
The Arbuscular Mycorrhizal Fungus Glomus viscosum Improves the Tolerance to Verticillium Wilt in Artichoke by Modulating the Antioxidant Defense Systems


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material and Sampling
2.3. Source of V. dahliae Isolates
2.4. Disease Assessments
2.5. Determination of ASC and GSH Pool Contents
2.6. Enzyme Assays
2.7. Electrophoretic Analyses
2.8. Lipid Peroxidation Analysis and H2O2 Content
2.9. Statistical Analysis
3. Results
3.1. Disease Assessments
3.2. Ascorbate and Glutathione Pool Content
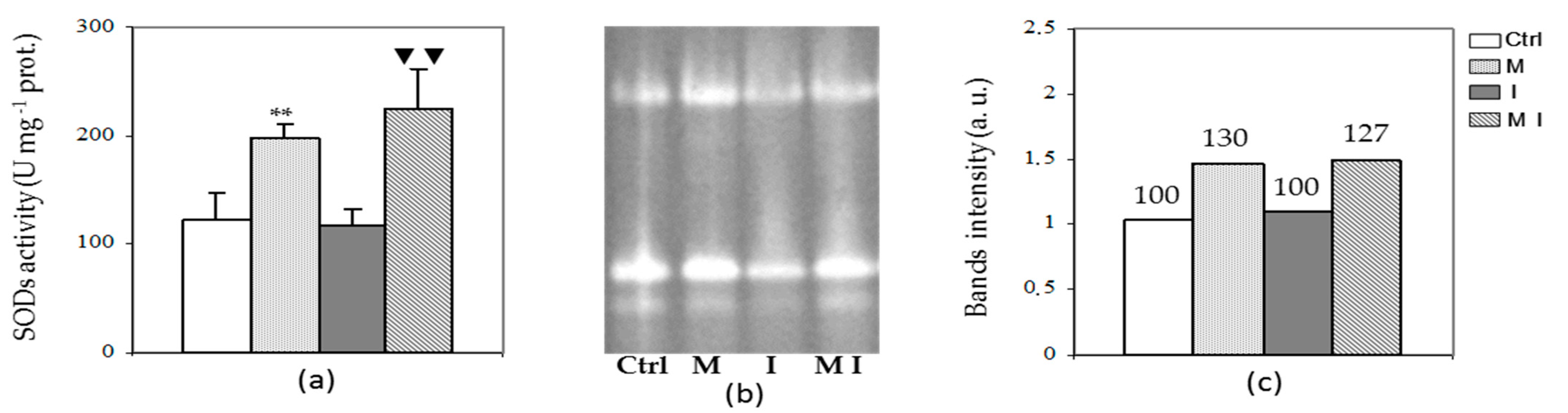
3.3. Antioxidant Enzyme Assays
3.4. H2O2 Content and Lipid Peroxidation Assay
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lattanzio, V.; Paul, A.K.; Lansalta, V.; Cardinali, A. Globe artichoke: A functional food and source of nutraceutical ingredients. J. Funct. Foods 2009, 1, 131–144. [Google Scholar] [CrossRef]
- Lombardo, S.; Pandino, G.; Ierna, A.; Mauromicale, G. Variation of polyphenols in a germplasm collection of globe artichoke. Food Res. Int. 2012, 46, 544–551. [Google Scholar] [CrossRef]
- Salem, M.B.; Affes, H.; Ksouda, K.; Dhouibi, R.; Sahnoun, Z.; Hammami, S.; Zeghal, K.M. Pharmacological studies of artichoke leaf extract and their health benefits. Plant Foods Hum. Nutr. 2015, 70, 441–453. [Google Scholar] [CrossRef]
- Fintelman, V. Therapeutic profile and mechanism of action of artichoke leaf extract: Hypolipemic, antioxidant, hepatoprotective and choleretic properties. Phytomedica 1996, 201, 50–60. [Google Scholar]
- Liorach, R.; Espin, J.C.; Tomas-Barberan, F.A.; Ferreres, F. Artichoke byproducts as a potential source of health-promoting antioxidant phenolics. J. Agric. Food Chem. 2002, 50, 58–64. [Google Scholar]
- Garbetta, A.; Capotorto, I.; Cardinali, A.; D’Antuono, I.; Linsalata, V.; Pizzi, F.; Minervini, F. Antioxidant activity induced by main polyphenols present in edible artichoke heads: Influence of in vitro gastro-intestinal digestion. J. Funct. Foods 2014, 10, 456–464. [Google Scholar] [CrossRef]
- Behara, Y.B.; Pharm, B. Pharmacological studies on artichoke leaf extract –an edible herb of Mediterranean origin. J. Pharmaceut. Biomed. Sci. 2011, 15, 111–600. [Google Scholar]
- Mohammad, A.; Sahar, N.; Effat, B. Effet of Cynara scolymus on fasting blood sugar of rat. Int. J. Pharm. Ind. Res. 2013, 3, 180–182. [Google Scholar]
- Gebhardt, R. Antioxidative and protective properties of extracts from leaves of the artichoke (Cynara scolymus L.) against hydroperoxide-induced oxidative stress in cultured rat hepatocytes. Toxicol. Appl. Pharmacol. 1997, 144, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Colla, G.; Graziani, G.; Ritieni, A.; Cardarelli, M.; De Pascale, S. Phenolic composition, antioxidant activity and mineral profile in two seed-propagated artichoke cultivars as affected by microbial inoculants and planting time. Food Chem. 2017, 234, 10–19. [Google Scholar] [CrossRef]
- Ancora, G. Globe artichoke (Cynara scolymus L.). In Biotechnology in Agriculture and Forestry, 2. Crops; Bajai, Y.P.S., Ed.; Springer: Berlin, Germany, 1986; pp. 471–484. [Google Scholar] [CrossRef]
- Grabowska, A.; Caruso, G.; Mehrafarin, A.; Kalisz, A.; Gruszecki, R.; Kunicki, E.; Sękara, A. Application of modern agronomic and biotechnological strategies to valorise worldwide globe artichoke (Cynara cardunculus L.) potential-an analytical overview. Ital. J. Agron. 2018, 13, 279–289. [Google Scholar] [CrossRef]
