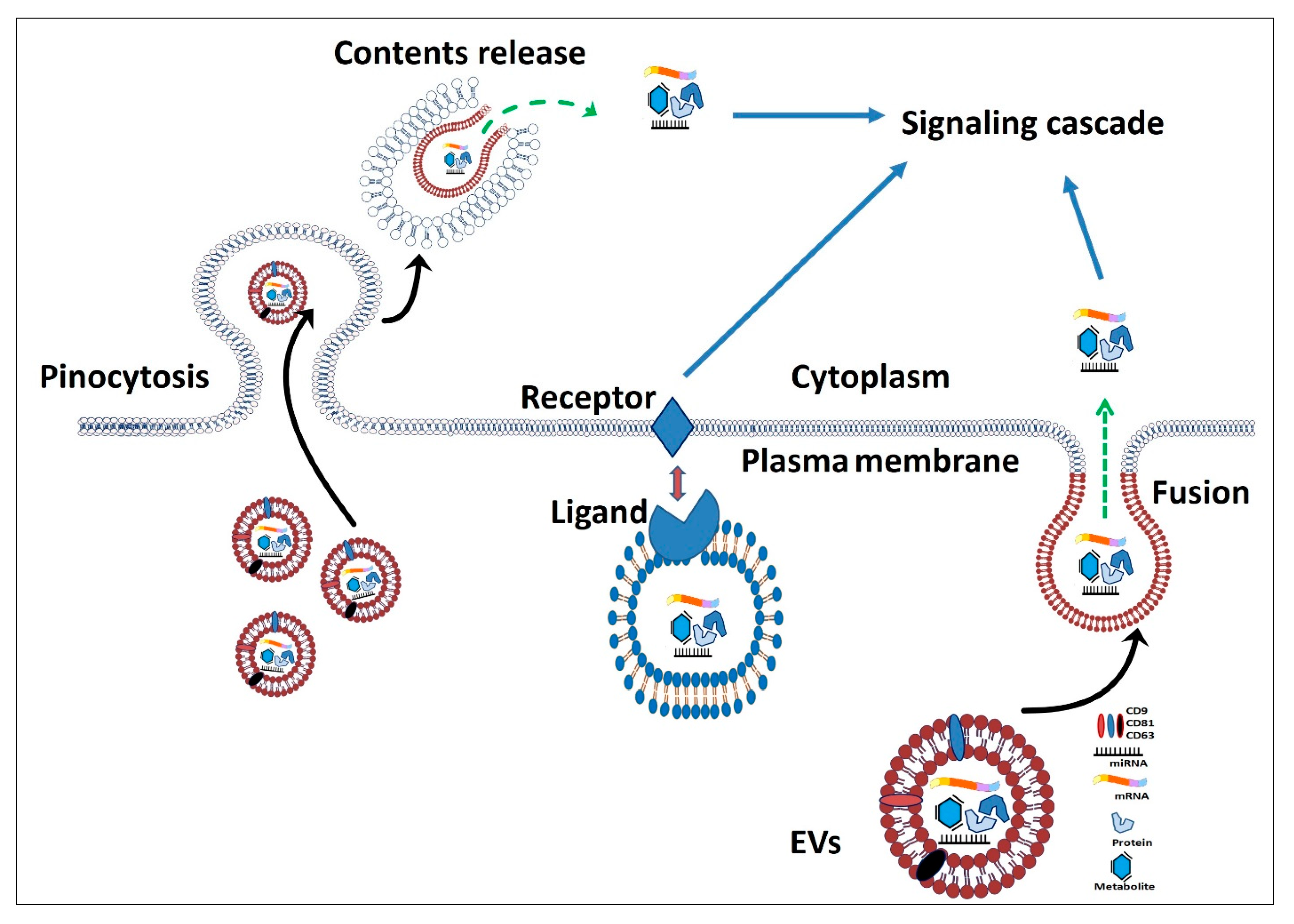
3. Extracellular Vesicles as Alternative Therapeutics
4. Stem Cells and Female Reproductive Issues
4.1. Premature Ovarian Failure
4.2. Polycystic Ovarian Syndrome
4.3. Endometrial Injuries
4.4. Endometrial Atrophy
4.5. Repeated Implantation Failure and Recurrent Miscarriage
4.6. Asherman’s Syndrome
4.7. Endometriosis
4.8. Adenomyosis
5. Stem Cells and Male Reproductive Issues
5.1. Hypogonadism
5.1.1. Varicocele
5.1.2. Testicular Trauma
5.1.3. Testicular Torsion
5.1.4. Klinefelter’s Syndrome
5.2. Epididymo-Orchitis
5.3. Gonadotoxicity
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fazeli, Z.; Abedindo, A.; Omrani, M.D.; Ghaderian, S.M.H. Mesenchymal stem cells (MSCs) therapy for recovery of fertility: A systematic review. Stem Cell Rev. Rep. 2018, 14, 1–12. [Google Scholar] [CrossRef]
- Miyamoto, T.; Tsujimura, A.; Miyagawa, Y.; Koh, E.; Namiki, M.; Sengoku, K. Male infertility and its causes in human. Adv. Urol. 2011, 2012, 384520. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, T.; Minase, G.; Okabe, K.; Ueda, H.; Sengoku, K. Male infertility and its genetic causes. J. Obstet. Gynaecol. Res. 2015, 41, 1501–1505. [Google Scholar] [CrossRef]
- Giudice, M.G.; Del Vento, F.; Wyns, C. Male fertility preservation in DSD, XXY, pre-gonadotoxic treatments–update, methods, ethical issues, current outcomes, future directions. Best Pract. Res. Clin. Endocrinol. Metab. 2019, 33, 101261. [Google Scholar] [CrossRef] [PubMed]
- Gidoni, Y.; Holzer, H.; Tulandi, T.; Tan, S.L. Fertility preservation in patients with non-oncological conditions. Reprod. Biomed. Online 2008, 16, 792–800. [Google Scholar] [CrossRef]
- Anwar, S.; Anwar, A. Infertility: A review on causes, treatment and management. Women’s Health Gynecol. 2016, 5, 2. [Google Scholar]
- Semet, M.; Paci, M.; Saïas-Magnan, J.; Metzler-Guillemain, C.; Boissier, R.; Lejeune, H.; Perrin, J. The impact of drugs on male fertility: A review. Andrology 2017, 5, 640–663. [Google Scholar] [CrossRef]
- Holmberg, L.; Iversen, O.-E.; Rudenstam, C.M.; Hammar, M.; Kumpulainen, E.; Jaskiewicz, J.; Jassem, J.; Dobaczewska, D.; Fjosne, H.E.; Peralta, O. Increased risk of recurrence after hormone replacement therapy in breast cancer survivors. J. Natl. Cancer Inst. 2008, 100, 475–482. [Google Scholar] [CrossRef] [Green Version]
- Vermeulen, R.; Korse, C.; Kenter, G.; Brood-van Zanten, M.; Beurden, M.v. Safety of hormone replacement therapy following risk-reducing salpingo-oophorectomy: Systematic review of literature and guidelines. Climacteric 2019, 22, 352–360. [Google Scholar] [CrossRef]
- Schlegel, P. Evaluation of male infertility. Minerva Ginecol. 2009, 61, 261–283. [Google Scholar]
- Gómez, R.; Soares, S.R.; Busso, C.; Garcia-Velasco, J.A.; Simon, C.; Pellicer, A. Physiology and pathology of ovarian hyperstimulation syndrome. Semin. Reprod. Med. 2010, 28, 448–457. [Google Scholar] [CrossRef]
- Prakash, A.; Mathur, R. Ovarian hyperstimulation syndrome. Obstet. Gynaecol. 2013, 15, 31–35. [Google Scholar] [CrossRef]
- Van Voorhis, B.J.; Ryan, G.L. Ethical obligation for restricting the number of embryos transferred to women: Combating the multiple-birth epidemic from in vitro fertilization. Semin. Reprod. Med. 2010, 28, 287–294. [Google Scholar] [CrossRef]
- Garcia Cruz, D.M.; Gomes, M.; Reis, R.L.; Moratal, D.; Salmerón-Sánchez, M.; Gomez Ribelles, J.L.; Mano, J.F. Differentiation of mesenchymal stem cells in chitosan scaffolds with double micro and macroporosity. J. Biomed. Mater. Res. Part A 2010, 95, 1182–1193. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.S.; Ryu, H.-S.; Jung, S.-E.; Shin, B.-J.; Won, J.-H.; Um, T.G.; Oh, H.; Kim, S.-H.; Ryu, B.-Y. Expression profile of spermatogenesis associated genes in male germ cells during postnatal development in mice. J. Anim. Reprod. Biotechnol. 2020, 35, 289–296. [Google Scholar] [CrossRef]
- Watt, F.M.; Hogan, B.L. Out of Eden: Stem cells and their niches. Science 2000, 287, 1427–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alison, M.R.; Poulsom, R.; Forbes, S.; Wright, N.A. An introduction to stem cells. J. Pathol. 2002, 197, 419–423. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Taylor, H.S. Stem cells and female reproduction. Reprod. Sci. 2009, 16, 126–139. [Google Scholar]
- Méndez-Ferrer, S.; Michurina, T.V.; Ferraro, F.; Mazloom, A.R.; MacArthur, B.D.; Lira, S.A.; Scadden, D.T.; Ma’ayan, A.; Enikolopov, G.N.; Frenette, P.S. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 2010, 466, 829–834. [Google Scholar] [CrossRef]
- Easley, C.A., IV; Simerly, C.R.; Schatten, G. Stem cell therapeutic possibilities: Future therapeutic options for male-factor and female-factor infertility? Reprod. Biomed. Online 2013, 27, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Brinster, R.L. Male germline stem cells: From mice to men. Science 2007, 316, 404–405. [Google Scholar] [CrossRef] [Green Version]
- Oatley, J.M.; Brinster, R.L. Spermatogonial stem cells. Methods Enzymol. 2006, 419, 259–282. [Google Scholar]
- Hermann, B.P.; Sukhwani, M.; Winkler, F.; Pascarella, J.N.; Peters, K.A.; Sheng, Y.; Valli, H.; Rodriguez, M.; Ezzelarab, M.; Dargo, G. Spermatogonial stem cell transplantation into rhesus testes regenerates spermatogenesis producing functional sperm. Cell Stem Cell 2012, 11, 715–726. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Liu, X.; Shi, Y.; Ocansey, D.K.W.; Hu, Y.; Li, X.; Zhang, C.; Xu, W.; Qian, H. Therapeutic Advances of Stem Cell-Derived Extracellular Vesicles in Regenerative Medicine. Cells 2020, 9, 707. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-C.; Lai, L.-C. The potential roles of stem cell-derived extracellular vesicles as a therapeutic tool. Ann. Transl. Med. 2019, 7, 693. [Google Scholar] [CrossRef]
- Lo, B.; Parham, L. Ethical Issues in Stem Cell Research. Endocr. Rev. 2009, 30, 204–213. [Google Scholar] [CrossRef]
- King, N.M.P.; Perrin, J. Ethical issues in stem cell research and therapy. Stem Cell Res. Ther. 2014, 5, 85. [Google Scholar] [CrossRef] [Green Version]
- Zakirova, E.Y.; Aimaletdinov, A.M.; Malanyeva, A.G.; Rutland, C.S.; Rizvanov, A.A. Extracellular Vesicles: New Perspectives of Regenerative and Reproductive Veterinary Medicine. Front. Vet. Sci. 2020, 7, 931. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Bedina Zavec, A.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, K.A.; Xiang, J. Mechanisms of cellular communication through intercellular protein transfer. J. Cell. Mol. Med. 2011, 15, 1458–1473. [Google Scholar] [CrossRef]
- Alzahrani, F.A.; Saadeldin, I.M. Role of Exosomes in Biological Communication Systems; Springer Nature: Singapore, 2021. [Google Scholar] [CrossRef]
- Al-shubaily, F.A.; Al-Zahrani, M.H. Characterization and Fine Structure of Exosomes. In Role of Exosomes in Biological Communication Systems; Springer: Singapore, 2021; pp. 27–75. [Google Scholar] [CrossRef]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Théry, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Tkach, M.; Kowal, J.; Zucchetti, A.E.; Enserink, L.; Jouve, M.; Lankar, D.; Saitakis, M.; Martin-Jaular, L.; Théry, C. Qualitative differences in T-cell activation by dendritic cell-derived extracellular vesicle subtypes. EMBO J. 2017, 36, 3012–3028. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.Y. Mesenchymal stem cells for restoration of ovarian function. Clin. Exp. Reprod. Med. 2019, 46, 1. [Google Scholar] [CrossRef] [Green Version]
- Rungsiwiwut, R.; Virutamasen, P.; Pruksananonda, K. Mesenchymal stem cells for restoring endometrial function: An infertility perspective. Reprod. Med. Biol. 2021, 20, 13–19. [Google Scholar] [CrossRef]
- Nelson, L.M.; Anasti, J.N.; Flak, M.R. Premature ovarian failure. In Reproductive Endocrinology, Surgery, and Technology; Adashi, E.Y., Rock, J.A., Rosenwaks, Z., Eds.; Lippincott-Raven: Philadelphia, PA, USA, 1996; pp. 1394–1410. [Google Scholar]
