
The TLR9 2848C/T Polymorphism Is Associated with the CMV DNAemia among HIV/CMV Co-Infected Patients

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Detection of TLR9 Polymorphisms
2.3. Assessment of CMV DNAemia
2.4. Measurement of Cytokine Levels
2.5. Statistical Analyses
3. Results
3.1. Heterozygous and Homozygous Recessive Genotypes of the TLR9 2848C/T SNP Are Prevalent in HIV/CMV Co-Infected Patients with CMV DNAemia
3.2. Mutation Present in at Least One Allele of the TLR9 2848C/T SNP Occurs More Frequently in HIV/CMV Co-Infected Patients with CMV Viremia
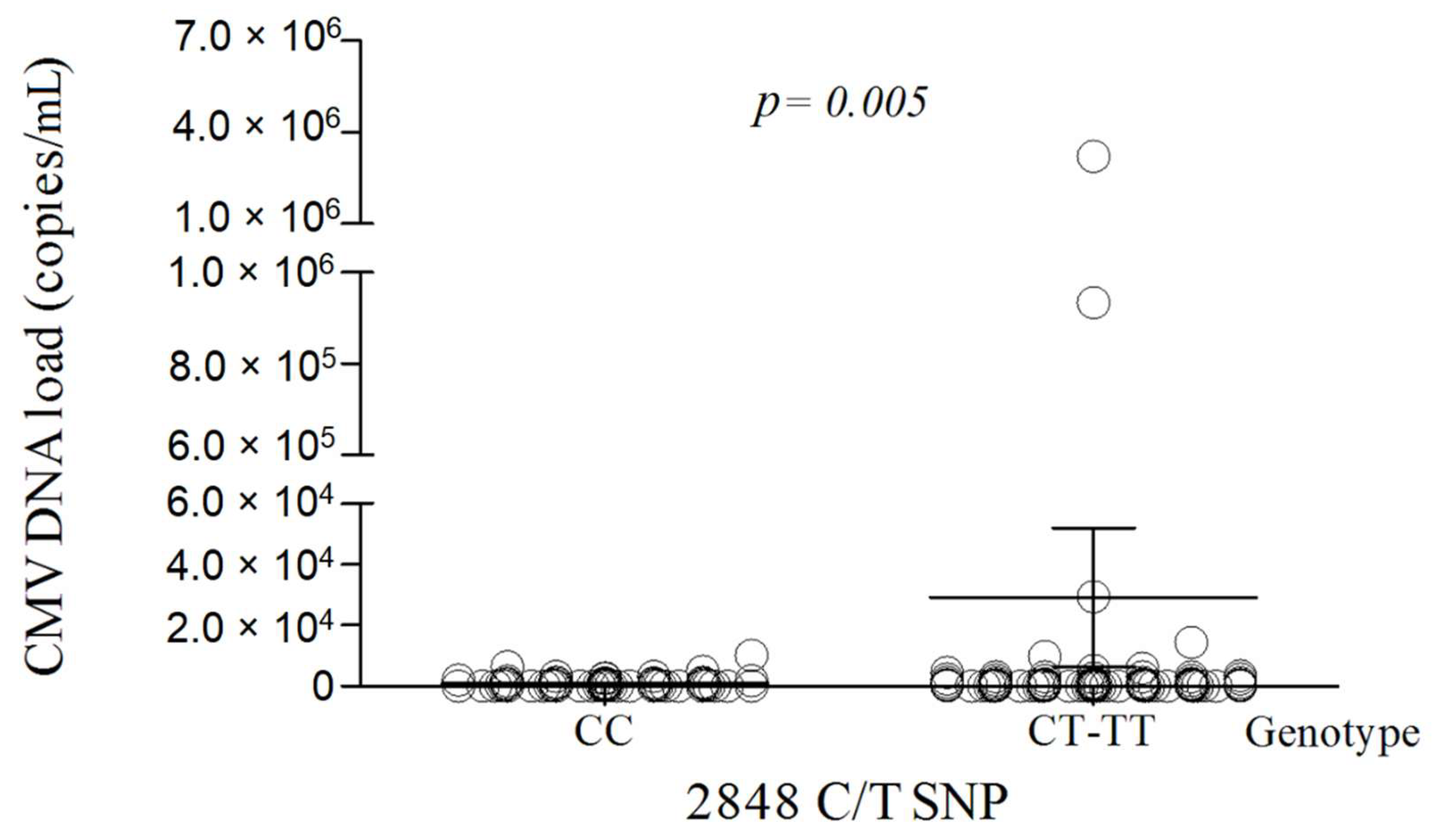
3.3. The TLR9 2848C/T Polymorphism Is Associated with the CMV Viremia
3.4. The TGT Haplotype of TLR9 −1237T/C, 1174G/A, and 2848C/T SNPs Is Prevalent in HIV/CMV Co-Infected Patients
3.5. The TLR9 2848C/T SNP Is Associated with Lower Serum IFN-β Level
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palella, F.J., Jr.; Delaney, K.M.; Moorman, A.C.; Loveless, M.O.; Fuhrer, J.; Satten, G.A.; Aschman, D.J.; Holmberg, S.D. Declining morbidity and mortality among patients with advanced human immunodeficiency virus infection. HIV outpatient study investigators. N. Engl. J. Med. 1998, 338, 853–860. [Google Scholar] [CrossRef] [PubMed]
- Emery, V.C.; Asher, K.; de Juan Sanjuan, C. Importance of the cytomegalovirus seropositive recipient as a contributor to disease burden after solid organ transplantation. J. Clin. Virol. 2012, 54, 125–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitlick, M.M.; Orr, K.; Momany, A.M.; McDonald, E.L.; Murray, J.C.; Ryckman, K.K. Determining the prevalence of cytomegalovirus infection in a cohort of preterm infants. J. Neonatal. Perinatal. Med. 2015, 8, 137–141. [Google Scholar] [CrossRef] [Green Version]
- Drew, W.L.; Lalezari, J.P. Cytomegalovirus and HIV. In HIV InSite Knowledge Base Chapter; University of California: San Francisco, CA, USA, 2006; Available online: http://hivinsite.ucsf.edu/InSite?page=kb-00&doc=kb-05-03-03 (accessed on 3 February 2021).
- Ho, M. The history of cytomegalovirus and its diseases. Med. Microbiol. Immunol. 2008, 197, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Medzhitov, R. Toll-like receptor control of the adaptive immune responses. Nat. Immunol. 2004, 5, 987–995. [Google Scholar] [CrossRef]
- Joshi, A.; Punke, E.B.; Mehmetoglu-Gurbuz, T.; Peralta, D.P.; Garg, H. TLR9 polymorphism correlates with immune activation, CD4 decline and plasma IP10 levels in HIV patients. BMC Infect. Dis. 2019, 19, 56. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, I.; Caux, C.; Hasan, U.; Bendriss-Vermare, N.; Olive, D. Impaired toll-like receptor 7 and 9 signaling: From chronic viral infections to cancer. Trends Immunol. 2010, 31, 391–397. [Google Scholar] [CrossRef]
