
The Structure of the Spinal Cord Ependymal Region in Adult Humans Is a Distinctive Trait among Mammals

 , , , and
, , , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
2.1.1. Naked Mole-Rats
2.1.2. Hyh Mice
2.1.3. Chimpanzee
2.1.4. Macaque
2.2. Human Tissue
2.3. Histology and Immunohistochemistry
2.3.1. Nissl Staining
2.3.2. Immunohistochemistry
2.4. Transmission Electron Microscopy
3. Results
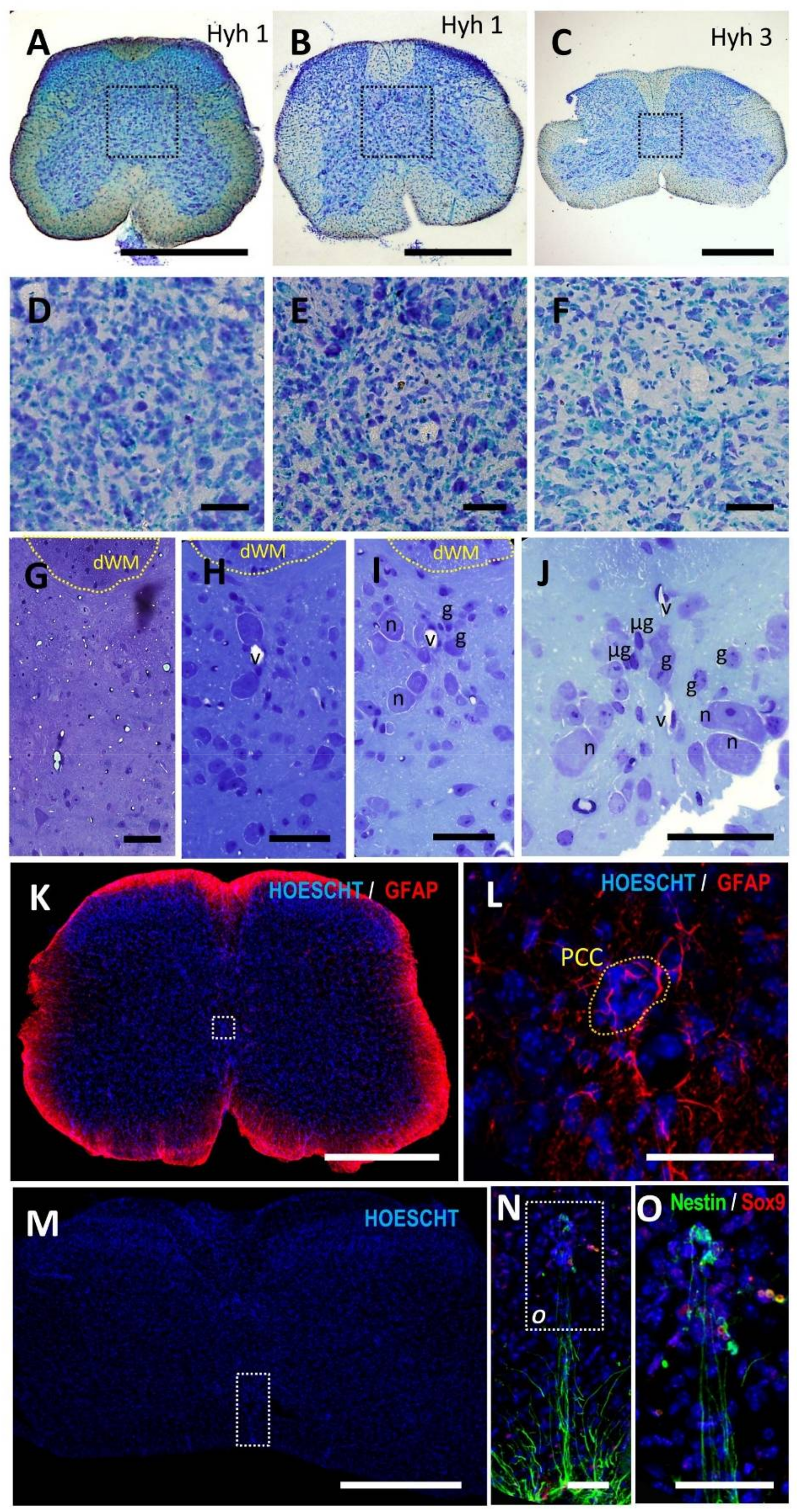
3.1. Adult Naked Mole-Rats (Heterocephalus Glaber) and the Hyh (Hydrocephalus with Hop Gait) Mutant Mice Lack Central Canal, but the Substituting Structure Is Notably Different from That in Humans
3.1.1. Naked Mole-Rats
3.1.2. Hyh Mice
3.2. The Closure of the Central Canal in the Adult Human Ependymal Region Is Preceded by a Delamination That Shows Features of Epithelial to Mesenchymal Transition (EMT)
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mokalled, M.H.; Patra, C.; Dickson, A.L.; Endo, T.; Stainier, D.Y.R.; Poss, K.D. Injury-Induced Ctgfa Directs Glial Bridging and Spinal Cord Regeneration in Zebrafish. Science 2016, 354, 630–634. [Google Scholar] [CrossRef] [Green Version]
- Becker, C.G.; Becker, T. Neuronal Regeneration from Ependymo-Radial Glial Cells: Cook, Little Pot, Cook! Dev. Cell 2015, 32, 516–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, T.; Becker, C.G. Dynamic Cell Interactions Allow Spinal Cord Regeneration in Zebrafish. Curr. Opin. Physiol. 2020, 14, 64–69. [Google Scholar] [CrossRef]
- Alfaro-Cervello, C.; Soriano-Navarro, M.; Mirzadeh, Z.; Alvarez-Buylla, A.; Garcia-Verdugo, J.M. Biciliated Ependymal Cell Proliferation Contributes to Spinal Cord Growth. J. Comp. Neurol. 2012, 15, 3528–3552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfaro-Cervello, C.; Cebrian-Silla, A.; Soriano-Navarro, M.; Garcia-Tarraga, P.; Matias-Guiu, J.; Gomez-Pinedo, U.; Molina Aguilar, P.; Alvarez-Buylla, A.; Luquin, M.R.; Garcia-Verdugo, J.M. The Adult Macaque Spinal Cord Central Canal Zone Contains Proliferative Cells and Closely Resembles the Human. J. Comp. Neurol. 2014, 522, 1800–1817. [Google Scholar] [CrossRef] [PubMed]
- Ghazale, H.; Ripoll, C.; Leventoux, N.; Jacob, L.; Azar, S.; Mamaeva, D.; Glasson, Y.; Calvo, C.F.; Thomas, J.L.; Meneceur, S.; et al. RNA Profiling of the Human and Mouse Spinal Cord Stem Cell Niches Reveals an Embryonic-like Regionalization with MSX1+ Roof-Plate-Derived Cells. Stem Cell Rep. 2019, 12, 1159–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hugnot, J.P.; Franzen, R. The Spinal Cord Ependymal Region: A Stem Cell Niche in the Caudal Central Nervous System. Front. Biosci. 2011, 16, 1044–1059. [Google Scholar] [CrossRef]
- Becker, C.G.; Becker, T.; Hugnot, J.-P. The Spinal Ependymal Zone as a Source of Endogenous Repair Cells across Vertebrates. Prog. Neurobiol. 2018, 170, 67–80. [Google Scholar] [CrossRef]
- Garcia-Ovejero, D.; Arevalo-Martin, A.; Paniagua-Torija, B.; Sierra-Palomares, Y.; Molina-Holgado, E. A Cell Population That Strongly Expresses the CB1 Cannabinoid Receptor in the Ependyma of the Rat Spinal Cord. J. Comp. Neurol. 2013, 1, 233–251. [Google Scholar] [CrossRef]
- Vigh, B.; Manzano e Silva, M.J.; Frank, C.L.; Vincze, C.; Czirok, S.J.; Szabo, A.; Lukats, A.; Szel, A. The System of Cerebrospinal Fluid-Contacting Neurons. Its Supposed Role in the Nonsynaptic Signal Transmission of the Brain. Histol Histopathol 2004, 19, 607–628. [Google Scholar]