- Luisi, S.; Orlandini, C.; Regini, C.; Pizzo, A.; Vellucci, F.; Petraglia, F. Premature ovarian insufficiency: From pathogenesis to clinical management. J. Endocrinol. Investig. 2015, 38, 597–603. [Google Scholar] [CrossRef]
- McGuire, M.M.; Bowden, W.; Engel, N.J.; Ahn, H.W.; Kovanci, E.; Rajkovic, A. Genomic analysis using high-resolution single-nucleotide polymorphism arrays reveals novel microdeletions associated with premature ovarian failure. Fertil. Steril. 2011, 95, 1595–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, M.; Cummings, L.; Chada, K. Germ cell deficient (gcd) mouse as a model of premature ovarian failure. Biol. Reprod. 1993, 49, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.; Yu, Y.; Huang, H. An update on primary ovarian insufficiency. Sci. China Life Sci. 2012, 55, 677–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, G.-Y.; Liu, I.-H.; Cheng, C.-C.; Chang, C.-C.; Lee, Y.-H.; Cheng, W.T.-K.; Wu, S.-C. Amniotic fluid stem cells prevent follicle atresia and rescue fertility of mice with premature ovarian failure induced by chemotherapy. PLoS ONE 2014, 9, e106538. [Google Scholar] [CrossRef]
- Bao, R.; Xu, P.; Wang, Y.; Wang, J.; Xiao, L.; Li, G.; Zhang, C. Bone marrow derived mesenchymal stem cells transplantation rescues premature ovarian insufficiency induced by chemotherapy. Gynecol. Endocrinol. 2018, 34, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Zhong, Y.; Qian, C.; Zou, Q.; Ou, J.; Shi, Y.; Gao, L.; Wang, G.; Liu, Z.; Li, H. Human umbilical cord mesenchymal stem cells therapy in cyclophosphamide-induced premature ovarian failure rat model. BioMed. Res. Int. 2016, 2016, 2517514. [Google Scholar] [CrossRef] [Green Version]
- Herraiz, S.; Pellicer, N.; Romeu, M.; Pellicer, A. Treatment potential of bone marrow-derived stem cells in women with diminished ovarian reserves and premature ovarian failure. Curr. Opin. Obstet. Gynecol. 2019, 31, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Lin, H. Asymmetric germ cell division and oocyte determination during Drosophila oogenesis. Int. Rev. Cytol. 2001, 203, 93–138. [Google Scholar]
- Liu, J.; Zhang, H.; Zhang, Y.; Li, N.; Wen, Y.; Cao, F.; Ai, H.; Xue, X. Homing and restorative effects of bone marrow-derived mesenchymal stem cells on cisplatin injured ovaries in rats. Mol. Cells 2014, 37, 865. [Google Scholar] [CrossRef] [Green Version]
- Ozcan Cenksoy, P.; Oktem, M.; Erdem, O.; Karakaya, C.; Cenksoy, C.; Erdem, A.; Guner, H.; Karabacak, O. A potential novel treatment strategy: Inhibition of angiogenesis and inflammation by resveratrol for regression of endometriosis in an experimental rat model. Gynecol. Endocrinol. 2015, 31, 219–224. [Google Scholar] [CrossRef]
- Ding, C.; Zou, Q.; Wang, F.; Wu, H.; Wang, W.; Li, H.; Huang, B. HGF and BFGF secretion by human adipose-derived stem cells improves ovarian function during natural aging via activation of the SIRT1/FOXO1 signaling pathway. Cell. Physiol. Biochem. 2018, 45, 1316–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terraciano, P.; Garcez, T.; Ayres, L.; Durli, I.; Baggio, M.; Kuhl, C.P.; Laurino, C.; Passos, E.; Paz, A.H.; Cirne-Lima, E. Cell therapy for chemically induced ovarian failure in mice. Stem Cells Int. 2014, 2014, 720753. [Google Scholar] [CrossRef]
- Sun, M.; Wang, S.; Li, Y.; Yu, L.; Gu, F.; Wang, C.; Yao, Y. Adipose-derived stem cells improved mouse ovary function after chemotherapy-induced ovary failure. Stem Cell Res. Ther. 2013, 4, 80. [Google Scholar] [CrossRef] [Green Version]
- Abd-Allah, S.H.; Shalaby, S.M.; Pasha, H.F.; Amal, S.; Raafat, N.; Shabrawy, S.M.; Awad, H.A.; Amer, M.G.; Gharib, M.A.; El Gendy, E.A. Mechanistic action of mesenchymal stem cell injection in the treatment of chemically induced ovarian failure in rabbits. Cytotherapy 2013, 15, 64–75. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.; Bagley, J.; Skaznik-Wikiel, M.; Lee, H.-J.; Adams, G.B.; Niikura, Y.; Tschudy, K.S.; Tilly, J.C.; Cortes, M.L.; Forkert, R. Oocyte generation in adult mammalian ovaries by putative germ cells in bone marrow and peripheral blood. Cell 2005, 122, 303–315. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Huang, Y.; Zhang, J.; Qin, W.; Chi, H.; Chen, J.; Yu, Z.; Chen, C. Transplantation of human menstrual blood stem cells to treat premature ovarian failure in mouse model. Stem Cells Dev. 2014, 23, 1548–1557. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, Y.; Yang, T.; Li, J.; Yang, X. Study of the reparative effects of menstrual-derived stem cells on premature ovarian failure in mice. Stem Cell Res. Ther. 2017, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Elfayomy, A.K.; Almasry, S.M.; El-Tarhouny, S.A.; Eldomiaty, M.A. Human umbilical cord blood-mesenchymal stem cells transplantation renovates the ovarian surface epithelium in a rat model of premature ovarian failure: Possible direct and indirect effects. Tissue Cell 2016, 48, 370–382. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Yan, G.; Wang, B.; Xu, L.; Gu, Y.; Ru, T.; Cui, X.; Lei, L.; Liu, J.; Sheng, X. Transplantation of UC-MSCs on collagen scaffold activates follicles in dormant ovaries of POF patients with long history of infertility. Sci. China Life Sci. 2018, 61, 1554–1565. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yu, L.; Sun, M.; Mu, S.; Wang, C.; Wang, D.; Yao, Y. The therapeutic potential of umbilical cord mesenchymal stem cells in mice premature ovarian failure. Biomed Res. Int. 2013, 2013, 690491. [Google Scholar] [CrossRef]
- Lai, D.; Wang, F.; Dong, Z.; Zhang, Q. Skin-derived mesenchymal stem cells help restore function to ovaries in a premature ovarian failure mouse model. PLoS ONE 2014, 9, e98749. [Google Scholar] [CrossRef]
- Kilic, S.; Pinarli, F.; Ozogul, C.; Tasdemir, N.; Naz Sarac, G.; Delibasi, T. Protection from cyclophosphamide-induced ovarian damage with bone marrow-derived mesenchymal stem cells during puberty. Gynecol. Endocrinol. 2014, 30, 135–140. [Google Scholar] [CrossRef]
- Cho, J.; Kim, T.-H.; Seok, J.; Jun, J.H.; Park, H.; Kweon, M.; Lim, J.-Y.; Kim, G.J. Vascular remodeling by placenta-derived mesenchymal stem cells restores ovarian function in ovariectomized rat model via the VEGF pathway. Lab. Investig. 2021, 101, 304–317. [Google Scholar] [CrossRef] [PubMed]
- Seok, J.; Park, H.; Choi, J.H.; Lim, J.-Y.; Kim, K.G.; Kim, G.J. Placenta-derived mesenchymal stem cells restore the ovary function in an ovariectomized rat model via an antioxidant effect. Antioxidants 2020, 9, 591. [Google Scholar] [CrossRef]
- Fouad, H.; Sabry, D.; Elsetohy, K.; Fathy, N. Therapeutic efficacy of amniotic membrane stem cells and adipose tissue stem cells in rats with chemically induced ovarian failure. J. Adv. Res. 2016, 7, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Du, X.; Wang, C.; Zhang, J.; Liu, C.; Li, Y.; Jiang, H. Therapeutic effects of human umbilical cord mesenchymal stem cell-derived microvesicles on premature ovarian insufficiency in mice. Stem Cell Res. Ther. 2019, 10, 250. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Li, D.; Song, K.; Wei, J.; Yao, S.; Li, Z.; Su, X.; Ju, X.; Chao, L.; Deng, X. Exosomes derived from human umbilical cord mesenchymal stem cells protect against cisplatin-induced ovarian granulosa cell stress and apoptosis in vitro. Sci. Rep. 2017, 7, 2552. [Google Scholar] [CrossRef] [Green Version]
- Hong, L.; Yan, L.; Xin, Z.; Hao, J.; Liu, W.; Wang, S.; Liao, S.; Wang, H.; Yang, X. Protective effects of human umbilical cord mesenchymal stem cell-derived conditioned medium on ovarian damage. J. Mol. Cell Biol. 2020, 12, 372–385. [Google Scholar] [CrossRef]
- Zhang, J.; Yin, H.; Jiang, H.; Du, X.; Yang, Z. The protective effects of human umbilical cord mesenchymal stem cell-derived extracellular vesicles on cisplatin-damaged granulosa cells. Taiwan. J. Obstet. Gynecol. 2020, 59, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Bu, S.; Sun, J.; Xu, M.; Yao, X.; He, K.; Lai, D. Paracrine effects of human amniotic epithelial cells protect against chemotherapy-induced ovarian damage. Stem Cell Res. Ther. 2017, 8, 270. [Google Scholar] [CrossRef] [Green Version]