- Paradowska, E.; Jabłońska, A.; Studzińska, M.; Skowrońska, K.; Suski, P.; Wiśniewska-Ligier, M.; Woźniakowska-Gęsicka, T.; Nowakowska, D.; Gaj, Z.; Wilczyński, J.; et al. TLR9 -1486T/C and 2848C/T are associated with human cytomegalovirus infection in infants. PLoS ONE 2016, 11, e0154100. [Google Scholar] [CrossRef]
- Seo, S.; Fan, W.; Hansen, J.A.; Storer, B.E.; Pergam, S.A.; Green, M.L.; Ozkok, S.; Goyal, S.; Fisher, C.; Hingorani, S.; et al. Evaluation of published single-nucleotide polymorphisms (SNPs) associated with cytomegalovirus infection and disease after hematopoietic cell transplantation in a large Caucasian cohort. Blood 2015, 124, 182. [Google Scholar] [CrossRef]
- Xiao, H.W.; Luo, Y.; Lai, X.Y.; Shi, J.M.; Tan, Y.M.; He, J.S.; Xie, W.Z.; Zheng, W.Y.; Ye, X.J.; Yu, X.H.; et al. Donor TLR9 gene tagSNPs influence susceptibility to aGVHD and CMV reactivation in the allo-HSCT setting without polymorphisms in the TLR4 and NOD2 genes. Bone Marrow Transplant. 2014, 49, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, P.; Singh, A.; Kesarwani, P.; Jaiswal, P.K.; Singh, V.; Mittal, R.D. Association studies of toll-like receptor gene polymorphisms with allograft survival in renal transplant recipients of North India. Clin. Transplant. 2012, 26, 581–588. [Google Scholar] [CrossRef]
- Kim, T.H.; Jeong, K.H.; Kim, S.K.; Lee, S.H.; Ihm, C.G.; Lee, T.W.; Moon, J.Y.; Yoon, Y.C.; Chung, J.H.; Park, S.J.; et al. TLR9 gene polymorphism (rs187084, rs352140): Association with acute rejection and estimated glomerular filtration rate in renal transplant recipients. Int. J. Immunogenet. 2013, 40, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Wujcicka, W.; Paradowska, E.; Studzińska, M.; Gaj, Z.; Wilczyński, J.; Leśnikowski, Z.; Nowakowska, D. TLR9 2848 GA heterozygotic status possibly predisposes fetuses and newborns to congenital infection with human cytomegalovirus. PLoS ONE 2015, 10, e0122831. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Ruiz, M.; Corrales, I.; Arias, M.; Campistol, J.M.; Giménez, E.; Crespo, J.; López-Oliva, M.O.; Beneyto, I.; Martín-Moreno, P.L.; Llamas-Fuente, F.; et al. Association between individual and combined SNPs in genes related to innate immunity and incidence of CMV infection in seropositive kidney transplant recipients. Am. J. Transplant. 2015, 15, 1323–1335. [Google Scholar] [CrossRef] [PubMed]