- Orts-Del’Immagine, A.; Trouslard, J.; Airault, C.; Hugnot, J.-P.; Cordier, B.; Doan, T.; Kastner, A.; Wanaverbecq, N. Postnatal Maturation of Mouse Medullo-Spinal Cerebrospinal Fluid-Contacting Neurons. Neuroscience 2017, 343, 39–54. [Google Scholar] [CrossRef]
- Ren, Y.; Ao, Y.; O’Shea, T.M.; Burda, J.E.; Bernstein, A.M.; Brumm, A.J.; Muthusamy, N.; Ghashghaei, H.T.; Carmichael, S.T.; Cheng, L.; et al. Ependymal Cell Contribution to Scar Formation after Spinal Cord Injury Is Minimal, Local and Dependent on Direct Ependymal Injury. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Maeda, Y.; Ma, J.; Delgado, M.; Sohn, J.; Miers, L.; Ko, E.M.; Bannerman, P.; Xu, J.; Wang, Y.; et al. Macroglial Plasticity and the Origins of Reactive Astroglia in Experimental Autoimmune Encephalomyelitis. J. Neurosci. 2011, 31, 11914–11928. [Google Scholar] [CrossRef] [Green Version]
- Barnabé-Heider, F.; Göritz, C.; Sabelström, H.; Takebayashi, H.; Pfrieger, F.W.; Meletis, K.; Frisén, J. Origin of New Glial Cells in Intact and Injured Adult Spinal Cord. Cell Stem Cell 2010, 7, 470–482. [Google Scholar] [CrossRef] [Green Version]
- Llorens-Bobadilla, E.; Chell, J.M.; Le Merre, P.; Wu, Y.; Zamboni, M.; Bergenstråhle, J.; Stenudd, M.; Sopova, E.; Lundeberg, J.; Shupliakov, O.; et al. A Latent Lineage Potential in Resident Neural Stem Cells Enables Spinal Cord Repair. Science 2020, 370, eabb8795. [Google Scholar] [CrossRef]
- Horner, P.J.; Power, A.E.; Kempermann, G.; Kuhn, H.G.; Palmer, T.D.; Winkler, J.; Thal, L.J.; Gage, F.H. Proliferation and Differentiation of Progenitor Cells throughout the Intact Adult Rat Spinal Cord. J. Neurosci. 2000, 20, 2218–2228. [Google Scholar] [CrossRef] [PubMed]
- Sabourin, J.-C.; Ackema, K.B.; Ohayon, D.; Guichet, P.-O.; Perrin, F.E.; Garces, A.; Ripoll, C.; Charité, J.; Simonneau, L.; Kettenmann, H.; et al. A Mesenchymal-Like ZEB1+ Niche Harbors Dorsal Radial Glial Fibrillary Acidic Protein-Positive Stem Cells in the Spinal Cord. Stem Cells 2009, 27, 2722–2733. [Google Scholar] [CrossRef]
- Nelson, C.M.; Lennon, V.A.; Lee, H.; Krug, R.G.; Kamalova, A.; Madigan, N.N.; Clark, K.J.; Windebank, A.J.; Henley, J.R. Glucocorticoids Target Ependymal Glia and Inhibit Repair of the Injured Spinal Cord. Front. Cell Dev. Biol. 2019, 7, 56. [Google Scholar] [CrossRef]
- Gonzalez-Fernandez, C.; Arevalo-Martin, A.; Paniagua-Torija, B.; Ferrer, I.; Rodriguez, F.J.; Garcia-Ovejero, D. Wnts Are Expressed in the Ependymal Region of the Adult Spinal Cord. Mol. Neurobiol. 2017, 54, 6342–6355. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ovejero, D.; Arevalo-Martin, A.; Paniagua-Torija, B.; Florensa-Vila, J.; Ferrer, I.; Grassner, L.; Molina-Holgado, E. The Ependymal Region of the Adult Human Spinal Cord Differs from Other Species and Shows Ependymoma-like Features. Brain 2015, 138, 1583–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marichal, N.; García, G.; Radmilovich, M.; Trujillo-Cenóz, O.; Russo, R.E. Spatial Domains of Progenitor-Like Cells and Functional Complexity of a Stem Cell Niche in the Neonatal Rat Spinal Cord. Stem Cells 2012, 30, 2020–2031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ševc, J.; Daxnerová, Z.; Haňová, V.; Koval’, J. Novel Observations on the Origin of Ependymal Cells in the Ventricular Zone of the Rat Spinal Cord. Acta Histochem. 2011, 113, 156–162. [Google Scholar] [CrossRef]