- Sun, B.; Ma, Y.; Wang, F.; Hu, L.; Sun, Y. miR-644-5p carried by bone mesenchymal stem cell-derived exosomes targets regulation of p53 to inhibit ovarian granulosa cell apoptosis. Stem Cell Res. Ther. 2019, 10, 360. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Lin, L.; Sha, C.; Li, T.; Zhao, D.; Wei, H.; Chen, Q.; Liu, Y.; Chen, X.; Xu, W. Bone marrow mesenchymal stem cell-derived exosomal miR-144-5p improves rat ovarian function after chemotherapy-induced ovarian failure by targeting PTEN. Lab. Investig. 2020, 100, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Sun, J.; Huang, Y.; Bu, S.; Guo, Y.; Gu, T.; Li, B.; Wang, C.; Lai, D. Human amniotic epithelial cell-derived exosomes restore ovarian function by transferring microRNAs against apoptosis. Mol. Ther.-Nucleic Acids 2019, 16, 407–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, C.; Qian, C.; Hou, S.; Lu, J.; Zou, Q.; Li, H.; Huang, B. Exosomal miRNA-320a is released from hAMSCs and regulates SIRT4 to prevent reactive oxygen species generation in POI. Mol. Ther.-Nucleic Acids 2020, 21, 37–50. [Google Scholar] [CrossRef]
- Shi, Z.; Wang, Q.; Zhang, Y.; Jiang, D. Extracellular vesicles produced by bone marrow mesenchymal stem cells attenuate renal fibrosis, in part by inhibiting the RhoA/ROCK pathway, in a UUO rat model. Stem Cell Res. Ther. 2020, 11, 253. [Google Scholar] [CrossRef]
- Cacciottolo, M.; Morgan, T.E.; Saffari, A.A.; Shirmohammadi, F.; Forman, H.J.; Sioutas, C.; Finch, C.E. Traffic-related air pollutants (TRAP-PM) promote neuronal amyloidogenesis through oxidative damage to lipid rafts. Free Radic. Biol. Med. 2020, 147, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Korytkowski, M. Polycystic ovary syndrome: How best to establish the diagnosis. Women’s Health Prim. Care 2000, 3, 55–58. [Google Scholar]
- Franks, S.; White, D.M. Prevalence of and etiological factors in polycystic ovarian syndrome. Ann. N. Y. Acad. Sci. 1993, 687, 112–114. [Google Scholar] [CrossRef] [PubMed]
- Eid, G.M.; Cottam, D.R.; Velcu, L.M.; Mattar, S.G.; Korytkowski, M.T.; Gosman, G.; Hindi, P.; Schauer, P.R. Effective treatment of polycystic ovarian syndrome with Roux-en-Y gastric bypass. Surg. Obes. Relat. Dis. 2005, 1, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Talbott, E.; Guzick, D.; Clerici, A.; Berga, S.; Detre, K.; Weimer, K.; Kuller, L. Coronary heart disease risk factors in women with polycystic ovary syndrome. Arterioscler. Thromb. Vasc. Biol. 1995, 15, 821–826. [Google Scholar] [CrossRef]
- Talbott, E.; Clerici, A.; Berga, S.L.; Kuller, L.; Guzick, D.; Detre, K.; Daniels, T.; Engberg, R.A. Adverse lipid and coronary heart disease risk profiles in young women with polycystic ovary syndrome: Results of a case-control study. J. Clin. Epidemiol. 1998, 51, 415–422. [Google Scholar] [CrossRef]
- Parham, K.J. Patient presentations: Panel discussion: Adenocarcinoma of the endometrium associated with the Stein-Leventhal syndrome. Am. J. Obstet. Gynecol. 1969, 105, 113–115. [Google Scholar] [CrossRef]
- Stein, I.F. Amenorrhea associated with bilateral polycystic ovaries. Am. J. Obs. Gynecol. 1935, 29, 181–191. [Google Scholar] [CrossRef]
- Harwood, K.; Vuguin, P.; DiMartino-Nardi, J. Current approaches to the diagnosis and treatment of polycystic ovarian syndrome in youth. Horm. Res. Paediatr. 2007, 68, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Xie, Q.; Xiong, X.; Xiao, N.; He, K.; Chen, M.; Peng, J.; Su, X.; Mei, H.; Dai, Y.; Wei, D. Mesenchymal stem cells alleviate DHEA-Induced polycystic ovary syndrome (PCOS) by inhibiting inflammation in mice. Stem Cells Int. 2019, 2019, 9782373. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Tao, M.; Wei, M.; Du, S.; Wang, H.; Wang, X. Mesenchymal stem cells derived exosomal miR-323-3p promotes proliferation and inhibits apoptosis of cumulus cells in polycystic ovary syndrome (PCOS). Artif. Cells Nanomed. Biotechnol. 2019, 47, 3804–3813. [Google Scholar] [CrossRef] [PubMed]
- Gargett, C.E. Uterine stem cells: What is the evidence? Hum. Reprod. Update 2007, 13, 87–101. [Google Scholar] [CrossRef]
- Jabbour, H.N.; Kelly, R.W.; Fraser, H.M.; Critchley, H.O. Endocrine regulation of menstruation. Endocr. Rev. 2006, 27, 17–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, W.; Xie, B.; Li, C.; Yan, Y.; Zhang, Y.; Liu, W.; Huang, J.; Chen, D. The clinical applications of endometrial mesenchymal stem cells. Biopreserv. Biobank. 2018, 16, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Cervello, I.; Mas, A.; Gil-Sanchis, C.; Peris, L.; Faus, A.; Saunders, P.T.; Critchley, H.O.; Simon, C. Reconstruction of endometrium from human endometrial side population cell lines. PLoS ONE 2011, 6, e21221. [Google Scholar] [CrossRef] [PubMed]
- Gargett, C.E.; Schwab, K.E.; Zillwood, R.M.; Nguyen, H.P.; Wu, D. Isolation and culture of epithelial progenitors and mesenchymal stem cells from human endometrium. Biol. Reprod. 2009, 80, 1136–1145. [Google Scholar] [CrossRef] [Green Version]
- Gargett, C.E.; Nguyen, H.P.; Ye, L. Endometrial regeneration and endometrial stem/progenitor cells. Rev. Endocr. Metab. Disord. 2012, 13, 235–251. [Google Scholar] [CrossRef]
- Ding, L.; Sun, H.; Su, J.; Lin, N.; Péault, B.; Song, T.; Yang, J.; Dai, J.; Hu, Y. Transplantation of bone marrow mesenchymal stem cells on collagen scaffolds for the functional regeneration of injured rat uterus. Biomaterials 2014, 35, 4888–4900. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, Y.; Guan, C.Y.; Tian, S.; Lv, X.D.; Li, J.-H.; Ma, X.; Xia, H.-F. Therapeutic effect of human umbilical cord-derived mesenchymal stem cells on injured rat endometrium during its chronic phase. Stem Cell Res. Ther. 2018, 9, 36. [Google Scholar] [CrossRef] [Green Version]
- Amable, P.R.; Carias, R.B.V.; Teixeira, M.V.T.; da Cruz Pacheco, Í.; do Amaral, R.J.F.C.; Granjeiro, J.M.; Borojevic, R. Platelet-rich plasma preparation for regenerative medicine: Optimization and quantification of cytokines and growth factors. Stem Cell Res. Ther. 2013, 4, 67. [Google Scholar] [CrossRef] [Green Version]
- Fotouhi, A.; Maleki, A.; Dolati, S.; Aghebati-Maleki, A.; Aghebati-Maleki, L. Platelet rich plasma, stromal vascular fraction and autologous conditioned serum in treatment of knee osteoarthritis. Biomed. Pharmacother. 2018, 104, 652–660. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.W.; Cheng, M.J.; Xu, C.J. Differentiation of mouse bone marrow mesenchymal stem cells into endometrial epithelial cells in vitro. Prog. Obstet. Gynecol. 2010, 19, 257–260. [Google Scholar]
- Yang, X.; Zhang, M.; Zhang, Y.; Li, W.; Yang, B. Mesenchymal stem cells derived from Wharton jelly of the human umbilical cord ameliorate damage to human endometrial stromal cells. Fertil. Steril. 2011, 96, 1029–1036.e1024. [Google Scholar] [CrossRef]
- Koippallil Gopalakrishnan, A.R.; Kishore, U.; Madan, T. Mesenchymal stem cells: A promising tool for targeted gene therapy of endometriosis. Regen. Med. 2017, 12, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Xiao, B.; Zhu, Y.; Huang, J.; Wang, T.; Wang, F.; Sun, S. Exosomal transfer of bone marrow mesenchymal stem cell-derived miR-340 attenuates endometrial fibrosis. Biol. Open 2019, 8, bio039958. [Google Scholar] [CrossRef] [Green Version]
- Liang, L.; Wang, L.; Zhou, S.; Li, J.; Meng, L.; Zhang, H.; Cui, C.; Zhang, C. Exosomes derived from human umbilical cord mesenchymal stem cells repair injured endometrial epithelial cells. J. Assist. Reprod. Genet. 2020, 37, 395–403. [Google Scholar] [CrossRef]
- Wang, J.; Hu, R.; Xing, Q.; Feng, X.; Jiang, X.; Xu, Y.; Wei, Z. Exosomes Derived from Umbilical Cord Mesenchymal Stem Cells Alleviate Mifepristone-Induced Human Endometrial Stromal Cell Injury. Stem Cells Int. 2020, 2020, 6091269. [Google Scholar] [CrossRef] [PubMed]
- Senturk, L.M.; Erel, C.T. Thin endometrium in assisted reproductive technology. Curr. Opin. Obstet. Gynecol. 2008, 20, 221–228. [Google Scholar] [CrossRef]