- Bhat, R.G.; Subbarao, K.V.; Bari, M.A. First report of Verticillium dahliae causing artichoke wilt in California. Plant Dis. 1999, 83, 782. [Google Scholar] [CrossRef]
- Berbegal, M.; Ortega, A.; García-Jiménez, J.; Armengol, J. Inoculum density-disease development relationship in Verticillium wilt of artichoke caused by Verticillium dahliae. Plant Dis. 2007, 91, 1131–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirulli, M.; Bubici, G.; Amenduni, M.; Armengol, J.; Berbegal, M.; Jiménez-Gasco, M.D.M.; Jiménez-Díaz, R.M. Verticillium wilt: A threat to artichoke production. Plant Dis. 2010, 94, 1176–1187. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Plant Health (PLH). Scientific opinion on the pest categorisation of Verticillium dahliae Kleb. EFSA J. 2014, 12, 3928. [Google Scholar]
- Fradin, E.F.; Thomma, B.P. Physiology and molecular aspects of Verticillium wilt diseases caused by V. dahliae and V. albo-atrum. Mol. Plant Pathol. 2006, 7, 71–86. [Google Scholar] [CrossRef]
- Klosterman, S.J.; Atallah, Z.K.; Vallad, G.E.; Subbarao, K.V. Diversity, pathogenicity and management of Verticillium species. Annu. Rev. Phytopathol. 2009, 47, 39–62. [Google Scholar] [CrossRef] [Green Version]
- Cirulli, M.; Ciccarese, F.; Amenduni, M. Evaluation of Italian Clones of Artichoke for Resistance to Verticillium dahliae. Plant Dis. 1994, 78, 680–682. [Google Scholar] [CrossRef]
- Pegg, G.F.; Brady, B.L. Verticillium Wilts; Cromwell Press: Trowbridge, UK, 2002. [Google Scholar] [CrossRef]
- Puri, K.D.; Hu, X.; Gurung, S.; Short, D.; Sandoya Miranda, G.V.; Schild, M.; Zhang, Y.; Zhao, J.; Anchieta, A.; Klosterman, S.J.; et al. Verticillium klebahnii and V. isaacii Isolates Exhibit Host-dependent Biological Control of Verticillium Wilt Caused by V. dahliae. Phytofrontiers 2021. [Google Scholar] [CrossRef]
- Villarino, M.; Larena, I.; Melgarejo, P.; De Cal, A. Effect of chemical alternatives to methyl bromide on soil-borne disease incidence and fungal populations in Spanish strawberry nurseries: A long-term study. Pest Manag. Sci. 2021, 77, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Bubici, G.; Marsico, A.D.; Gaber, L.; Tsror, L. Evaluation of thiophanate-methyl in controlling Verticillium wilt of potato and artichoke. Crop Prot. 2019, 119, 1–8. [Google Scholar] [CrossRef]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of arbuscular mycorrhizal fungi in plant growth regulation: Implications in abiotic stress tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, A.; Kadian, N.; Tanwar, A.; Yadav, A.; Gupta, K.K. Role of arbuscular mycorrhizal fungi (AMF) in global sustainable development. J. Nat. Appl. Sci. 2011, 3, 340–351. [Google Scholar] [CrossRef]
- Kapoor, R.; Singh, N. Arbuscular mycorrhiza and reactive oxygen species. In Arbuscular Mycorrhizas and Stress Tolerance of Plants; Springer: Berlin/Heidelberg, Germany, 2017; pp. 225–243. [Google Scholar] [CrossRef]
- Avio, L.; Maggini, R.; Ujvári, G.; Incrocci, L.; Giovannetti, M.; Turrini, A. Phenolics content and antioxidant activity in the leaves of two artichoke cultivars are differentially affected by six mycorrhizal symbionts. Sci. Hortic. 2020, 264, 109153. [Google Scholar] [CrossRef]