- Sevc, J.; Matiasova, A.; Kutna, V.; Daxnerova, Z. Evidence That the Central Canal Lining of the Spinal Cord Contributes to Oligodendrogenesis during Postnatal Development and Adulthood in Intact Rats. J. Comp. Neurol. 2014, 522, 3194–3207. [Google Scholar] [CrossRef] [PubMed]
- Watson, C.; Sengul, G.; Tanaka, I.; Rusznak, Z.; Tokuno, H. The Spinal Cord of the Common Marmoset (Callithrix Jacchus). Neurosci. Res. 2015, 93, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, S.; Hein, S.; Yulis, R.; Delannoy, L.; Siegmund, I.; Rodríguez, E. Reissner’s Fiber and the Wall of the Central Canal in the Lumbo-Sacral Region of the Bovine Spinal Cord. Cell Tissue Res. 1985, 240, 649–662. [Google Scholar] [CrossRef]
- Badlangana, N.L.; Bhagwandin, A.; Fuxe, K.; Manger, P.R. Observations on the Giraffe Central Nervous System Related to the Corticospinal Tract, Motor Cortex and Spinal Cord: What Difference Does a Long Neck Make? Neuroscience 2007, 148, 522–534. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Hernandez, M.; Tadokoro, T.; Navarro, M.R.; Platoshyn, O.; Kobayashi, Y.; Marsala, S.; Miyanohara, A.; Juhas, S.; Juhasova, J.; Skalnikova, H.; et al. Spinal Subpial Delivery of AAV9 Enables Widespread Gene Silencing and Blocks Motoneuron Degeneration in ALS. Nat. Med. 2020, 26, 118–130. [Google Scholar] [CrossRef]
- Marín-García, P.; González-Soriano, J.; Martinez-Sainz, P.; Contreras-Rodríguez, J.; Del Corral-Gros, C.; Rodríguez-Veiga, E. Spinal Cord Central Canal of the German Shepherd Dog: Morphological, Histological, and Ultrastructural Considerations. J. Morphol. 1995, 224, 205–212. [Google Scholar] [CrossRef]
- Böhme, G. Formation of the Central Canal and Dorsal Glial Septum in the Spinal Cord of the Domestic Cat. J. Anat. 1988, 159, 37–47. [Google Scholar]
- Wong, J.; Hemley, S.; Jones, N.; Cheng, S.; Bilston, L.; Stoodley, M. Fluid Outflow in a Large-Animal Model of Posttraumatic Syringomyelia. Neurosurgery 2012, 71, 474–480. [Google Scholar] [CrossRef]
- Billig, I.; Foris, J.M.; Enquist, L.W.; Card, J.P.; Yates, B.J. Definition of Neuronal Circuitry Controlling the Activity of Phrenic and Abdominal Motoneurons in the Ferret Using Recombinant Strains of Pseudorabies Virus. J. Neurosci. 2000, 20, 7446–7454. [Google Scholar] [CrossRef] [Green Version]
- Künzle, H. Tectal and Related Target Areas of Spinal and Dorsal Column Nuclear Projections in Hedgehog Tenrecs. Somat. Mot. Res. 1993, 10, 339–353. [Google Scholar] [CrossRef]
- Sawyer, E.K.; Turner, E.C.; Kaas, J.H. Somatosensory Brainstem, Thalamus, and Cortex of the California Sea Lion (Zalophus Californianus). J. Comp. Neurol. 2016, 524, 1957–1975. [Google Scholar] [CrossRef] [Green Version]
- Duffield, M.S.; Phillips, J.I.; Vieira-Makings, E.; Van Der Westhuyzen, J.; Metz, J. Demyelinisation in the Spinal Cord of Vitamin B12 Deficient Fruit Bats. Comp. Biochem. Physiol. Part C Comp. Pharm. 1990, 96, 291–297. [Google Scholar] [CrossRef]
- Dromard, C.; Guillon, H.; Rigau, V.; Ripoll, C.; Sabourin, J.C.; Perrin, F.E.; Scamps, F.; Bozza, S.; Sabatier, P.; Lonjon, N.; et al. Adult Human Spinal Cord Harbors Neural Precursor Cells That Generate Neurons and Glial Cells In Vitro. J. Neurosci. Res. 2008, 1926, 1916–1926. [Google Scholar] [CrossRef]