- Krüger, B.; Banas, M.C.; Walberer, A.; Böger, C.A.; Farkas, S.; Hoffmann, U.; Fischereder, M.; Banas, B.; Krämer, B.K. A comprehensive genotype-phenotype interaction of different toll-like receptor variations in a renal transplant cohort. Clin. Sci. 2010, 119, 535–544. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, A.; Cunha, C.; Carotti, A.; Aloisi, T.; Guarrera, O.; Di Ianni, M.; Falzetti, F.; Bistoni, F.; Aversa, F.; Pitzurra, L.; et al. Polymorphisms in toll-like receptor genes and susceptibility to infections in allogeneic stem cell transplantation. Exp. Hematol. 2009, 37, 1022–1029. [Google Scholar] [CrossRef]
- Beima-Sofie, K.; Wamalwa, D.; Maleche-Obimbo, E.; Lingappa, J.R.; Mackelprang, R.; Gantt, S.; John-Stewart, G.; Casper, C.; Slyker, J.A. Toll-like receptor 9 polymorphism is associated with increased Epstein-Barr virus and Cytomegalovirus acquisition in HIV-exposed infants. AIDS 2018, 32, 267–270. [Google Scholar] [CrossRef]
- Bochud, P.Y.; Hersberger, M.; Taffé, P.; Bochud, M.; Stein, C.M.; Rodrigues, S.D.; Calandra, T.; Francioli, P.; Telenti, A.; Speck, R.F.; et al. Polymorphisms in toll-like receptor 9 influence the clinical course of HIV-1 infection. AIDS 2007, 21, 441–446. [Google Scholar] [CrossRef]
- Pine, S.O.; McElrath, M.; Bochud, P.Y. Polymorphisms in toll-like receptor 4 and toll-like receptor 9 influence viral load in a seroincident cohort of HIV-1-infected individuals. AIDS 2009, 23, 2387–2395. [Google Scholar] [CrossRef]
- Shaikh, N.; Nirmalkar, A.; Thakar, M. Polymorphisms in toll-like receptors (TLRs)-7 and 9 genes in Indian population with progressive and nonprogressive HIV-1 infection. AIDS Res. Hum. Retrovir. 2019, 35, 577–582. [Google Scholar] [CrossRef] [Green Version]
- Mackelprang, R.D.; Carrington, M.; Thomas, K.K.; Hughes, J.P.; Baeten, J.M.; Wald, A.; Farquhar, C.; Fife, K.; Campbell, M.S.; Kapiga, S.; et al. Host genetic and viral determinants of HIV-1 RNA set point among HIV-1 seroconverters from sub-saharan Africa. J. Virol. 2015, 89, 2104–2111. [Google Scholar] [CrossRef] [Green Version]
- Valverde-Villegas, J.M.; Dos Santos, B.P.; de Medeiros, R.M.; Mattevi, V.S.; Lazzaretti, R.K.; Sprinz, E.; Kuhmmer, R.; Chies, J.A. Endosomal toll-like receptor gene polymorphisms and susceptibility to HIV and HCV co-infection—Differential influence in individuals with distinct ethnic background. Hum. Immunol. 2017, 78, 221–226. [Google Scholar] [CrossRef]
- Ricci, E.; Malacrida, S.; Zanchetta, M.; Mosconi, I.; Montagna, M.; Giaquinto, C.; De Rossi, A. Toll-like receptor 9 polymorphisms influence mother-to-child transmission of human immunodeficiency virus type 1. J. Transl. Med. 2010, 8, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soriano-Sarabia, N.; Vallejo, A.; Ramírez-Lorca, R.; Rodríguez Mdel, M.; Salinas, A.; Pulido, I.; Sáez, M.E.; Leal, M. Influence of the toll-like receptor 9 1635A/G polymorphism on the CD4 count, HIV viral load, and clinical progression. J. Acquir. Immune Defic. Syndr. 