- Wu, Q.S.; Zou, Y.N.; Xia, R.X. Effects of water stress and arbuscular mycorrhizal fungi on reactive oxygen metabolism and antioxidant production by citrus (Citrus tangerine) roots. Eur. J. Soil Biol. 2006, 42, 166–172. [Google Scholar] [CrossRef]
- Wu, Q.S.; Zou, Y.N. Mycorrhiza has a direct effect on reactive oxygen metabolism of drought-stressed citrus. Plant Soil Environ. 2009, 55, 436–442. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.N.; Wu, Q.S.; Kuča, K. Unravelling the role of arbuscular mycorrhizal fungi in mitigating the oxidative burst of plants under drought stress. Plant Biol. J. 2020, 23, 50–57. [Google Scholar] [CrossRef]
- Boutaj, H.; Chakhchar, A.; Meddich, A.; Wahbi, S.; El Alaoui-Talibi, Z.; Douira, A.; Filali-Maltouf, A.; El Modafar, C. Bioprotection of olive tree from Verticillium wilt by autochthonous endomycorrhizal fungi. J. Plant Dis. Prot. 2020, 127, 349–357. [Google Scholar] [CrossRef]
- Boutaj, H.; Meddich, A.; Wahbi, S.; Moukhli, A.; El Alaoui-Talibi, Z.; Douira, A.; Filali-Maltouf, A.; El Modafar, C. Effect of arbuscular mycorrhizal fungi on verticillium wilt development of olive trees caused by Verticillium dahliae. Res. J. Biotechnol. 2019, 14, 79–88. [Google Scholar]
- Boutaj, H.; Meddich, A.; Chakhchar, A.; Wahbi, S.; El Alaoui-Talibi, Z.; Douira, A.; Filali-Maltouf, A.; El Modafar, C. Arbuscular mycorrhizal fungi improve mineral nutrition and tolerance of olive tree to Verticillium wilt. Arch. Phytopathol. Plant Prot. 2020, 53, 673–689. [Google Scholar] [CrossRef]
- Karagiannidis, N.; Bletsos, F.; Stavropoulos, N. Effect of Verticillium wilt (Verticillium dahliae Kleb.) and mycorrhiza (Glomus mosseae) on root colonization, growth and nutrient uptake in tomato and eggplant seedlings. Sci. Hortic. 2002, 94, 145–156. [Google Scholar] [CrossRef]
- Demír, S.; ¸Sensoy, S.; Ocak, E.; Tüfenkçi, ¸S.; Durak, E.D.; Erdinç, Ç.; Ünsal, H. Effects of arbuscular mycorrhizal fungus, humic acid, and whey on wilt disease caused by Verticillium dahliae Kleb. in three solanaceous crops. Turk. J. Agric. For. 2015, 39, 300–309. [Google Scholar] [CrossRef]
- Berg, G.; Zachow, C.; Lottmann, J.; Go, M.; Costa, R.; Icrobiol, A.P.P.L.E.N.M. Impact of Plant Species and Site on Rhizosphere-Associated Fungi Antagonistic to Verticillium dahliae Kleb. Appl. Environ. Microbiol. 2005, 71, 4203–4213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutaj, H.; Meddich, A.; Chakhchar, A.; Wahbi, S.; Alaoui-Talibi, Z.E.; Douira, A.; Filali-Maltouf, A.; El Modafar, C. Induction of early oxidative events in mycorrhizal olive tree in response to Verticillium wilt. Arch. Phytopathol. Plant Prot. 2021, 1–23. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Gara, L.; de Pinto, M.C.; Tommasi, F. The antioxidant systems vis-à-vis reactive oxygen species during plant–pathogen interaction. Plant Physiol. Biochem. 2003, 41, 863–870. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and Glutathione: Keeping Active Oxygen Under Control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Morone Fortunato, I.; Ruta, C.; Castrignanò, A.; Saccardo, F. The effect of mycorrhizal symbiosis on the development of micropropagated artichokes. Sci. Hortic. 2005, 106, 472–483. [Google Scholar] [CrossRef]