- Sakakibara, A.; Aoki, E.; Hashizume, Y.; Mori, N.; Nakayama, A. Distribution of Nestin and Other Stem Cell-Related Molecules in Developing and Diseased Human Spinal Cord. Pathol. Int. 2007, 57, 358–368. [Google Scholar] [CrossRef] [PubMed]
- Lavezzi, A.M.; Corna, M.F.; Matturri, L. Ependymal Alterations in Sudden Intrauterine Unexplained Death and Sudden Infant Death Syndrome: Possible Primary Consequence of Prenatal Exposure to Cigarette Smoking. Development 2010, 19, 5–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Lenhossek, M. Der Feinere Bau Des Nervenssystems Im Lichte Neuerer Forschung; Gustav Fischer: Berlin, Germany, 1895. [Google Scholar]
- Yasui, K.; Hashizume, Y.; Yoshida, M.; Kameyama, T.; Sobue, G. Age-Related Morphologic Changes of the Central Canal of the Human Spinal Cord Occluded Type. J. Neurol. Sci. 1999, 97, 253–259. [Google Scholar]
- Milhorat, T.H.; Kotzen, R.M.; Anzil, A.P. Stenosis of Central Canal of Spinal Cord in Man: Incidence and Pathological Findings in 232 Autopsy Cases. J. Neurosurg. 1994, 80, 716–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasantikul, V.; Netsky, M.G.; James, A.E., Jr. Relation of Age and Cerebral Ventricle Size to Central Canal in Man. Morphological Analysis. J. Neurosurg. 1979, 51, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Paniagua-Torija, B.; Norenberg, M.; Arevalo-Martin, A.; Carballosa-Gautam, M.M.; Campos-Martin, Y.; Molina-Holgado, E.; Garcia-Ovejero, D. Cells in the Adult Human Spinal Cord Ependymal Region Do Not Proliferate after Injury. J. Pathol. 2018, 246, 415–421. [Google Scholar] [CrossRef]
- Bronson, R.T.; Lane, P.W. Hydrocephalus with Hop Gait (Hyh): A New Mutation on Chromosome 7 in the Mouse. Dev. Brain Res. 1990, 54, 131–136. [Google Scholar] [CrossRef]
- Anyan, J.J.; Seney, M.L.; Holley, A.; Bengston, L.; Goldman, B.D.; Forger, N.G.; Holmes, M.M. Social Status and Sex Effects on Neural Morphology in Damaraland Mole-Rats, Fukomys Damarensis. Brain. Behav. Evol. 2011, 77, 291–298. [Google Scholar] [CrossRef] [Green Version]
- Peroulakis, M.E.; Goldman, B.; Forger, N.G. Perineal Muscles and Motoneurons Are Sexually Monomorphic in the Naked Mole-Rat (Heterocephalus Glaber). J. Neurobiol. 2002, 51, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Chae, T.H.; Kim, S.; Marz, K.E.; Hanson, P.I.; Walsh, C.A. The Hyh Mutation Uncovers Roles for ±Snap in Apical Protein Localization and Control of Neural Cell Fate. Nat. Genet. 2004, 36, 264–270. [Google Scholar] [CrossRef]
- Hong, H.K.; Chakravarti, A.; Takahashi, J.S. The Gene for Soluble N-Ethylmaleimide Sensitive Factor Attachment Protein α Is Mutated in Hydrocephaly with Hop Gait (Hyh) Mice. Proc. Natl. Acad. Sci. USA 2004, 101, 1748–1753. [Google Scholar] [CrossRef] [Green Version]
- Jiménez, A.J.; Tomé, M.; Páez, P.; Wagner, C.; Rodríguez, S.; Fernández-Llebrez, P.; Rodríguez, E.M.; Pérez-Fígares, J.M. A Programmed Ependymal Denudation Precedes Congenital Hydrocephalus in the Hyh Mutant Mouse. J. Neuropathol. Exp. Neurol. 2001, 60, 1105–1119. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.D.; Rossiter, S.J.; Faulkes, C.G. The Relationship between Individual Phenotype and the Division of Labour in Naked Mole-Rats: It’s Complicated. PeerJ 2020, 8, e9891. [Google Scholar] [CrossRef]