- Chang, Y.; Li, J.; Chen, Y.; Wei, L.; Yang, X.; Shi, Y.; Liang, X. Autologous platelet-rich plasma promotes endometrial growth and improves pregnancy outcome during in vitro fertilization. Int. J. Clin. Exp. Med. 2015, 8, 1286. [Google Scholar] [PubMed]
- Tandulwadkar, S.R.; Naralkar, M.V.; Surana, A.D.; Selvakarthick, M.; Kharat, A.H. Autologous intrauterine platelet-rich plasma instillation for suboptimal endometrium in frozen embryo transfer cycles: A pilot study. J. Hum. Reprod. Sci. 2017, 10, 208. [Google Scholar] [CrossRef]
- Molina, A.; Sánchez, J.; Sánchez, W.; Vielma, V. Platelet-rich plasma as an adjuvant in the endometrial preparation of patients with refractory endometrium. JBRA Assist. Reprod. 2018, 22, 42. [Google Scholar] [CrossRef]
- Santamaria, X.; Cabanillas, S.; Cervello, I.; Arbona, C.; Raga, F.; Ferro, J.; Palmero, J.; Remohí, J.; Pellicer, A.; Simón, C. Autologous cell therapy with CD133+ bone marrow-derived stem cells for refractory Asherman’s syndrome and endometrial atrophy: A pilot cohort study. Hum. Reprod. 2016, 31, 1087–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tersoglio, A.E.; Tersoglio, S.; Salatino, D.R.; Castro, M.; Gonzalez, A.; Hinojosa, M.; Castellano, O. Regenerative therapy by endometrial mesenchymal stem cells in thin endometrium with repeated implantation failure. A novel strategy. JBRA Assist. Reprod. 2020, 24, 118. [Google Scholar] [CrossRef]
- Jing, Z.; Qiong, Z.; Yonggang, W.; Yanping, L. Rat bone marrow mesenchymal stem cells improve regeneration of thin endometrium in rat. Fertil. Steril. 2014, 101, 587–594. [Google Scholar] [CrossRef]
- Simon, A.; Laufer, N. Assessment and treatment of repeated implantation failure (RIF). J. Assist. Reprod. Genet. 2012, 29, 1227–1239. [Google Scholar] [CrossRef] [Green Version]
- Achache, H.; Revel, A. Endometrial receptivity markers, the journey to successful embryo implantation. Hum. Reprod. Update 2006, 12, 731–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukui, A.; Funamizu, A.; Yokota, M.; Yamada, K.; Nakamua, R.; Fukuhara, R.; Kimura, H.; Mizunuma, H. Uterine and circulating natural killer cells and their roles in women with recurrent pregnancy loss, implantation failure and preeclampsia. J. Reprod. Immunol. 2011, 90, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Gonen-Gross, T.; Goldman-Wohl, D.; Huppertz, B.; Lankry, D.; Greenfield, C.; Natanson-Yaron, S.; Hamani, Y.; Gilad, R.; Yagel, S.; Mandelboim, O. Inhibitory NK receptor recognition of HLA-G: Regulation by contact residues and by cell specific expression at the fetal-maternal interface. PLoS ONE 2010, 5, e8941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asghari, S.; Valizadeh, A.; Aghebati-Maleki, L.; Nouri, M.; Yousefi, M. Endometriosis: Perspective, lights, and shadows of etiology. Biomed. Pharmacother. 2018, 106, 163–174. [Google Scholar] [CrossRef]
- Abdolmohammadi-Vahid, S.; Pashazadeh, F.; Pourmoghaddam, Z.; Aghebati-Maleki, L.; Abdollahi-Fard, S.; Yousefi, M. The effectiveness of IVIG therapy in pregnancy and live birth rate of women with recurrent implantation failure (RIF): A systematic review and meta-analysis. J. Reprod. Immunol. 2019, 134, 28–33. [Google Scholar] [CrossRef]
- Practice Committee of the American Society for Reproductive Medicine. Definitions of infertility and recurrent pregnancy loss: A committee opinion. Fertil. Steril. 2013, 99, 63. [Google Scholar] [CrossRef]
- Pourakbari, R.; Ahmadi, H.; Yousefi, M.; Aghebati-Maleki, L. Cell therapy in female infertility-related diseases: Emphasis on recurrent miscarriage and repeated implantation failure. Life Sci. 2020, 258, 118181. [Google Scholar] [CrossRef]
- Yu, N.; Zhang, B.; Xu, M.; Wang, S.; Liu, R.; Wu, J.; Yang, J.; Feng, L. Intrauterine administration of autologous peripheral blood mononuclear cells (PBMC s) activated by HCG improves the implantation and pregnancy rates in patients with repeated implantation failure: A prospective randomized study. Am. J. Reprod. Immunol. 2016, 76, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Okitsu, O.; Kiyokawa, M.; Oda, T.; Miyake, K.; Sato, Y.; Fujiwara, H. Intrauterine administration of autologous peripheral blood mononuclear cells increases clinical pregnancy rates in frozen/thawed embryo transfer cycles of patients with repeated implantation failure. J. Reprod. Immunol. 2011, 92, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, S.; Fujiwara, H.; Nakayama, T.; Kosaka, K.; Mori, T.; Fujii, S. Intrauterine administration of autologous peripheral blood mononuclear cells promotes implantation rates in patients with repeated failure of IVF–embryo transfer. Hum. Reprod. 2006, 21, 3290–3294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashii, K.; Fujiwara, H.; Yoshioka, S.; Kataoka, N.; Yamada, S.; Hirano, T.; Mori, T.; Fujii, S.; Maeda, M. Peripheral blood mononuclear cells stimulate progesterone production by luteal cells derived from pregnant and non-pregnant women: Possible involvement of interleukin-4 and interleukin-10 in corpus luteum function and differentiation. Hum. Reprod. (Oxf. Engl.) 1998, 13, 2738–2744. [Google Scholar] [CrossRef] [Green Version]
- Jensen, F.; Muzzio, D.; Soldati, R.; Fest, S.; Zenclussen, A.C. Regulatory B10 cells restore pregnancy tolerance in a mouse model. Biol. Reprod. 2013, 89, 90-1–90-7. [Google Scholar] [CrossRef] [Green Version]
- Gentile, T.; Borel, I.M.; Angelucci, J.; Miranda, S.; Margni, R.A. Preferential synthesis of asymmetric antibodies in rats immunized with paternal particulate antigens. Effect on pregnancy. J. Reprod. Immunol. 1992, 22, 173–183. [Google Scholar] [CrossRef]
- Sato, Y.; Fujiwara, H.; Zeng, B.-X.; Higuchi, T.; Yoshioka, S.; Fujii, S. Platelet-derived soluble factors induce human extravillous trophoblast migration and differentiation: Platelets are a possible regulator of trophoblast infiltration into maternal spiral arteries. Blood 2005, 106, 428–435. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, K.; Fujiwara, H.; Sato, Y.; Zeng, B.-X.; Fujii, H.; Yoshioka, S.; Nishi, E.; Nishio, T. Platelets are novel regulators of neovascularization and luteinization during human corpus luteum formation. Endocrinology 2007, 148, 3056–3064. [Google Scholar] [CrossRef]
- Fujiwara, H. Immune cells contribute to systemic cross-talk between the embryo and mother during early pregnancy in cooperation with the endocrine system. Reprod. Med. Biol. 2006, 5, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Metcalf, E. The effect of platelet-rich plasma (PRP) on intraluminal fluid and pregnancy rates in mares susceptible to persistent mating-induced endometritis (PMIE). J. Equine Vet. Sci. 2014, 1, 128. [Google Scholar] [CrossRef]
- Zadehmodarres, S.; Salehpour, S.; Saharkhiz, N.; Nazari, L. Treatment of thin endometrium with autologous platelet-rich plasma: A pilot study. JBRA Assist. Reprod. 2017, 21, 54. [Google Scholar] [CrossRef]
- Sadighi-Moghaddam, B.; Salek Farrokhi, A.; Namdar Ahmadabad, H.; Barati, M.; Moazzeni, S.M. Mesenchymal stem cell therapy prevents abortion in CBA/J× DBA/2 mating. Reprod. Sci. 2018, 25, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- Dmowski, W.P.; Greenblatt, R.B. Asherman’s syndrome and risk of placenta accreta. Obstet. Gynecol. 1969, 34, 288–299. [Google Scholar]
- Chirputkar, R.; Vaidya, A. Understanding Infertility and the Potential Role of Stem Cells in Infertility Treatment: A Short Com-munication. Int. J. Reprod. Fertil. Sex Health 2015, 2, 37–40. [Google Scholar]
- Yu, D.; Wong, Y.; Cheong, Y.; Xia, E.; Li, T.C. Asherman syndrome—one century later. Fertil. Steril. 2008, 89, 759–779. [Google Scholar] [CrossRef]
- March, C.M. Asherman’s Syndrome. Semin. Reprod. Med. 2011, 29, 83–94. [Google Scholar] [CrossRef]
- Lee, S.Y.; Shin, J.E.; Kwon, H.; Choi, D.H.; Kim, J.H. Effect of autologous adipose-derived stromal vascular fraction transplantation on endometrial regeneration in patients of Asherman’s syndrome: A pilot study. Reprod. Sci. 2020, 27, 561–568. [Google Scholar] [CrossRef]