2008, 49, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Said, E.A.; Al-Yafei, F.; Zadjali, F.; Hasson, S.S.; Al-Balushi, M.S.; Al-Mahruqi, S.; Koh, C.Y.; Al-Naamani, K.; Al-Busaidi, J.Z.; Idris, M.A.; et al. Association of single-nucleotide polymorphisms in TLR7 (Gln11Leu) and TLR9 (1635A/G) with a higher CD4T cell count during HIV infection. Immunol. Lett. 2014, 160, 58–64. [Google Scholar] [CrossRef]
- Royse, K.E.; Kempf, M.C.; McGwin, G., Jr.; Wilson, C.M.; Tang, J.; Shrestha, S. Toll-like receptor gene variants associated with bacterial vaginosis among HIV-1 infected adolescents. J. Reprod. Immunol. 2012, 96, 84–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; He, H.; Sun, C.; Fu, J.; Ghosh, D.; Deng, C.; Sheng, Y. Association of toll-like receptor polymorphisms with acquisition of HIV infection and clinical findings: A protocol for systematic review and meta-analysis. Medicine 2020, 99, e236632020. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.T.H.; Van’t Hof, R.; Crockett, J.C.; Hope, M.E.; Berry, S.; Thomson, J.; McLean, M.H.; McColl, K.E.; El-Omar, E.M.; Hold, G.L. Increase in NF-kappaB binding affinity of the variant C allele of the toll-like receptor 9 -1237T/C polymorphism is associated with Helicobacter pylori-induced gastric disease. Infect. Immun. 2010, 78, 1345–1352. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, A.; Osório, N.S.; Saraiva, M.; Cunha, C.; Almeida, A.J.; Teixeira-Coelho, M.; Ludovico, P.; Pedrosa, J.; Pitzurra, L.; Aversa, F.; et al. The C allele of rs5743836 polymorphism in the human TLR9 promoter links IL-6 and TLR9 up-regulation and confers increased B-cell proliferation. PLoS ONE 2011, 6, e28256. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulos, A.I.; Ferwerda, B.; Antoniadou, A.; Sakka, V.; Galani, L.; Kavatha, D.; Panagopoulos, P.; Poulakou, G.; Kanellakopoulou, K.; van der Meer, J.W.; et al. Association of toll-like receptor 4 Asp299Gly and Thr399Ile polymorphisms with increased infection risk in patients with advanced HIV-1 infection. Clin. Infect. Dis. 2010, 51, 242–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paradowska, E.; Przepiórkiewicz, M.; Nowakowska, D.; Studzińska, M.; Wilczyński, J.; Emery, V.C.; Leśnikowski, Z.J. Detection of cytomegalovirus in human placental cells by polymerase chain reaction. APMIS 2006, 114, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, R.; Klimecki, W.T.; Raby, B.A.; Vercelli, D.; Palmer, L.J.; Kwiatkowski, D.J.; Silverman, E.K.; Martinez, F.; Weiss, S.T. Single-nucleotide polymorphisms in the toll-like receptor 9 gene (TLR9): Frequencies, pairwise linkage disequilibrium, and haplotypes in three U.S. ethnic groups and exploratory case-control disease association studies. Genomics 2003, 81, 85–91. [Google Scholar] [CrossRef]