- Inderbitzin, P.; Bostock, R.M.; Davis, R.M.; Usami, T.; Platt, H.W.; Subbarao, K.V. Phylogenetics and taxonomy of the fungal vascular wilt pathogen Verticillium, with the descriptions of five new species. PLoS ONE 2011, 6, e28341. [Google Scholar] [CrossRef] [PubMed]
- Uppal, A.K.; El Hadrami, A.; Adam, L.R.; Tenuta, M.; Daayf, F. Biological control of potato Verticillium wilt under controlled and field conditions using selected bacterial antagonists and plant extracts. Biol. Control 2008, 44, 90–100. [Google Scholar] [CrossRef]
- Zhang, J.; Kirkham, B. Antioxidant responses to drought in sunflower and sorghum seedlings. New Phytol. 1996, 132, 361–373. [Google Scholar] [CrossRef]
- Mastropasqua, L.; Dipierro, N.; Paciolla, C. Effects of Darkness and Light Spectra on Nutrients and Pigments in Radish, Soybean, Mung Bean and Pumpkin Sprouts. Antioxidants 2020, 9, 558. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Paciolla, C.; Ippolito, M.P.; Logrieco, A.; Dipierro, N.; Mule, G.; Dipierro, S. A different trend of antioxidant defence responses makes tomato plants less susceptible to beauvericin than to T-2 mycotoxin phytotoxicity. Physiol. Mol. Plant Pathol. 2008, 72, 3–9. [Google Scholar] [CrossRef]
- Mastropasqua, L.; Borraccino, G.; Bianco, L.; Paciolla, C. Light qualities and dose influence ascorbate pool size in detached oat leaves. Plant Sci. 2012, 183, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Paciolla, C.; De Leonardis, S.; Zonno, M.C.; Vurro, M. Antioxidant response in Chenopodium album elicited by Ascochyta caulina mycoherbicide phytotoxins. Phytopathol. Mediterr. 2016, 346–354. [Google Scholar] [CrossRef]
- Lee, D.H.; Lee, C.B. Chilling stress-induced changes of antioxidant enzymes in the leaves of cucumber: In gel enzyme activity assays. Plant Sci. 2000, 159, 75–85. [Google Scholar] [CrossRef]
- Pedranzani, H.; Rodríguez-Rivera, M.; Gutiérrez, M.; Porcel, R.; Hause, B.; Ruiz-Lozano, J.M. Arbuscular mycorrhizal symbiosis regulates physiology and performance of Digitaria eriantha plants subjected to abiotic stresses by modulating antioxidant and jasmonate levels. Mycorrhiza 2016, 26, 141–152. [Google Scholar] [CrossRef] [Green Version]
- Pandino, G.; Lombardo, S.; Antonino, L.M.; Ruta, C.; Mauromicale, G. In vitro micropropagation and mycorrhizal treatment influences the polyphenols content profile of globe artichoke under field conditions. Food Res. Int. 2017, 99, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.M.; Arato, M.M.; Borghi, L.L.; Nouri, E.; Reinhardt, D. Beneficial services of arbuscular mycorrhizal fungi—From ecology to application. Front. Plant Sci. 2018, 4, 1270. [Google Scholar] [CrossRef] [PubMed]
- Gujre, N.; Soni, A.; Rangan, L.; Tsang, D.C.; Mitra, S. Sustainable improvement of soil health utilizing biochar and arbuscular mycorrhizal fungi: A review. Environ. Pollut. 2020, 115549. [Google Scholar] [CrossRef]
- Carillo, P.; Kyratzis, A.; Kyriacou, M.C.; Dell’Aversana, E.; Fusco, G.M.; Corrado, G.; Rouphael, Y. Biostimulatory Action of Arbuscular Mycorrhizal Fungi Enhances Productivity, Functional and Sensory Quality in ‘Piennolo del Vesuvio’ Cherry Tomato Landraces. Agronomy 2020, 10, 911. [Google Scholar] [CrossRef]