- Singh, N.; Mohanty, S.; Seth, T.; Shankar, M.; Bhaskaran, S.; Dharmendra, S. Autologous stem cell transplantation in refractory Asherman’s syndrome: A novel cell based therapy. J. Hum. Reprod. Sci. 2014, 7, 93. [Google Scholar] [CrossRef]
- Nagori, C.B.; Panchal, S.Y.; Patel, H. Endometrial regeneration using autologous adult stem cells followed by conception by in vitro fertilization in a patient of severe Asherman’s syndrome. J. Hum. Reprod. Sci. 2011, 4, 43–48. [Google Scholar]
- Tan, J.; Li, P.; Wang, Q.; Li, Y.; Li, X.; Zhao, D.; Xu, X.; Kong, L. Autologous menstrual blood-derived stromal cells transplantation for severe Asherman’s syndrome. Hum. Reprod. 2016, 31, 2723–2729. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Qi, W.; Zheng, J.; Tian, Y.; Qi, X.; Kong, D.; Zhang, J.; Huang, X. Exosomes derived from adipose mesenchymal stem cells restore functional endometrium in a rat model of intrauterine adhesions. Reprod. Sci. 2020, 27, 1266–1275. [Google Scholar] [CrossRef]
- Xin, L.; Lin, X.; Zhou, F.; Li, C.; Wang, X.; Yu, H.; Pan, Y.; Fei, H.; Ma, L.; Zhang, S. A scaffold laden with mesenchymal stem cell-derived exosomes for promoting endometrium regeneration and fertility restoration through macrophage immunomodulation. Acta Biomater. 2020, 113, 252–266. [Google Scholar] [CrossRef]
- Yao, Y.; Chen, R.; Wang, G.; Zhang, Y.; Liu, F. Exosomes derived from mesenchymal stem cells reverse EMT via TGF-β1/Smad pathway and promote repair of damaged endometrium. Stem Cell Res. Ther. 2019, 10, 225. [Google Scholar] [CrossRef]
- Saribas, G.S.; Ozogul, C.; Tiryaki, M.; Pinarli, F.A.; Kilic, S.H. Effects of uterus derived mesenchymal stem cells and their exosomes on Asherman’s syndrome. Acta Histochem. 2020, 122, 151465. [Google Scholar] [CrossRef]
- Perrini, C.; Strillacci, M.G.; Bagnato, A.; Esposti, P.; Marini, M.G.; Corradetti, B.; Bizzaro, D.; Idda, A.; Ledda, S.; Capra, E. Microvesicles secreted from equine amniotic-derived cells and their potential role in reducing inflammation in endometrial cells in an in-vitro model. Stem Cell Res. Ther. 2016, 7, 169. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-H.; Chen, Y.-H.; Chang, H.-Y.; Au, H.-K.; Tzeng, C.-R.; Huang, Y.-H. Chronic niche inflammation in endometriosis-associated infertility: Current understanding and future therapeutic strategies. Int. J. Mol. Sci. 2018, 19, 2385. [Google Scholar] [CrossRef] [Green Version]
- Parasar, P.; Ozcan, P.; Terry, K.L. Endometriosis: Epidemiology, diagnosis and clinical management. Curr. Obstet. Gynecol. Rep. 2017, 6, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Bungum, H.F.; Vestergaard, C.; Knudsen, U.B. Endometriosis and type 1 allergies/immediate type hypersensitivity: A systematic review. Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 179, 209–215. [Google Scholar] [CrossRef]
- Nielsen, N.M.; Jørgensen, K.T.; Pedersen, B.V.; Rostgaard, K.; Frisch, M. The co-occurrence of endometriosis with multiple sclerosis, systemic lupus erythematosus and Sjögren syndrome. Hum. Reprod. 2011, 26, 1555–1559. [Google Scholar] [CrossRef] [Green Version]
- Mu, F.; Rich-Edwards, J.; Rimm, E.B.; Spiegelman, D.; Forman, J.P.; Missmer, S.A. Association between endometriosis and hypercholesterolemia or hypertension. Hypertension 2017, 70, 59–65. [Google Scholar] [CrossRef]
- Kok, V.C.; Tsai, H.-J.; Su, C.-F.; Lee, C.-K. The risks for ovarian, endometrial, breast, colorectal, and other cancers in women with newly diagnosed endometriosis or adenomyosis: A population-based study. Int. J. Gynecol. Cancer 2015, 25, 968–976. [Google Scholar] [CrossRef]
- Králíčková, M.; Vetvicka, V. Endometriosis and ovarian cancer. World J. Clin. Oncol. 2014, 5, 800. [Google Scholar] [CrossRef]
- Zhao, Y.-x.; Chen, S.-r.; Su, P.-p.; Huang, F.-h.; Shi, Y.-c.; Shi, Q.-y.; Lin, S. Using mesenchymal stem cells to treat female infertility: An update on female reproductive diseases. Stem Cells Int. 2019, 2019, 9071720. [Google Scholar] [CrossRef]
- Vercellini, P.; Viganò, P.; Somigliana, E.; Fedele, L. Endometriosis: Pathogenesis and treatment. Nat. Rev. Endocrinol. 2014, 10, 261. [Google Scholar] [CrossRef]
- Maruyama, T.; Yoshimura, Y. Stem cell theory for the pathogenesis of endometriosis. Front. Biosci. 2012, 4, 2754–2763. [Google Scholar] [CrossRef] [Green Version]
- Serdar, E.; Bulun, E. Mechanisms of Disease. N. Engl. J. Med. 2009, 360, 268–279. [Google Scholar]
- Dunselman, G.; Vermeulen, N.; Becker, C.; Calhaz-Jorge, C.; D’Hooghe, T.; De Bie, B.; Heikinheimo, O.; Horne, A.; Kiesel, L.; Nap, A. ESHRE guideline: Management of women with endometriosis. Hum. Reprod. 2014, 29, 400–412. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Z.; Yang, F.; Wang, H.; Liang, S.; Wang, H.; Yang, J.; Lin, J. The role of endometrial stem cells in the pathogenesis of endometriosis and their application to its early diagnosis. Biol. Reprod. 2020, 102, 1153–1159. [Google Scholar] [CrossRef] [PubMed]
- Simoni, M.; Taylor, H.S. Therapeutic strategies involving uterine stem cells in reproductive medicine. Curr. Opin. Obstet. Gynecol. 2018, 30, 209–216. [Google Scholar] [CrossRef]
- PAIN, O.P. Treatment of pelvic pain associated with endometriosis: A committee opinion. Fertil. Steril. 2014, 101, 927–935. [Google Scholar]
- Johnson, N.P.; Hummelshoj, L.; Consortium, W.E.S.M.; Abrao, M.; Adamson, G.; Allaire, C.; Amelung, V.; Andersson, E.; Becker, C.; Birna Árdal, K. Consensus on current management of endometriosis. Hum. Reprod. 2013, 28, 1552–1568. [Google Scholar] [CrossRef] [PubMed]
- Zito, G.; Luppi, S.; Giolo, E.; Martinelli, M.; Venturin, I.; Di Lorenzo, G.; Ricci, G. Medical treatments for endometriosis-associated pelvic pain. BioMed. Res. Int. 2014, 2014, 191967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopalakrishnan, A.K.; Pandit, H.; Metkari, S.; Warty, N.; Madan, T. Adenoviral vector encoding soluble Flt-1 engineered human endometrial mesenchymal stem cells effectively regress endometriotic lesions in NOD/SCID mice. Gene Ther. 2016, 23, 580–591. [Google Scholar] [CrossRef]
- Hajazimian, S.; Maleki, M.; Mehrabad, S.D.; Isazadeh, A. Human Wharton’s jelly stem cells inhibit endometriosis through apoptosis induction. Reproduction 2020, 159, 549–558. [Google Scholar] [CrossRef]
- LA, E. Problems in the diagnosis of adenomyosis uteri; with special reference to dysfunctional bleeding. West. J. Surg. Obstet. Gynecol. 1956, 64, 291–304, discussion, 304. [Google Scholar]
- Ferenczy, A. Pathophysiology of adenomyosis. Hum. Reprod. Update 1998, 4, 312–322. [Google Scholar] [CrossRef] [Green Version]
- Israel, L.S.; Woutersz, T.B. Adenomyosis: A neglected diagnosis. Obstet. Gynecol. 1959, 14, 168–173. [Google Scholar]
- Wood, C. Surgical and medical treatment of adenomyosis. Hum. Reprod. Update 1998, 4, 323–336. [Google Scholar] [CrossRef] [Green Version]
- Garcia, L.; Isaacson, K. Adenomyosis: Review of the literature. J. Minim. Invasive Gynecol. 2011, 18, 428–437. [Google Scholar] [CrossRef]
- Kim, M.D.; Kim, S.; Kim, N.K.; Lee, M.H.; Ahn, E.H.; Kim, H.J.; Cho, J.H.; Cha, S.H. Long-term results of uterine artery embolization for symptomatic adenomyosis. Am. J. Roentgenol. 2007, 188, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.-H.; Liu, W.-M.; Fuh, J.-L.; Cheng, M.-H.; Chao, H.-T. Comparison of surgery alone and combined surgical-medical treatment in the management of symptomatic uterine adenomyoma. Fertil. Steril. 2009, 92, 876–885. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.R.; Nathanson, H.G.; Milim, S.J.; Haselkorn, J.S. Laparoscopic bipolar coagulation for the conservative treatment of adenomyomata. J. Am. Assoc. Gynecol. Laparosc. 1996, 4, 19–24. [Google Scholar] [CrossRef]