- Faulkes, C.G.; Abbott, D.H.; Liddell, C.E.; George, L.M.; Jarvis, J.U.M. Hormonal and behavioral aspects of reproductive suppression in female naked mole-rats. In The Biology of the Naked Mole-Rat; Sherman, P.W., Jarvis, J.U.M., Alexander, R.D., Eds.; Princeton University Press: Princeton, NJ, USA, 1991. [Google Scholar]
- Bátiz, L.F.; Roales-Buján, R.; Rodríguez-Pérez, L.M.; Matas, I.M.; Páez, P.; Roque, M.; Jiménez, A.J.; Ramos, C.; Pérez-Fígares, J.M. A Simple PCR-Based Genotyping Method for M105I Mutation of Alpha-SNAP Enhances the Study of Early Pathological Changes in Hyh Phenotype. Mol. Cell. Probes 2009, 23, 281–290. [Google Scholar] [CrossRef]
- Rivadulla, C.; Aguilar, J.; Coletti, M.; Aguila, J.; Prieto, S.; Cudeiro, J. Static Magnetic Fields Reduce Epileptiform Activity in Anesthetized Rat and Monkey. Sci. Rep. 2018, 8, 15985. [Google Scholar] [CrossRef]
- Namimatsu, S.; Ghazizadeh, M.; Sugisaki, Y. Reversing the Effects of Formalin Fixation with Citraconic Anhydride and Heat: A Universal Antigen Retrieval Method. J. Histochem Cytochem 2005, 53, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Alelu-Paz, R.; Iturrieta-Zuazo, I.; Byne, W.; Haroutunian, V.; Garcia-Villanueva, M.; Rabano, A.; Garcia-Amado, M.; Prensa, L.; Gimenez-Amaya, J.M. A New Antigen Retrieval Technique for Human Brain Tissue. PLoS ONE 2008, 3, e3378. [Google Scholar] [CrossRef]
- Venable, J.H.; Coggeshall, R. A Simplified Lead Citrate Stain for Use in Electron Microscopy. J. Cell Biol 1965, 25, 407–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, J.L. Further Researches on the Grey Substance of the Spinal Cord. Philos. Trans. R. Soc. Lond. 1859, 149, 437–467. [Google Scholar]
- Weigert, K. Beiträge Zur Kenntnis Der Normalen Menschlichen Neuroglia; Diesterweg: Frankfurt, Germany, 1895. [Google Scholar]
- Cornil Mosinger, M.L. Sur Les Processus Prolifereratifs de l’ependyme Medullaire (Rapports Avec Les Tumeurs Intramedullaires et La Syringomyelie). Rev. Neurol. 1933, 1, 749–754. [Google Scholar]
- Milhorat, T.H.; Capocelli Jr., A.L.; Anzil, A.P.; Kotzen, R.M.; Milhorat, R.H. Pathological Basis of Spinal Cord Cavitation in Syringomyelia: Analysis of 105 Autopsy Cases. J. Neurosurg. 1995, 82, 802–812. [Google Scholar] [CrossRef]
- Paniagua-Torija, B.; Arevalo-Martin, A.; Ferrer, I.; Molina-Holgado, E.; Garcia-Ovejero, D. CB1 Cannabinoid Receptor Enrichment in the Ependymal Region of the Adult Human Spinal Cord. Sci. Rep. 2015, 5, 17745. [Google Scholar] [CrossRef]
- Lowe, R.; Danson, A.F.; Rakyan, V.K.; Yildizoglu, S.; Saldmann, F.; Viltard, M.; Friedlander, G.; Faulkes, C.G. DNA Methylation Clocks as a Predictor for Ageing and Age Estimation in Naked Mole-Rats, Heterocephalus Glaber. Aging 2020, 12, 4394–4406. [Google Scholar] [CrossRef]
- Omerbašić, D.; Smith, E.S.J.; Moroni, M.; Homfeld, J.; Eigenbrod, O.; Bennett, N.C.; Reznick, J.; Faulkes, C.G.; Selbach, M.; Lewin, G.R. Hypofunctional TrkA Accounts for the Absence of Pain Sensitization in the African Naked Mole-Rat. Cell Rep. 2016, 17, 748–758. [Google Scholar] [CrossRef] [Green Version]
- Faulkes, C.G.; Bennett, N.C. Family Values: Group Dynamics and Social Control of Reproduction in African Mole-Rats. Trends Ecol. Evol. 2001, 16, 184–190. [Google Scholar] [CrossRef]