- Federico, N.S.; Carmona, M.; Balestrasse, K.; Chiocchio, V.; Giacometti, R.; Lavado, R.S. The arbuscular mycorrhizal fungus Rhizophagus intraradices reduces the root rot caused by Fusarium pseudograminearum in wheat. Rhizosphere 2021, 100369. [Google Scholar] [CrossRef]
- Garmendia, I.; Goicoechea, N.; Aguirreolea, J. Effectiveness of three Glomus species in protecting pepper (Capsicum annuum L.) against verticillium wilt. Biol. Control 2004, 31, 296–305. [Google Scholar] [CrossRef]
- Evelin, H.; Kapoor, R.; Giri, B. Arbuscular mycorrhizal fungi in alleviation of salt stress: A review. Ann. Bot. 2009, 104, 1263–1280. [Google Scholar] [CrossRef] [Green Version]
- Ceccarelli, N.; Curadi, M.; Martelloni, L.; Sbrana, C.; Picciarelli, P.; Giovannetti, M. Mycorrhizal colonization impacts on phenolic content and antioxidant properties of artichoke leaves and flower heads two years after field transplant. Plant Soil 2010, 335, 311–323. [Google Scholar] [CrossRef]
- Wang, J.; Zhai, L.; Ma, J.; Zhang, J.; Wang, G.G.; Liu, X.; Zhang, S.; Song, J.; Wu, Y. Comparative physiological mechanisms of arbuscular mycorrhizal fungi in mitigating salt-induced adverse effects on leaves and roots of Zelkova serrata. Mycorrhiza 2020, 30, 341–355. [Google Scholar] [CrossRef] [PubMed]
- Campanelli, A.; Ruta, C.; Tagarelli, A.; Morone-Fortunato, I.; De Mastro, G. Effectiveness of mycorrhizal fungi on globe artichoke (Cynara cardunculus L. var. scolymus) micropropagation. J. Plant Interact. 2014, 9, 100–106. [Google Scholar] [CrossRef]
- Zhang, Q.; Gao, X.; Ren, Y.; Ding, X.; Qiu, J.; Li, N.; Zeng, F.; Chu, Z. Improvement of Verticillium wilt resistance by applying arbuscular mycorrhizal fungi to a cotton variety with high symbiotic efficiency under field conditions. Int. J. Mol. Sci. 2018, 19, 241. [Google Scholar] [CrossRef] [Green Version]
- Ismail, Y.; McCormick, S.; Hijri, M. The arbuscular mycorrhizal fungus, Glomus irregulare, controls the mycotoxin production of Fusarium sambucinum in the pathogenesis of potato. FEMS Microbiol. Lett. 2013, 348, 46–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, P.; Guo, Y.; Li, Y.; Duan, T. Effects of dual inoculation of AMF and rhizobium on alfalfa (Medicago sativa) root rot caused by Microdochium tabacinum. Australas. Plant Pathol. 2018, 47, 195–203. [Google Scholar] [CrossRef]
- Campanella, V.; Miceli, C. Biological control of Fusarium wilt of Ustica landrace lentil. Crop Prot. 2021, 145, 105635. [Google Scholar] [CrossRef]
- Hernández, J.A.; Barba-Espín, G.; Diaz-Vivancos, P. Glutathione-mediated biotic stress tolerance in plants. In Glutathione in Plant Growth, Development, and Stress Tolerance; Hossain, M., Mostofa, M., Diaz-Vivancos, P., Burritt, D., Fujita, M., Tran, L.S., Eds.; Springer: Cham, Switzerland, 2017; pp. 309–319. [Google Scholar] [CrossRef]
- Bernal-Vicente, A.; Pascual, J.A.; Tittarelli, F.; Hernández, J.A.; Diaz-Vivancos, P. Trichoderma harzianum T-78 supplementation of compost stimulates the antioxidant defence system in melon plants. J. Sci. Food Agric. 2015, 95, 2208–2214. [Google Scholar] [CrossRef] [Green Version]
- Saroy, K.; Garg, N. Relative effectiveness of arbuscular mycorrhiza and polyamines in modulating ROS generation and ascorbate-glutathione cycle in Cajanus cajan under nickel stress. Environ. Sci. Pollut. Res. 2021, 1–18. [Google Scholar] [CrossRef]