- Levgur, M. Therapeutic options for adenomyosis: A review. Arch. Gynecol. Obstet. 2007, 276, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Fujishita, A.; Masuzaki, H.; Khan, K.N.; Kitajima, M.; Ishimaru, T. Modified reduction surgery for adenomyosis. Gynecol. Obstet. Investig. 2004, 57, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-J.; Li, H.-Y.; Chang, Y.-L.; Yuan, C.-C.; Tai, L.-K.; Lu, K.H.; Chang, C.-M.; Chiou, S.-H. Suppression of migratory/invasive ability and induction of apoptosis in adenomyo-sis-derived mesenchymal stem cells by cyclooxygenase-2 inhibitors. Fertil. Steril. 2010, 94, 1972–1979. [Google Scholar] [CrossRef]
- World Health Organisation. WHO Laboratory Manual for the Examination of Human Semen and Sperm-Cervical Mucus Interaction; Cambridge University Press: Cambridge, MA, USA, 1999. [Google Scholar]
- Harris, I.D.; Fronczak, C.; Roth, L.; Meacham, R.B. Fertility and the aging male. Rev. Urol. 2011, 13, e184. [Google Scholar]
- Tüttelmann, F. Classification of andrological disorders. In Andrology: Male Reproductive Health and Dysfunction; Nieschlag, E., Behre, H.M., Nieschlag, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 4, pp. 87–92. [Google Scholar]
- Lipshultz, L.I.; Howards, S.S.; Niederberger, C.S. Infertility in the Male; Cambridge University Press: Cambridge, MA, USA, 2009. [Google Scholar]
- Irvine, D.S. Epidemiology and aetiology of male infertility. Hum. Reprod. 1998, 13, 33–44. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Liu, X.; Peng, J.; He, D.; Lin, T.; Zhu, J.; Li, X.; Zhang, Y.; Wei, G. Potential spermatogenesis recovery with bone marrow mesenchymal stem cells in an azoospermic rat model. Int. J. Mol. Sci. 2014, 15, 13151–13165. [Google Scholar] [CrossRef]
- Blackhall, F.H.; Atkinson, A.; Maaya, M.; Ryder, W.D.J.; Horne, G.; Brison, D.R.; Lieberman, B.; Radford, J.A. Semen cryopreservation, utilisation and reproductive outcome in men treated for Hodgkin’s disease. Br. J. Cancer 2002, 87, 381–384. [Google Scholar] [CrossRef] [Green Version]
- Fang, F.; Li, Z.; Zhao, Q.; Li, H.; Xiong, C. Human induced pluripotent stem cells and male infertility: An overview of current progress and perspectives. Hum. Reprod. 2018, 33, 188–195. [Google Scholar] [CrossRef]
- Esteves, S.C.; Miyaoka, R.; Agarwal, A. Surgical treatment of male infertility in the era of intracytoplasmic sperm injection-new insights. Clinics 2011, 66, 1463–1478. [Google Scholar] [CrossRef] [Green Version]
- Qamar, A.Y.; Fang, X.; Kim, M.J.; Cho, J. Improved viability and fertility of frozen-thawed dog sperm using adipose-derived mesenchymal stem cells. Sci. Rep. 2020, 10, 7034. [Google Scholar] [CrossRef]
- Qamar, A.Y.; Fang, X.; Kim, M.J.; Cho, J. Improved post-thaw quality of canine semen after treatment with exosomes from conditioned medium of adipose-derived mesenchymal stem cells. Animals 2019, 9, 865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghasemzadeh-Hasankolaei, M.; Eslaminejad, M.B.; Sedighi-Gilani, M. Derivation of male germ cells from ram bone marrow mesenchymal stem cells by three different methods and evaluation of their fate after transplantation into the testis. Vitr. Cell. Dev. Biol.-Anim. 2016, 52, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Xie, L.; Mohsin, A.; Ahmed, W.; Xu, C.; Peng, Y.; Hang, H.; Zhuang, Y.; Chu, J.; Guo, M. Efficient generation of male germ-like cells derived during co-culturing of adipose-derived mesenchymal stem cells with Sertoli cells under retinoic acid and testosterone induction. Stem Cell Res. Ther. 2019, 10, 91. [Google Scholar] [CrossRef] [PubMed]
- Karimaghai, N.; Tamadon, A.; Rahmanifar, F.; Mehrabani, D.; Jahromi, A.R.; Zare, S.; Khodabandeh, Z.; Jahromi, I.R.; Koohi-Hoseinabadi, O.; Dianatpour, M. Spermatogenesis after transplantation of adipose tissue-derived mesenchymal stem cells in busulfan-induced azoospermic hamster. Iran. J. Basic Med. Sci. 2018, 21, 660–667. [Google Scholar]
- Tamadon, A.; Mehrabani, D.; Rahmanifar, F.; Jahromi, A.R.; Panahi, M.; Zare, S.; Khodabandeh, Z.; Jahromi, I.R.; Tanideh, N.; Dianatpour, M. Induction of spermatogenesis by bone marrow-derived mesenchymal stem cells in busulfan-induced azoospermia in hamster. Int. J. Stem Cells 2015, 8, 134. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Yang, S.; Yang, H.; Liu, Y.; Liu, Y.; Hai, Y.; Chen, Z.; Guo, Y.; Gong, Y.; Gao, W.-Q. Generation of male differentiated germ cells from various types of stem cells. Reproduction 2014, 147, R179–R188. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Banks, S.M.; Barnes, K.M.; Sherins, R.J. Two-year comparison of testicular responses to pulsatile gonadotropin-releasing hormone and exogenous gonadotropins from the inception of therapy in men with isolated hypogonadotropic hypogonadism. J. Clin. Endocrinol. Metab. 1988, 67, 1140–1145. [Google Scholar] [CrossRef] [PubMed]
- Rohayem, J.; Hauffa, B.P.; Zacharin, M.; Kliesch, S.; Zitzmann, M.; German Adolescent Hypogonadotropic Hypogonadism Study Group; Wüsthof, A.; Tittel, B.; Tittel, B.; Morlot, M. Testicular growth and spermatogenesis: New goals for pubertal hormone replacement in boys with hypogonadotropic hypogonadism? -a multicentre prospective study of hCG/rFSH treatment outcomes during adolescence-. Clin. Endocrinol. 2017, 86, 75–87. [Google Scholar] [CrossRef]
- Sani, R.N.; Tajik, P.; Yousefi, M.H.; Movahedin, M.; Qasemi-Panahi, B.; Shafiei, S.; Hamedani, M.A. Follicle stimulating hormone increases spermatogonial stem cell colonization during in vitro co-culture. In Veterinary Research Forum: An International Quarterly Journal; Faculty of Veterinary Medicine, Urmia University: Urmia, Iran, 2013; p. 37. [Google Scholar]
- Tajik, P.; Sani, R.N.; Moezifar, M.; Yousefi, M.; Movahedin, M.; Qasemi-Panahi, B.; Shafiei, S.; Fili, P.R. Effect of follicle-stimulating hormone and testosterone on colony formation of bovine spermatogonial stem cell. Comp. Clin. Pathol. 2014, 23, 901–906. [Google Scholar] [CrossRef]
- Su, J.S.; Farber, N.J.; Vij, S.C. Pathophysiology and treatment options of varicocele: An overview. Andrologia 2020, 53, e13576. [Google Scholar] [CrossRef]
- Agarwal, A.; Hamada, A.; Esteves, S.C. Insight into oxidative stress in varicocele-associated male infertility: Part 1. Nat. Rev. Urol. 2012, 9, 678. [Google Scholar] [CrossRef] [PubMed]
- Raheem, A.A.; Ralph, D. Male infertility: Causes and investigations. Trends Urol. Men’s Health 2011, 2, 8–11. [Google Scholar] [CrossRef]
- Goldstein, M.; Eid, J.-F. Elevation of intratesticular and scrotal skin surface temperature in men with varicocele. J. Urol. 1989, 142, 743–745. [Google Scholar] [CrossRef]
- Lipshultz, L.I.; Corriere, J.N., Jr. Progressive testicular atrophy in the varicocele patient. J. Urol. 1977, 117, 175–176. [Google Scholar] [CrossRef]
- Rashtbari, H.; Razi, M.; Hassani-Bafrani, H.; Najaran, H. Berberine reinforces Sertoli cells niche and accelerates spermatogonial stem cells renewal in experimentally-induced varicocele condition in rats. Phytomedicine 2018, 40, 68–78. [Google Scholar] [CrossRef]
- Liu, H.-C.; Xie, Y.; Deng, C.-H.; Liu, G.-H. Stem cell-based therapies for fertility preservation in males: Current status and future prospects. World J. Stem Cells 2020, 12, 1097. [Google Scholar] [CrossRef]
- Mohammad, S.; Aghamir, K.; Salavati, A.; Yousefie, R.; Tootian, Z.; Ghazaleh, N.; Jamali, M.; Azimi, P. Does Bone Marrow–derived Mesenchymal Stem Cell Transfusion Prevent Antisperm Antibody Production After Traumatic Testis Rupture? Urology 2014, 86, 82–86. [Google Scholar] [CrossRef]
- Addas, F.; Yan, S.; Hadjipavlou, M.; Gonsalves, M.; Sabbagh, S. Testicular Rupture or Testicular Fracture? A Case Report and Literature Review. Case Rep. Urol. 2018, 2018, 1323780. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasa, G.; Kavitha, P.; Chaithra, P.; Vineeth, V.; Kumar, C.S.; Malini, S.S. Clinical Significance of antisperm antibody analysis in evaluating male infertility of south karnataka. Age 2011, 20, 31–40. [Google Scholar]
- Fahlbusch, B.; Fahlbusch, M.; Thon, W. Blunt testicular injury-conservative or surgical treatment? Aktuelle Urol. 2003, 34, 176–178. [Google Scholar] [PubMed]