- Browe, B.M.; Vice, E.N.; Park, T.J. Naked Mole-Rats: Blind, Naked, and Feeling No Pain. Anat. Rec. 2020, 303, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Hepburn, D.; Waterston, D.; Turner, W.E.S. A Comparative Study of the Grey and White Matter of the Motor Cell Groups, and of the Spinal Accessory Nerve, in the Spinal Cord of the Porpoise (Phocoena Communis). Proc. R. Soc. Lond. 1903, 71, 444–445. [Google Scholar] [CrossRef]
- Hepburn, D.; Waterston, D. A Comparative Study of the Grey and White Matter, of the Motor-Cell Groups, and of the Spinal Accessory Nerve, in the Spinal Cord of the Porpoise (Phocoena Communis): Part II. J. Anat. Physiol. 1904, 38, 295–311. [Google Scholar]
- Seki, Y. Observations on the Spinal Cord of the Right Whale. Sci. Rep. Whale Res. Inst. Tokyo 1958, 13, 231–251. [Google Scholar]
- Bombardi, C.; Grandis, A.; Gardini, A.; Cozzi, B. Nitrergic Neurons in the Spinal Cord of the Bottlenose Dolphin (Tursiops Truncatus). Anat. Rec. 2013, 296, 1603–1614. [Google Scholar] [CrossRef]
- Bátiz, L.F.; Páez, P.; Jiménez, A.J.; Rodríguez, S.; Wagner, C.; Pérez-Fígares, J.M.; Rodríguez, E.M. Heterogeneous Expression of Hydrocephalic Phenotype in the Hyh Mice Carrying a Point Mutation in α-SNAP. Neurobiol. Dis. 2006, 23, 152–168. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.; Batiz, L.F.; Rodríguez, S.; Jiménez, A.J.; Páez, P.; Tomé, M.; Pérez-Fígares, J.M.; Rodríguez, E.M. Cellular Mechanisms Involved in the Stenosis and Obliteration of the Cerebral Aqueduct of Hyh Mutant Mice Developing Congenital Hydrocephalus. J. Neuropathol. Exp. Neurol. 2003, 62, 1019–1040. [Google Scholar] [CrossRef] [Green Version]
- Skarlatou, S.; Hérent, C.; Toscano, E.; Mendes, C.S.; Bouvier, J.; Zampieri, N. Afadin Signaling at the Spinal Neuroepithelium Regulates Central Canal Formation and Gait Selection. Cell Rep. 2020, 31, 107741. [Google Scholar] [CrossRef]
- Vogel, J.K.; Weider, M.; Engler, L.A.; Hillgärtner, S.; Schmitt, C.; Hermans-Borgmeyer, I.; Wegner, M. Sox9 Overexpression Exerts Multiple Stage-Dependent Effects on Mouse Spinal Cord Development. Glia 2020, 68, 932–946. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Liu, B.; Yang, X.; Yue, X.; Diao, L.; Wang, J.; Chang, J. Genetic Deletion of Rnd3 Results in Aqueductal Stenosis Leading to Hydrocephalus through Up-Regulation of Notch Signaling. Proc. Natl. Acad. Sci. USA 2013, 110, 8236–8241. [Google Scholar] [CrossRef] [Green Version]
- Abdi, K.; Lai, C.-H.; Paez-Gonzalez, P.; Lay, M.; Pyun, J.; Kuo, C.T. Uncovering Inherent Cellular Plasticity of Multiciliated Ependyma Leading to Ventricular Wall Transformation and Hydrocephalus. Nat. Commun. 2018, 9, 1655. [Google Scholar] [CrossRef]
- Hayashi, K.; Iwasaki, Y.; Yanagi, K. Herpes Simplex Virus Type 1-Induced Hydrocephalus in Mice. J. Virol. 1986, 57, 942–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conrady, C.D.; Zheng, M.; van Rooijen, N.; Drevets, D.A.; Royer, D.; Alleman, A.; Carr, D.J.J. Microglia and a Functional Type I IFN Pathway Are Required To Counter HSV-1–Driven Brain Lateral Ventricle Enlargement and Encephalitis. J. Immunol. 2013, 190, 2807–2817. [Google Scholar] [CrossRef] [PubMed]
- Milhorat, T.H.; Kotzen, R.M. Stenosis of the Central Canal of the Spinal Cord Following Inoculation of Suckling Hamsters with Reovirus Type I. J. Neurosurg. 1994, 81, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Meletis, K.; Barnabe-Heider, F.; Carlen, M.; Evergren, E.; Tomilin, N.; Shupliakov, O.; Frisen, J. Spinal Cord Injury Reveals Multilineage Differentiation of Ependymal Cells. PLoS Biol. 2008, 6, e182. [Google Scholar] [CrossRef]