- Nahiyan, A.S.M.; Matsubara, Y.I. Tolerance to Fusarium root rot and changes in antioxidative ability in mycorrhizal asparagus plants. HortScience 2012, 47, 356–360. [Google Scholar] [CrossRef] [Green Version]
- Kuzniak, E.; Skłodowska, M. Differential Implication of Glutathione, Glutathione Metabolizing Enzymes and Ascorbate in Tomato Resistance to Pseudomonas syringae. J. Phytopathol. 2004, 152, 529–536. [Google Scholar] [CrossRef]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.; Bakker, P.A. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, A.J.; Hell, R. Glutathione homeostasis and redox-regulation by sulfhydryl groups. Photosynth. Res. 2005, 86, 435–457. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Chaouch, S.; Han, Y.; Neukermans, J.; Marquez-Garcia, B.; Queval, G.; Foyer, C.H. Glutathione in plants: An integrated overview. Plant Cell Environ. 2012, 35, 454–484. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Anjum, N.A.; Hasanuzzaman, M.; Gill, R.; Trived, D.K.; Ahmad, I.; Pereira, E.; Tuteja, N. Glutathione reductase and glutathione: A boon in disguise for plant abiotic stress defense operations. Plant Physiol. Biochem. 2013, 70, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Gamble, P.; Burke, J.J. Effect of water stress on the chloroplast antioxidant system. Alteration in glutathione reductase activity. Plant Physiol. 1984, 76, 615–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alguacil, M.M.; Hernández, J.A.; Caravaca, F.; Portillo, B.; Roldán, A. Antioxidant enzyme activities in shoots from three mycorrhizal shrub species afforested in a degraded semi-arid soil. Physiol. Plant. 2003, 118, 562–570. [Google Scholar] [CrossRef]
- Kopriva, S.; Rennenberg, H. Control of sulphate assimilation and glutathione synthesis: Interaction with N and C metabolism. J. Exp. Bot. 2004, 55, 1831–1842. [Google Scholar] [CrossRef] [PubMed]
- Paciolla, C.; De Tullio, M.C.; Chiappetta, A.; Innocenti, A.M.; Bitonti, M.B.; Liso, R.; Arrigoni, O. Short- and Long-Term Effects of Dehydroascorbate in Lupinus albus and Allium cepa Roots. Plant Cell Physiol. 2001, 42, 857–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morell, S.; Follmann, H.; De Tullio, M.; Häberlein, I. Dehydroascorbate and dehydroascorbate reductase are phantom indicators of oxidative stress in plants. FEBS Lett. 1997, 414, 567–570. [Google Scholar] [CrossRef] [Green Version]
- Loi, M.; De Leonardis, S.; Mulè, G.; Logrieco, A.F.; Paciolla, C. A Novel and Potentially Multifaceted Dehydroascorbate Reductase Increasing the Antioxidant Systems is Induced by Beauvericin in Tomato. Antioxidants 2020, 9, 435. [Google Scholar] [CrossRef] [PubMed]
- Mittova, V.; Tal, M.; Volokita, M.; Guy, M. Up-regulation of the leaf mitochondrial and peroxisomal antioxidative systems in response to salt-induced oxidative stress in the wild salt-tolerant tomato species Lycopersicon pennellii. Plant Cell Environ. 2003, 26, 845–856. [Google Scholar] [CrossRef] [PubMed]
- Kavitha, K.; George, S.; Venkataraman, G.; Parida, A. A salt-inducible chloroplastic monodehydroascorbate reductase from halophyte Avicennia marina confers salt stress tolerance on transgenic plants. Biochimie 2010, 92, 1321–1329. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Xiangpeng, L.; Wenying, Z.; Jinggui, F. The Regulatory and Signaling Roles of Glutathione in Modulating Abiotic Stress Responses and Tolerance. In Glutathione in Plant Growth, Development, and Stress Tolerance; Hossain, M.A., Mostofa, M.G., Diaz-Vivancos, P., Burritt, D.J., Fujita, M., Tran, L.S.P., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 147–169. [Google Scholar] [CrossRef]