- Singh, V.; Srivastava, N.; Kapoor, R.; Mittal, R.D. Single-nucleotide polymorphisms in genes encoding toll-like receptor -2, -3, -4, and -9 in a case-control study with bladder cancer susceptibility in a North Indian population. Arch. Med. Res. 2013, 44, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Mandal, R.K.; George, G.P.; Mittal, R.D. Association of toll-like receptor (TLR) 2, 3 and 9 genes polymorphism with prostate cancer risk in North Indian population. Mol. Biol. Rep. 2012, 39, 7263–7269. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.N.; Pisetsky, D.S.; Schwartz, D.A. Toll-like receptors in the pathogenesis of human disease. Nat. Immunol. 2004, 5, 975–979. [Google Scholar] [CrossRef]
- Puel, A.; Picard, C.; Ku, C.L.; Smahi, A.; Casanova, J.L. Inherited disorders of NF-kappaB-mediated immunity in man. Curr. Opin. Immunol. 2004, 16, 34–41. [Google Scholar] [CrossRef]
- Barton, G.M.; Kagan, J.C.; Medzhitov, R. Intracellular localization of toll-like receptor 9 prevents recognition of self DNA but facilitates access to viral DNA. Nat. Immunol. 2006, 7, 49–56. [Google Scholar] [CrossRef]
- Ito, R.; Ota, M.; Meguro, A.; Katsuyama, Y.; Uemoto, R.; Nomura, E.; Nishide, T.; Kitaichi, N.; Horie, Y.; Namba, K.; et al. Investigation of association between TLR9 gene polymorphisms and VKH in Japanese patients. Ocul. Immunol. Inflamm. 2011, 19, 202–205. [Google Scholar] [CrossRef]
- Tabeta, K.; Georgel, P.; Janssen, E.; Du, X.; Hoebe, K.; Crozat, K.; Mudd, S.; Shamel, L.; Sovath, S.; Goode, J.; et al. Toll-like receptors 9 and 3 as essential components of innate immune defense against mouse cytomegalovirus infection. Proc. Natl. Acad. Sci. USA 2004, 101, 3516–3521. [Google Scholar] [CrossRef] [Green Version]
- Bozza, S.; Gaziano, R.; Bonifazi, P.; Zelante, T.; Pitzurra, L.; Montagnoli, C.; Moretti, S.; Castronari, R.; Sinibaldi, P.; Rasi, G.; et al. Thymosin alpha1 activates the TLR9/MyD88/IRF7-dependent murine cytomegalovirus sensing for induction of anti-viral responses in vivo. Int. Immunol. 2007, 19, 1261–1270. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Poltorak, A.; Wei, Y.; Beutler, B. Three novel mammalian toll-like receptors: Gene structure, expression, and evolution. Eur. Cytokine. Netw. 2000, 11, 362–371. [Google Scholar]
- Funderburg, N.; Luciano, A.A.; Jiang, W.; Rodriguez, B.; Sieg, S.F.; Lederman, M.M. Toll-like receptor ligands induce human T cell activation and death, a model for HIV pathogenesis. PLoS ONE 2008, 3, e1915. [Google Scholar] [CrossRef] [Green Version]