- O’Shea, T.M.; Burda, J.E.; Sofroniew, M.V. Cell Biology of Spinal Cord Injury and Repair. J. Clin. Investig. 2017, 127, 3259–3270. [Google Scholar] [CrossRef]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A Summary. Neuro. Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef]
- López-Novoa, J.M.; Nieto, M.A. Inflammation and EMT: An Alliance towards Organ Fibrosis and Cancer Progression. Embo Mol. Med. 2009, 1, 303–314. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Antin, P.; Berx, G.; Blanpain, C.; Brabletz, T.; Bronner, M.; Campbell, K.; Cano, A.; Casanova, J.; Christofori, G.; et al. Guidelines and Definitions for Research on Epithelial–Mesenchymal Transition. Nat. Rev. Mol. Cell Biol. 2020, 21, 341–352. [Google Scholar] [CrossRef] [Green Version]
- Nieto, M.A. The Ins and Outs of the Epithelial to Mesenchymal Transition in Health and Disease. Annu. Rev. Cell Dev. Biol. 2011, 27, 347–376. [Google Scholar] [CrossRef] [Green Version]
- Klatt Shaw, D.; Saraswathy, V.M.; Zhou, L.; McAdow, A.R.; Burris, B.; Butka, E.; Morris, S.A.; Dietmann, S.; Mokalled, M.H. Localized EMT Reprograms Glial Progenitors to Promote Spinal Cord Repair. Dev. Cell 2021, 56, 613–626. [Google Scholar] [CrossRef]
- Franco, D.L.; Mainez, J.; Vega, S.; Sancho, P.; Murillo, M.M.; de Frutos, C.A.; del Castillo, G.; López-Blau, C.; Fabregat, I.; Nieto, M.A. Snail1 Suppresses TGF-β-Induced Apoptosis and Is Sufficient to Trigger EMT in Hepatocytes. J. Cell Sci. 2010, 123, 3467–3477. [Google Scholar] [CrossRef] [Green Version]
- Cañizares, M.A.; Albors, A.R.; Singer, G.; Suttie, N.; Gorkic, M.; Felts, P.; Storey, K.G. Multiple Steps Characterise Ventricular Layer Attrition to Form the Ependymal Cell Lining of the Adult Mouse Spinal Cord Central Canal. J. Anat. 2020, 236, 334–350. [Google Scholar] [CrossRef]
- Tait, C.M.; Chinnaiya, K.; Manning, E.; Murtaza, M.; Ashton, J.P.; Furley, N.; Hill, C.J.; Alves, C.H.; Wijnholds, J.; Erdmann, K.S.; et al. Crumbs2 Mediates Ventricular Layer Remodelling to Form the Spinal Cord Central Canal. PLoS Biol. 2020, 18, e3000470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ševc, J.; Daxnerová, Z.; Miklošová, M. Role of Radial Glia in Transformation of the Primitive Lumen to the Central Canal in the Developing Rat Spinal Cord. Cell. Mol. Neurobiol. 2009, 29, 927–936. [Google Scholar] [CrossRef]
- Sun, A.X.; Londono, R.; Hudnall, M.L.; Tuan, R.S.; Lozito, T.P. Differences in Neural Stem Cell Identity and Differentiation Capacity Drive Divergent Regenerative Outcomes in Lizards and Salamanders. Proc. Natl. Acad. Sci. USA 2018, 115, 8256–8265. [Google Scholar] [CrossRef] [Green Version]
- Mothe, A.J.; Zahir, T.; Santaguida, C.; Cook, D.; Tator, C.H. Neural Stem/Progenitor Cells from the Adult Human Spinal Cord Are Multipotent and Self-Renewing and Differentiate after Transplantation. PLoS ONE 2011, 6, e27079. [Google Scholar] [CrossRef] [PubMed]
- Mamaeva, D.; Ripoll, C.; Bony, C.; Teigell, M.; Perrin, F.E.; Rothhut, B.; Bieche, I.; Lidereau, R.; Privat, A.; Rigau, V.; et al. Isolation of Mineralizing Nestin+ Nkx6.1+ Vascular Muscular Cells from the Adult Human Spinal Cord. BMC Neurosci. 2011, 12, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Autopsy Number | Cause of Death | Gender | Age | Canal Patency | Postmortem Delay |
