
Identification of Cilia in Different Mouse Tissues


Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Genotyping
2.3. Preparation of Frozen Sections and Observation of Cilia
2.4. Immunofluorescence Staining
2.5. Quantitative Analysis of the Ciliary Number and Length
2.6. Time-Lapse Observations of Cilia in Living Nucleus Pulposus (NP) Tissue
3. Results
3.1. Cilia Are Present in Different Tissues of the Respiratory and Circulatory Systems
3.2. Primary Cilia Are Present in Different Organs of the Immune System
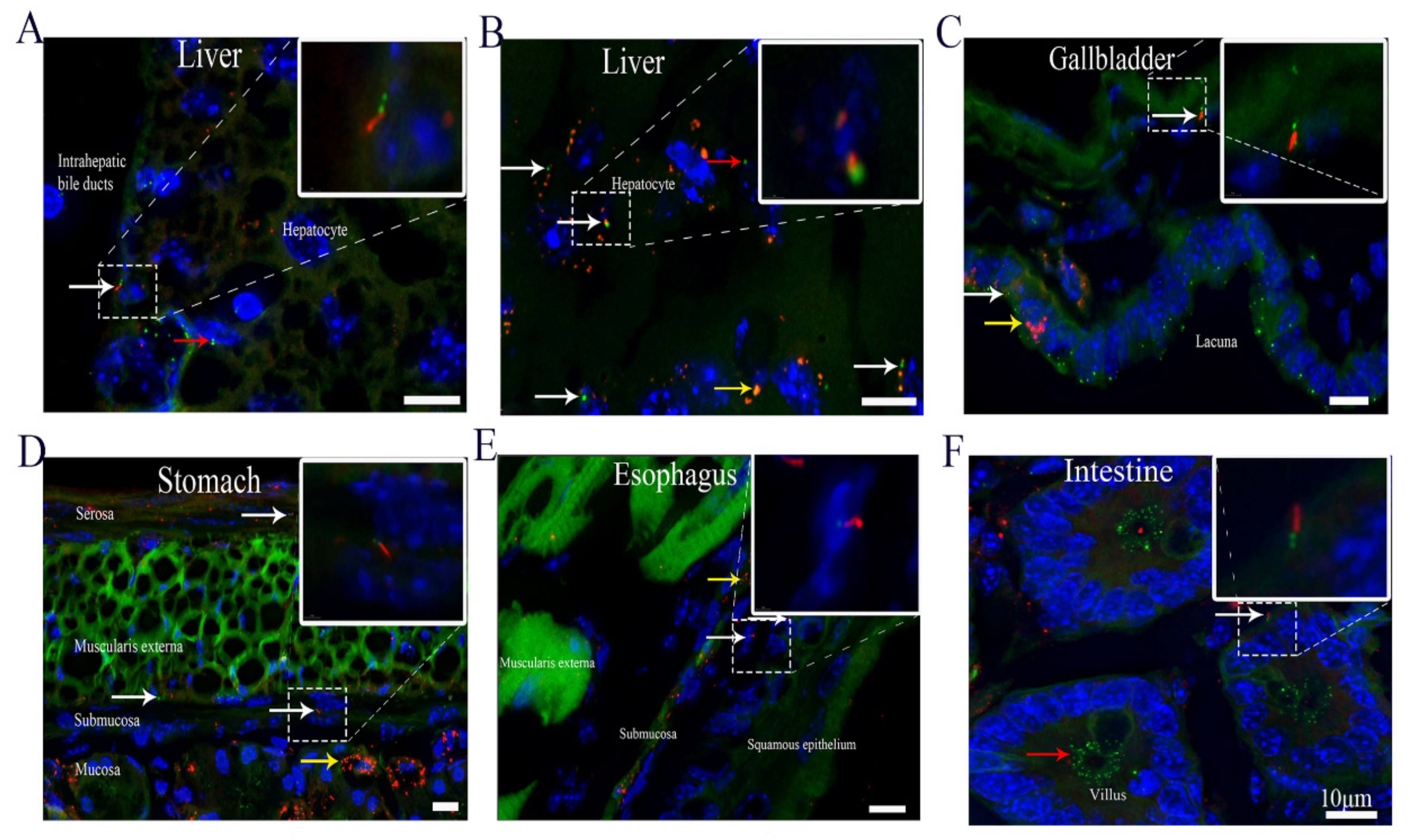
3.3. Cilia Are Present in Different Organs of the Digestive System, Including Hepatocytes
3.4. Cilia Are Present in Different Organs in the Urinary and Reproductive System
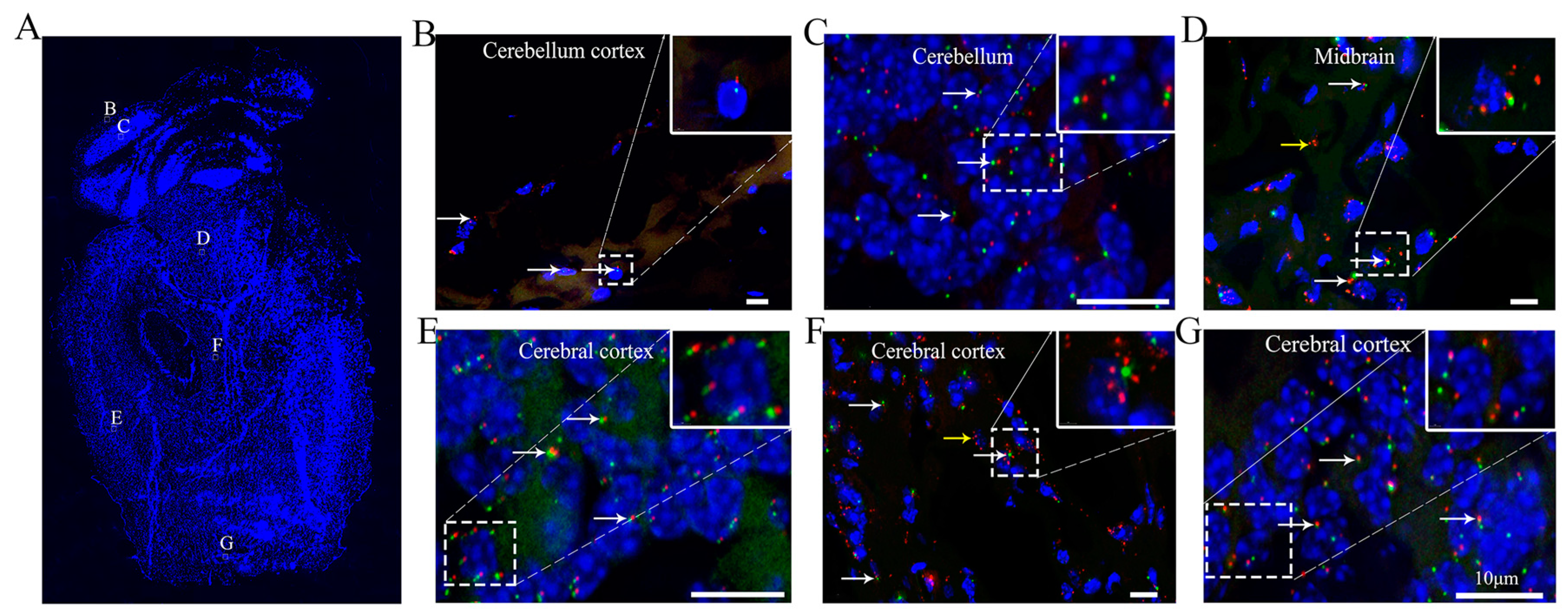
3.5. Ciliary Arrangement and Patterns in the Cerebral Cortex of the Brain
3.6. Cilia Are Present in Different Organs in the Musculoskeletal System
3.7. ARL13B+ Inclusions Are Present in the Thyroid and Islets of Langerhans
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bangs, F.K.; Schrode, N.; Hadjantonakis, A.K.; Anderson, K.V. Lineage specificity of primary cilia in the mouse embryo. Nat. Cell Biol. 2015, 17, 113–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Amsterdam, A.; Pazour, G.J.; Cole, D.G.; Miller, M.S.; Hopkins, N. A genetic screen in zebrafish identifies cilia genes as a principal cause of cystic kidney. Development 2004, 131, 4085–4093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz, F.; Burtscher, I.; Stauber, M.; Gossler, A.; Lickert, H. A novel Cre-inducible knock-in ARL13B-tRFP usion cilium reporter. Genesis 2017, 55. [Google Scholar] [CrossRef]