- Kastelijn, E.A.; van Moorsel, C.H.; Rijkers, G.T.; Ruven, H.J.; Karthaus, V.; Kwakkel-van Erp, J.M.; van de Graaf, E.A.; Zanen, P.; van Kessel, D.A.; Grutters, J.C.; et al. Polymorphisms in innate immunity genes associated with development of bronchiolitis obliterans after lung transplantation. J. Heart. Lung. Transplant. 2010, 29, 665–671. [Google Scholar] [CrossRef]
- Tao, K.; Fujii, M.; Tsukumo, S.; Maekawa, Y.; Kishihara, K.; Kimoto, Y.; Horiuchi, T.; Hisaeda, H.; Akira, S.; Kagami, S.; et al. Genetic variations of toll-like receptor 9 predispose to systemic lupus erythematosus in Japanese population. Ann. Rheum. Dis. 2007, 66, 905–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi, K.; Lian, Z.X.; Kimura, Y.; Selmi, C.; Yang, G.X.; Gordon, S.C.; Invernizzi, P.; Podda, M.; Coppel, R.L.; Ansari, A.A.; et al. Genetic polymorphisms of toll-like receptor 9 influence the immune response to CpG and contribute to hyper-IgM in primary biliary cirrhosis. J. Autoimmun. 2005, 24, 347–352. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Variables | Results |
|---|---|
| Age (median and range, years) | 37.2 (17–72 years) |
| Sex, n (%) | |
| female | 46/205 (22.4) |
| male | 159/205 (77.6) |
| CD4 count, n (%) | |
| ≤200 cells/mm3 | 43/205 (21.0) |
| >200 cells/mm3 | 162/205 (79.0) |
| CD4 nadir, n (%) | |
| ≤200 cells/mm3 | 135/205 (65.9) |
| >200 cells/mm3 | 70/205 (34.1) |
| CMV IgG, n (%) | |
| positive | 192/205 (93.7) |
| negative | 13/205 (6.3) |
| HBsAg positive, n (%) | 34/205 (16.6) |
| anti HCV positive, n (%) | 54/205 (26.3) |
| HIV RNA load, n (%) | |
| <20 HIV RNA copies/mL | 170/205 (82.9) |
| >20 HIV RNA copies/mL | 35/205 (17.1) |
| Self-reported HIV-1 transmission route | |
| MSM | 107/205 (52.2) |
| HET | 44/205 (21.5) |
| MSM/HET | 5/205 (2.4) |
| IDU | 49/205 (23.9) |
| TLR9 SNP | Genotype/Allele | Patients with CMV DNAemia, n (%) | Patients without CMV DNAemia, n (%) | p-Value |
|---|---|---|---|---|
| −1237T/C | TT | 117 (96.7) | 82 (97.6) | 1.000 |
| (rs5743836) | TC | 4 (3.3) | 2 (2.4) | 1.000 |
| T | 238 (98.3) | 166 (98.8) | 1.000 | |
| C | 4 (1.7) | 2 (1.2) | 1.000 | |
| −1486T/C | TT | 31 (25.6) | 28 (33.3) | 0.273 |
| (rs187084) | TC | 57 (47.1) | 31 (36.9) | 0.155 |
| CC | 33 (27.3) | 25 (29.8) | 0.753 | |
| T | 119 (49.2) | 87 (51.8) | 0.617 | |
| C | 123 (50.8) | 81 (48.2) | 0.617 | |
| 1174G/A | GG | 49 (40.5) | 34 (40.5) | 1.000 |
| (rs352139) | GA | 62 (51.2) | 40 (47.6) | 0.671 |
| AA | 10 (8.3) | 10 (11.9) | 0.474 | |
| G | 160 (66.1) | 108 (64.3) | 0.752 | |
| A | 82 (33.9) | 60 (35.7) | 0.752 | |
| 2848C/T | CC | 29 (24.0) | 36 (42.8) | 0.006 |
| (rs352140) | CT | 58 (47.9) | 33 (39.3) | 0.254 |