- Porcel, R.; Ruiz-lozano, J.M. Arbuscular mycorrhizal influence on leaf water potential, solute accumulation, and oxidative stress in soybean plants subjected to drought stress. J. Exp. Bot. 2004, 55, 1743–1750. [Google Scholar] [CrossRef] [Green Version]
- Garmendia, I.; Aguirreolea, J.; Goicoechea, N. Defence-related Enzymes in Pepper Roots During Interactions with Arbuscular Mycorrhizal Fungi and/or Verticillium dahliae. Biocontrol 2006, 51, 293. [Google Scholar] [CrossRef] [Green Version]
- Ortega, E.P.; Noval, B.M.L.; Coca, B.M.; Noval, W.T.L.; Carmona, A.M.; Hernández, A.; León, O. Induction of defense mechanisms in mycorrhized tomato plants against the attack of Oidiopsis taurica (Lev.) Salm. Cult. Trop. 2015, 36, 94–102. [Google Scholar]
- Asada, K. Ascorbate peroxidase–a hydrogen peroxide-scavenging enzyme in plants. Physiol. Plant. 1992, 85, 235–241. [Google Scholar] [CrossRef]
- Dat, J.; Vandenabeele, S.; Vranova, E.; Van Montagu, M.; Inzé, D.; Van Breusegem, F. Dual action of the active oxygen species during plant stress responses. Cell Mol. Life Sci. 2000, 57, 779–795. [Google Scholar] [CrossRef] [PubMed]
- Yamane, K.; Mitsuya, S.; Taniguchi, M.; Miyake, H. Transcription profiles of genes encoding catalase and ascorbate peroxidase in the rice leaf tissues under salinity. Plant Prod. Sci. 2010, 13, 164–168. [Google Scholar] [CrossRef]
- Estrada, B.; Aroca, R.; Maathuis, F.J.M.; Barea, J.M.; Ruiz-Lozano, J.M. Arbuscular mycorrhizal fungi native from a Mediterranean saline area enhance maize tolerance to salinity through improved ion homeostasis. Plant Cell Environ. 2013, 36, 1771–1782. [Google Scholar] [CrossRef]
- Polidoros, A.N.; Scandalios, J.G. Role of hydrogen peroxide and different classes of antioxidants in the regulation of catalase and glutathione S-transferase gene expression in maize (Zea mays L.). Physiol. Plant. 1999, 106, 112–120. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Disease Severity | Flower Heads for Plants | |
|---|---|---|---|
| Foliar Tissue | Vascular System | ||
| Ctrl | 0.0 c | 0.0 b | 13 |
| I | 3.9 a | 3.0 a | 4 |
| M | 0.0 c | 0.0 b | 16 |
| MI | 1.6 b | 2.7 a | 9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villani, A.; Tommasi, F.; Paciolla, C. The Arbuscular Mycorrhizal Fungus Glomus viscosum Improves the Tolerance to Verticillium Wilt in Artichoke by Modulating the Antioxidant Defense Systems. Cells 2021, 10, 1944. https://doi.org/10.3390/cells10081944
Villani A, Tommasi F, Paciolla C. The Arbuscular Mycorrhizal Fungus Glomus viscosum Improves the Tolerance to Verticillium Wilt in Artichoke by Modulating the Antioxidant Defense Systems. Cells. 2021; 10(8):1944. https://doi.org/10.3390/cells10081944
Chicago/Turabian StyleVillani, Alessandra, Franca Tommasi, and Costantino Paciolla. 2021. "The Arbuscular Mycorrhizal Fungus Glomus viscosum Improves the Tolerance to Verticillium Wilt in Artichoke by Modulating the Antioxidant Defense Systems" Cells 10, no. 8: 1944. https://doi.org/10.3390/cells10081944