- Janssen, P.H. Fermentation of L-tartrate by a newly isolated gram-negative glycolytic bacterium. Antonie Van Leeuwenhoek 1991, 59, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Cantagrel, V.; Mariani, L.; Serre, V.; Lee, J.E.; Elkhartoufi, N.; de Lonlay, P.; Desguerre, I.; Munnich, A.; Boddaert, N.; et al. Identification of a novel ARL13B variant in a Joubert syndrome-affected patient with retinal impairment and obesity. Eur. J. Hum. Genet. 2015, 23, 621–627. [Google Scholar] [CrossRef] [Green Version]
- Higginbotham, H.; Bielas, S.; Tanaka, T.; Gleeson, J.G. Transgenic mouse line with green-fluorescent protein-labeled Centrin 2 allows visualization of the centrosome in living cells. Transgenic Res. 2004, 13, 155–164. [Google Scholar] [CrossRef]
- Hughes, J.W.; Cho, J.H.; Conway, H.E.; DiGruccio, M.R.; Ng, X.W.; Roseman, H.F.; Abreu, D.; Urano, F.; Piston, D.W. Primary cilia control glucose homeostasis via islet paracrine interactions. Proc. Natl. Acad. Sci. USA 2020, 117, 8912–8923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, F.; Schwartz, A.G.; Moore, E.R.; Sup, M.E.; Thomopoulos, S. Primary cilia as the nexus of biophysical and hedgehog signaling at the tendon enthesis. Sci. Adv. 2020, 6. [Google Scholar] [CrossRef]
- Dummer, A.; Poelma, C.; DeRuiter, M.C.; Goumans, M.J.; Hierck, B.P. Measuring the primary cilium length: Improved method for unbiased high-throughput analysis. Cilia 2016, 5, 7. [Google Scholar] [CrossRef] [Green Version]
- Konishi, S.; Gotoh, S.; Tateishi, K.; Yamamoto, Y.; Korogi, Y.; Nagasaki, T.; Matsumoto, H.; Muro, S.; Hirai, T.; Ito, I.; et al. Directed Induction of Functional Multi-ciliated Cells in Proximal Airway Epithelial Spheroids from Human Pluripotent Stem Cells. Stem Cell Rep. 2016, 6, 18–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruhlen, R.; Marberry, K. The chondrocyte primary cilium. Osteoarthr. Cartil. 2014, 22, 1071–1076. [Google Scholar] [CrossRef] [Green Version]