| TT | 34 (28.1) | 15 (17.9) | 0.099 | |
| C | 116 (47.9) | 105 (62.5) | 0.005 | |
| T | 126 (52.1) | 63 (37.5) | 0.005 |
| TLR9 | Model | Genotype | Genotype Frequencies; n (%) 1 | Unadjusted | Adjusted 2 | |||
|---|---|---|---|---|---|---|---|---|
| SNPs | Patients with CMV DNAemia | Patients without CMV DNAemia | OR (95% CI) | p-Value | OR (95% CI) | p-Value | ||
| −1237T/C | - | TT | 117 (96.7) | 82 (97.6) | 1.00 | 0.7 | 1.00 | 1 |
| TC | 4 (3.3) | 2 (2.4) | 1.40 (0.25–7.83) | 1.00 (0.17–5.90) | ||||
| 1174G/A | codominant | GG | 49 (40.5) | 34 (40.5) | 1.00 | 0.67 | 1.00 | 0.44 |
| GA | 62 (51.2) | 40 (47.6) | 1.08 (0.60–1.94) | 1.04 (0.55–1.97) | ||||
| AA | 10 (8.3) | 10 (11.9) | 0.69 (0.26–1.85) | 0.54 (0.19–1.51) | ||||
| dominant | GG | 49 (40.5) | 34 (40.5) | 1.00 | 1 | 1.00 | 0.79 | |
| GA-GG | 72 (59.5) | 50 (59.5) | 1.00 (0.57–1.76) | 0.92 (0.50–1.69) | ||||
| recessive | GG-GA | 111 (91.7) | 74 (88.1) | 1.00 | 0.39 | 1.00 | 0.2 | |
| AA | 10 (8.3) | 10 (11.9) | 0.67 (0.26–1.68) | 0.53 (0.20–1.40) | ||||
| overdominant | GG-AA | 59 (48.8) | 44 (52.4) | 1.00 | 0.61 | 1.00 | 0.6 | |
| GA | 62 (51.2) | 40 (47.6) | 1.16 (0.66–2.02) | 1.17 (0.64–2.14) | ||||
| 2848C/T | codominant | CC | 29 (24.0) | 36 (42.8) | 1.00 | 0.014 | 1.00 | 0.058 |
| CT | 58 (47.9) | 33 (39.3) | 2.18 (1.14–4.18) | 1.88 (0.89–3.95) | ||||
| TT | 34 (28.1) | 15 (17.9) | 2.81 (1.29–6.14) | 2.69 (1.15–6.25) | ||||
| dominant | CC | 29 (24.0) | 36 (42.8) | 1.00 | 0.004 | 1.00 | 0.027 | |
| CT-TT | 92 (76.0) | 48 (57.2) | 2.38 (1.30–4.34) | 2.15 (1.09–4.26) | ||||
| recessive | CC-CT | 87 (71.9) | 69 (82.1) | 1.00 | 0.087 | 1.00 | 0.089 | |
| TT | 34 (28.1) | 15 (17.9) | 1.80 (0.91–3.57) | 1.86 (0.90–3.83) | ||||
| overdominant | CC-TT | 63 (52.1) | 51 (60.7) | 1.00 | 0.22 | 1.00 | 0.61 | |
| CT | 58 (47.9) | 33 (39.3) | 1.42 (0.81–2.50) | 1.18 (0.63–2.20) | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jabłońska, A.; Jabłonowska, E.; Studzińska, M.; Kamerys, J.; Paradowska, E. The TLR9 2848C/T Polymorphism Is Associated with the CMV DNAemia among HIV/CMV Co-Infected Patients. Cells 2021, 10, 2360. https://doi.org/10.3390/cells10092360
Jabłońska A, Jabłonowska E, Studzińska M, Kamerys J, Paradowska E. The TLR9 2848C/T Polymorphism Is Associated with the CMV DNAemia among HIV/CMV Co-Infected Patients. Cells. 2021; 10(9):2360. https://doi.org/10.3390/cells10092360
Chicago/Turabian StyleJabłońska, Agnieszka, Elżbieta Jabłonowska, Mirosława Studzińska, Juliusz Kamerys, and Edyta Paradowska. 2021. "The TLR9 2848C/T Polymorphism Is Associated with the CMV DNAemia among HIV/CMV Co-Infected Patients" Cells 10, no. 9: 2360. https://doi.org/10.3390/cells10092360