|---|---|---|---|---|---|
| BC00659 | Diffuse anoxia/ischemia | Male | 48 | Partially patent | Unknown |
| BC00692 | Microhemorrhage areas in Brain | Male | 19 | Partially patent | Unknown |
| BC01495 | Brain Edema | Male | 14 | Patent | Unknown |
| BC01684 | Acute Hypoxia–ischemia | Male | 27 | Partially patent | Unknown |
| BC01736 | Small Vessel Disease | Male | 30 | Patent | Unknown |
| BC01775 | Acute Hypoxia–ischemia | Male | 45 | Partially patent | Unknown |
| BC01800 | Hepatic encephalopathy | Male | 56 | Partially patent | Unknown |
| A07/044 | Cardiopulmonary arrest | Male | 39 | Closed | 3 h 30 min |
| A07/067 | Refractory septic shock and cardiac arrest. Ischemic cardiopathy | Male | 47 | Closed | 4 h 55 min |
| A10/017 | Hepatic metastasis. Probable pancreatic neoplasia | Male | 52 | Closed | 3 h |
| Antibody | Host Species | Manufacturer | Catalogue Number | Dilution |
|---|---|---|---|---|
| Activated Notch1-NICD | Rabbit | Abcam | Ab8925 | 1:1000 |
| GFAP | Chicken | Aves Labs | #GFAP | 1:300 |
| GFAP | Rabbit | DakoCytomation | #Z 0334 | 1:2000 |
| Iba1 | Rabbit | WAKO | 019-19741 | 1:500 |
| Laminin | Chicken | SIGMA | GW20044F | 1:600 |
| Nestin | Rabbit | ACRIS | APO9573PU-M | 1:250 |
| Phospho-SMAD3 (S423–S425) | Mouse | Abcam, clone EP823Y | Ab52903 | 1:100 |
| S100β | Mouse | SIGMA-Aldrich, clone SH-B1 | S2532 | 1:300 |
| Snai1 | Rabbit | Acris | AP20370PU-N | 1:300 |
| Sox2 | Mouse | R&D Systems, clone 245610 | MAB2018 | 1:200 |
| Sox9 | Goat | R&D Systems | AF3075 | 1:1000 |
| TJP1 (ZO-1) | Mouse | Thermo, clone ZO1-1A12 | 33-9100 | 1:200 |
| Vimentin | Mouse | DAKO clone V9 | M0725 | 1:500 |
| Antibody | Supplier | Dilution |
|---|---|---|
| Donkey anti-mouse biotin | Jackson IR, #715-065-151 | 1:500 |
| Donkey anti-rabbit biotin | Jackson IR, #711-065-152 | 1:500 |
| Streptavidin-Alexa 488 | Jackson IR, #016-540-084 | 1:500 |
| Donkey anti-mouse IgG Alexa 488 | Invitrogen #A-21202 | 1:1000 |
| Donkey anti-rabbit IgG Alexa 555 | Invitrogen #A-31572 | 1:1000 |
| Donkey anti-chicken IgY Cy5 | Jackson IR, #703-175-155 | 1:500 |
| Donkey anti-goat IgG Alexa 633 | Invitrogen #A-21082 | 1:1000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torrillas de la Cal, A.; Paniagua-Torija, B.; Arevalo-Martin, A.; Faulkes, C.G.; Jiménez, A.J.; Ferrer, I.; Molina-Holgado, E.; Garcia-Ovejero, D. The Structure of the Spinal Cord Ependymal Region in Adult Humans Is a Distinctive Trait among Mammals. Cells 2021, 10, 2235. https://doi.org/10.3390/cells10092235
Torrillas de la Cal A, Paniagua-Torija B, Arevalo-Martin A, Faulkes CG, Jiménez AJ, Ferrer I, Molina-Holgado E, Garcia-Ovejero D. The Structure of the Spinal Cord Ependymal Region in Adult Humans Is a Distinctive Trait among Mammals. Cells. 2021; 10(9):2235. https://doi.org/10.3390/cells10092235
Chicago/Turabian StyleTorrillas de la Cal, Alejandro, Beatriz Paniagua-Torija, Angel Arevalo-Martin, Christopher Guy Faulkes, Antonio Jesús Jiménez, Isidre Ferrer, Eduardo Molina-Holgado, and Daniel Garcia-Ovejero. 2021. "The Structure of the Spinal Cord Ependymal Region in Adult Humans Is a Distinctive Trait among Mammals" Cells 10, no. 9: 2235. https://doi.org/10.3390/cells10092235