- Haas, G.G., Jr.; Manganiello, P. A double-blind, placebo-controlled study of the use of methylprednisolone in infertile men with sperm-associated immunoglobulins. Fertil. Steril. 1987, 47, 295–301. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Abdi, R.; Fiorina, P.; Adra, C.N.; Atkinson, M.; Sayegh, M.H. Immunomodulation by mesenchymal stem cells: A potential therapeutic strategy for type 1 diabetes. Diabetes 2008, 57, 1759–1767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corcione, A.; Benvenuto, F.; Ferretti, E.; Giunti, D.; Cappiello, V.; Cazzanti, F.; Risso, M.; Gualandi, F.; Mancardi, G.L.; Pistoia, V. Human mesenchymal stem cells modulate B-cell functions. Blood 2006, 107, 367–372. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, C.-H.; Ji, A.T.-Q.; Chang, C.-C.; Cheng, C.-J.; Lee, L.M.; Ho, J.H.-C. Local injection of mesenchymal stem cells protects testicular torsion-induced germ cell injury. Stem Cell Res. Ther. 2015, 6, 113. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm Filho, D.; Torres, M.A.; Bordin, A.L.; Crezcynski-Pasa, T.B.; Boveris, A. Spermatic cord torsion, reactive oxygen and nitrogen species and ischemia–reperfusion injury. Mol. Asp. Med. 2004, 25, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Reyes, J.G.; Farias, J.G.; Henríquez-Olavarrieta, S.; Madrid, E.; Parraga, M.; Zepeda, A.B.; Moreno, R.D. The hypoxic testicle: Physiology and pathophysiology. Oxidative Med. Cell. Longev. 2012, 2012, 929285. [Google Scholar] [CrossRef]
- Dokmeci, D. Testicular torsion, oxidative stress and the role of antioxidant therapy. Folia Med. 2006, 48, 16–21. [Google Scholar]
- Mansbach, J.M.; Forbes, P.; Peters, C. Article. Arch. Pediatrics Adolesc. Med. 2005, 159, 1167–1171. [Google Scholar] [CrossRef] [PubMed]
- Azizollahi, S.; Aflatoonian, R.; Gilani, M.A.S.; Behnam, B.; Tajik, N.; Asghari-Jafarabadi, M.; Reza Asgari, H.; Koruji, M. Alteration of spermatogenesis following spermatogonial stem cells transplantation in testicular torsion-detorsion mice. J. Assist. Reprod. Genet. 2016, 33, 771–781. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, C.-H.; Ji, A.T.-Q.; Chang, C.-C.; Cheng, C.-J.; Lee, L.M.; Ho, J.H.-C. Mesenchymal stem cells restore the sperm motility from testicular torsion-detorsion injury by regulation of glucose metabolism in sperm. Stem Cell Res. Ther. 2019, 10, 270. [Google Scholar] [CrossRef]
- Zhong, L.; Yang, M.; Zou, X.; Du, T.; Xu, H.; Sun, J. Human umbilical cord multipotent mesenchymal stromal cells alleviate acute ischemia-reperfusion injury of spermatogenic cells via reducing inflammatory response and oxidative stress. Stem Cell Res. Ther. 2020, 11, 294. [Google Scholar] [CrossRef]
- Klinefelter, H.F., Jr.; Reifenstein, E.C., Jr.; Albright, F., Jr. Syndrome characterized by gynecomastia, aspermatogenesis without A-Leydigism, and increased excretion of follicle-stimulating hormone. J. Clin. Endocrinol. 1942, 2, 615–627. [Google Scholar] [CrossRef]
- Ferlin, A.; Arredi, B.; Foresta, C. Genetic causes of male infertility. Reprod. Toxicol. 2006, 22, 133–141. [Google Scholar] [CrossRef]
- Bojesen, A.; Juul, S.; Gravholt, C.H. Prenatal and postnatal prevalence of Klinefelter syndrome: A national registry study. J. Clin. Endocrinol. Metab. 2003, 88, 622–626. [Google Scholar] [CrossRef]
- Mehta, A.; Bolyakov, A.; Roosma, J.; Schlegel, P.N.; Paduch, D.A. Successful testicular sperm retrieval in adolescents with Klinefelter syndrome treated with at least 1 year of topical testosterone and aromatase inhibitor. Fertil. Steril. 2013, 100, 970–974. [Google Scholar] [CrossRef] [PubMed]
- Wyns, C.; Curaba, M.; Vanabelle, B.; Van Langendonckt, A.; Donnez, J. Options for fertility preservation in prepubertal boys. Hum. Reprod. Update 2010, 16, 312–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, K.-J.; Jeong, S.-M.; Park, S.-J.; Song, K.-H.; Choi, H.-J.; Lee, Y.-W.; Yoon, K.-Y.; Shin, S.-T.; Cho, S.-W.; Cho, J.-K. A Case of the Periorchiepididymitis in a Dog. J. Vet. Clin. 2008, 25, 40–43. [Google Scholar]
- Malolina, E.A.; Kulibin, A.Y.; Kushch, A.A. Neonatal testicular cell transplantation restores murine spermatogenesis damaged in the course of herpes simplex virus-induced orchitis. Reprod. Fertil. Dev. 2016, 28, 757–764. [Google Scholar] [CrossRef]
- Kaver, I.; Matzkin, H.; Braf, Z.F. Epididymo-orchitis: A retrospective study of 121 patients. J. Fam. Pract. 1990, 30, 548–553. [Google Scholar]
- Kadish, H.A.; Bolte, R.G. A retrospective review of pediatric patients with epididymitis, testicular torsion, and torsion of testicular appendages. Pediatrics 1998, 102, 73–76. [Google Scholar] [CrossRef]
- Luzzi, G.; O’brien, T. Acute epididymitis. BJU Int. 2001, 87, 747–755. [Google Scholar] [CrossRef]
- Thomas, T.H.; Lishnak, T.S.; Heiman, D. Epididymitis and orchitis: An overview. Am. Fam. Physician 2009, 79, 583–587. [Google Scholar]
- Ludwig, M. Diagnosis and therapy of acute prostatitis, epididymitis and orchitis. Andrologia 2008, 40, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Manavi, K.; Turner, K.; Scott, G.; Stewart, L. Audit on the management of epididymo-orchitis by the Department of Urology in Edinburgh. Int. J. STD AIDS 2005, 16, 386–387. [Google Scholar] [CrossRef] [PubMed]
- Erpenbach, K.H. Systemic treatment with Interferon-α2b: An effective method to prevent sterility after bilateral mumps orchitis. J. Urol. 1991, 146, 54–56. [Google Scholar] [CrossRef]
- Ku, J.-H.; Yim, Y.-S.; Lee, C.-H.; Kim, Y.-H.; Song, Y.-S.; Jeon, Y.-S.; Kim, M.-E.; Lee, N.-K.; Park, Y.-H. The Preventive Effect of Systemic Treatment with Interferon-${\alpha} 2B $ for Infertility of Mumps Orchitis. Clin. Exp. Reprod. Med. 1999, 26, 111–116. [Google Scholar]
- Yeniyol, C.O.; Sorguc, S.; Minareci, S.; Ayder, A.R. Role of interferon-alpha-2B in prevention of testicular atrophy with unilateral mumps orchitis. Urology 2000, 55, 931–933. [Google Scholar] [CrossRef]
- Sherif, I.O.; Abdel-Aziz, A.; Sarhan, O.M. Cisplatin-induced testicular toxicity in rats: The protective effect of arjunolic acid. J. Biochem. Mol. Toxicol. 2014, 28, 515–521. [Google Scholar] [CrossRef]
- Awadalla, E.A. Ameliorative effect of the crude oil of the Nigella sativa on oxidative stress induced in rat testes by cisplatin treatment. Biomed. Prev. Nutr. 2012, 2, 265–268. [Google Scholar] [CrossRef]
- Jahnukainen, K.; Mitchell, R.T.; Stukenborg, J.-B. Testicular function and fertility preservation after treatment for haematological cancer. Curr. Opin. Endocrinol. Diabetes Obes. 2015, 22, 217–223. [Google Scholar] [CrossRef]
- Clermont, Y. Kinetics of spermatogenesis in mammals: Seminiferous epithelium cycle and spermatogonial renewal. Physiol. Rev. 1972, 52, 198–236. [Google Scholar] [CrossRef]
- De Michele, F.; Vermeulen, M.; Wyns, C. Fertility restoration with spermatogonial stem cells. Curr. Opin. Endocrinol. Diabetes Obes. 2017, 24, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-J.; Yan, J.; Zou, X.-L.; Guo, K.-J.; Zhao, Y.; Meng, C.-Y.; Yin, F.; Guo, L. Bone marrow mesenchymal stem cells repair cadmium-induced rat testis injury by inhibiting mitochondrial apoptosis. Chem.-Biol. Interact. 2017, 271, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Sherif, I.O.; Sabry, D.; Abdel-Aziz, A.; Sarhan, O.M. The role of mesenchymal stem cells in chemotherapy-induced gonadotoxicity. Stem Cell Res. Ther. 2018, 9, 196. [Google Scholar] [CrossRef]
- Mohammed, F.Z.; Hassan Abdallah, S.; Hassan Watad, S. Treatment of cyclophosphamide induced infertile male mice with HSCs that homed by honey, bovine colostrum and umbilical cord blood derived mesenchymal stem cells. Biochem. Lett. 2019, 15, 98–113. [Google Scholar] [CrossRef]
- Abdelaziz, M.H.; Salah EL-Din, E.Y.; El-Dakdoky, M.H.; Ahmed, T.A. The impact of mesenchymal stem cells on doxorubicin-induced testicular toxicity and progeny outcome of male prepubertal rats. Birth Defects Res. 2019, 111, 906–919. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.-T.; Xiong, C.-L.; Liu, T.-S.; Shen, S.-L.; Rao, J.-P.; Qiu, F. Secretions released from mesenchymal stem cells improve spermatogenesis restoration of cytotoxic treatment with busulfan in azoospermia mice. Andrologia 2021, 53, e14144. [Google Scholar] [CrossRef]
- Cai, Y.t.; Xiong, C.l.; Shen, S.l.; Rao, J.p.; Liu, T.s.; Qiu, F. Mesenchymal stem cell-secreted factors delayed spermatogenesis injuries induced by busulfan involving intercellular adhesion molecule regulation. Andrologia 2019, 51, e13285. [Google Scholar] [CrossRef]
- Deng, C.; Xie, Y.; Zhang, C.; Ouyang, B.; Chen, H.; Lv, L.; Yao, J.; Liang, X.; Zhang, Y.; Sun, X. Urine-derived stem cells facilitate endogenous spermatogenesis restoration of busulfan-induced nonobstructive azoospermic mice by paracrine exosomes. Stem Cells Dev. 2019, 28, 1322–1333. [Google Scholar] [CrossRef]
- Mobarak, H.; Heidarpour, M.; Rahbarghazi, R.; Nouri, M.; Mahdipour, M. Amniotic fluid-derived exosomes improved spermatogenesis in a rat model of azoospermia. Life Sci. 2021, 274, 119336. [Google Scholar] [CrossRef]
- Zhou, Q.-Z.; Liu, C.-D.; Guo, X.-B.; Zhai, J.-W.; Xia, H.; Yang, J.-K.; Zhou, J.-H.; Guo, W.-B.; Yang, C.; Xia, M.; et al. Protective effect of bone marrow mesenchymal stem cell-derived exosomes against the reproductive toxicity of cyclophosphamide is associated with the p38MAPK/ERK and AKT signaling pathways. Asian J. Androl. 2021, 23, 386. [Google Scholar] [CrossRef]
- Salek, F.; Baharara, J.; Shahrokhabadi, K.N.; Amini, E. The guardians of germ cells; Sertoli-derived exosomes against electromagnetic field-induced oxidative stress in mouse spermatogonial stem cells. Theriogenology 2021, 173, 112–122. [Google Scholar] [CrossRef]
- Zhang, W.; Yang, C.; Guo, W.; Guo, X.; Bian, J.; Zhou, Q.; Chen, M.; Zhou, J.; Chen, Z.; Wang, P.; et al. Protective effect of bone marrow mesenchymal stem cells-derived exosomes against testicular ischemia-reperfusion injury in rats. Nan Fang Yi Ke Da Xue Xue Bao J. South. Med Univ. 2018, 38, 910–916. [Google Scholar] [CrossRef]
- Abdelaal, N.E.; Tanga, B.M.; Abdelgawad, M.; Allam, S.; Fathi, M.; Saadeldin, I.M.; Bang, S.; Cho, J. Cellular Therapy via Spermatogonial Stem Cells for Treating Impaired Spermatogenesis, Non-Obstructive Azoospermia. Cells 2021, 10, 1779. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Female Infertility Category | Affection | Evs Source/Route of Administration in Vivo | Main Mechanism of Actions | Reference |
|---|---|---|---|---|
| Premature ovarian failure | Premature ovarian insufficiency (in vivo, mice model) | HUCMSCs/intravenous injection | Upregulation of AKT, p-AKT, and angiogenic cytokines (including VEGF, IGF, and angiogenin) | [65] |
| Ovarian granulosa cell cisplatin-treated (in vitro, rats model) | HUCMSCs | Bcl-2 and caspase-3 were upregulated, whilst the expression of Bax, cleaved caspase-3, and cleaved PARP were downregulated | [66] | |
| Ovarian granulosa cell cisplatin-treated (in vitro, rats model) | HUCMSCs | Upregulated E2, StAR, and Bcl-2/Bax ratio downregulated Caspase 3 | [68] | |
| Premature ovarian insufficiency (in vivo, mice model) & human granulosa-lutein cells (in vitro) | HAECs/intraovarian injection | 109 cytokines involved in apoptosis, angiogenesis, cell cycle and immune response (in vivo) Decreasing apoptosis and TGF-β/Smad signaling pathway (in vitro) | [69] | |
| Premature ovarian insufficiency (in vivo, mice model) | BMSCs/intravenous injection | miR-664-5p targeted p53 and apoptosis | [70] | |
| Premature ovarian insufficiency (in vivo, rats model) | BMSCs/intraperitoneal injection | miR-144-5p targeted PTEN and apoptosis | [71] | |
| Premature ovarian insufficiency (in vivo, mice model) | HAECs/intraovarian injection | miR-1246 targeted phosphatidylinositol signaling and apoptosis pathways | [72] | |
| Premature ovarian insufficiency (in vivo, mice model) | AMSCs/intraovarian injection | miR-320a targeted SIRT4 and ROS formation | [73] | |
| PCOS | Cumulus cells of PCOS patients (in vitro) | Human AMSCs | miR-323-3p promoted cell proliferation and inhibited apoptosis in CCs through targeting PDCD4 | [85] |
| Endometrial injuries | Rats subjected to mechanical endometrial damage | BMSCs/intravenous injection | miR-340 downregulated collagen 1α1, α-SMA and transforming growth factor (TGF)-β1 | [99] |
| Mice injured endometrial epithelial cells (in vitro) | HUCMSCs | Inhibited IL-6, IL-1β, TLR4 and RelA Increased TNFA | [100] | |
| Human endometrial stromal cells injury (in vitro) | HUCMSCs | Increasing Bcl-2 level and downregulating Cleaved Caspase-3 level and activated the PTEN/AKT signaling pathway to regulate the proliferation and antiapoptosis | [101] | |
| Asherman’s syndrome | Rat model (in vivo) | ADMSCs/intrauterine injection | Enhanced the expression of integrin-β3, LIF, and VEGF | [137] |
| Rat model (in vivo) | HUCMSCs & collagen scaffold/endometrial transplantation | miRNAs facilitated CD163+ M2 macrophage polarization, reduced inflammation, and increased anti-inflammatory responses | [138] | |
| Rabbit model (in vivo) | BMSCs/Uterine muscle injection | CK19 level significantly increased whereas VIM level significantly decreased TGF-β1, TGF-β1R, and Smad2 mRNA were all significantly decreased | [139] | |
| Rat model (in vivo) | Uterus MSCs/intrauterine injection | MMP-2 and MMP-9 expression was enhanced TIMP-2 expression was decreased | [140] | |
| Endometriosis | Lipopolysaccharides effects on endometrial cell (equines, in vitro) | Equine AMSCs | Reduced tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), interleukin 1β (IL-1β), and metalloproteinases (MMP) 1 and 13, and the release of some pro- or anti-inflammatory cytokines. | [141] |
| Model of Disease | Therapeutic Intervention/Route of Administration | Core Findings | Reference |
|---|---|---|---|
| Busulfan-induced NOA mice model | Urine-derived stem cells -derived EVs/Intratesticular | spermatogenic genes (Pou5f1, Prm1, SYCP3, and DAZL) and the spermatogenic protein UCHL1 were significantly increased after 36 days of injection | [242] |
| Busulfan-induced NOA rats model | Amniotic fluid-derived EVs/Intratesticular | DAZL and VASA were increased significantly. Sperm parameters and spermatogenesis index were significantly improved. OCT-3/4+ cells were increased in NOA rats after AF-Exos injection, showing the restoration of spermatogenesis. | [243] |
| Cyclophosphamide-induced testicular spermatogenic dysfunction | Bone marrow mesenchymal stem cell-derived EVs/intravenous | Increased spermatogonia cell proliferation and reduced apoptosis. Phosphorylated levels of ERK, AKT, and p38MAPK proteins were reduced | [244] |
| Electromagnetic field-induced oxidative stress in mouse spermatogonial stem cells (in vitro) | Sertoli cells-derived EVs | down-regulation of the apoptotic gene (Caspase-3), and oxidative stress. Up-regulation of SSCs specific gene (GFRα1). | [245] |
| Testicular ischemia-reperfusion injury in rats | Bone marrow mesenchymal stem cell-derived EVs/Intratesticular | Reduced HMGB1, caspase-3, and cleaved caspase-3 | [246] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qamar, A.Y.; Hussain, T.; Rafique, M.K.; Bang, S.; Tanga, B.M.; Seong, G.; Fang, X.; Saadeldin, I.M.; Cho, J. The Role of Stem Cells and Their Derived Extracellular Vesicles in Restoring Female and Male Fertility. Cells 2021, 10, 2460. https://doi.org/10.3390/cells10092460
Qamar AY, Hussain T, Rafique MK, Bang S, Tanga BM, Seong G, Fang X, Saadeldin IM, Cho J. The Role of Stem Cells and Their Derived Extracellular Vesicles in Restoring Female and Male Fertility. Cells. 2021; 10(9):2460. https://doi.org/10.3390/cells10092460
Chicago/Turabian StyleQamar, Ahmad Yar, Tariq Hussain, Muhammad Kamran Rafique, Seonggyu Bang, Bereket Molla Tanga, Gyeonghwan Seong, Xun Fang, Islam M. Saadeldin, and Jongki Cho. 2021. "The Role of Stem Cells and Their Derived Extracellular Vesicles in Restoring Female and Male Fertility" Cells 10, no. 9: 2460. https://doi.org/10.3390/cells10092460