- Kaur, S.; McGlashan, S.R.; Ward, M.L. Evidence of primary cilia in the developing rat heart. Cilia 2018, 7, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Heiden, K.; Hierck, B.P.; Krams, R.; de Crom, R.; Cheng, C.; Baiker, M.; Pourquie, M.J.; Alkemade, F.E.; DeRuiter, M.C.; Gittenberger-de Groot, A.C.; et al. Endothelial primary cilia in areas of disturbed flow are at the base of atherosclerosis. Atherosclerosis 2008, 196, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Chaplin, D.D. Overview of the immune response. J. Allergy Clin. Immunol. 2010, 125, S3–S23. [Google Scholar] [CrossRef]
- Stephen, L.A.; ElMaghloob, Y.; McIlwraith, M.J.; Yelland, T.; Castro Sanchez, P.; Roda-Navarro, P.; Ismail, S. The Ciliary Machinery Is Repurposed for T Cell Immune Synapse Trafficking of LCK. Dev. Cell 2018, 47, 122–132.e124. [Google Scholar] [CrossRef] [Green Version]
- Mansini, A.P.; Peixoto, E.; Thelen, K.M.; Gaspari, C.; Jin, S.; Gradilone, S.A. The cholangiocyte primary cilium in health and disease. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Arterburn, L.M.; Zurlo, J.; Yager, J.D.; Overton, R.M.; Heifetz, A.H. A morphological study of differentiated hepatocytes in vitro. Hepatology 1995, 22, 175–187. [Google Scholar] [CrossRef]
- Saqui-Salces, M.; Dowdle, W.E.; Reiter, J.F.; Merchant, J.L. A high-fat diet regulates gastrin and acid secretion through primary cilia. FASEB J. 2012, 26, 3127–3139. [Google Scholar] [CrossRef] [Green Version]
- Said, H.M. Intestinal absorption of water-soluble vitamins in health and disease. Biochem. J. 2011, 437, 357–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoder, B.K. Role of primary cilia in the pathogenesis of polycystic kidney disease. J. Am. Soc. Nephrol. 2007, 18, 1381–1388. [Google Scholar] [CrossRef] [Green Version]
- Taub, M. Primary kidney cells. Methods Mol. Biol. 1997, 75, 153–161. [Google Scholar] [CrossRef]
- Du, E.; Lu, C.; Sheng, F.; Li, C.; Li, H.; Ding, N.; Chen, Y.; Zhang, T.; Yang, K.; Xu, Y. Analysis of potential genes associated with primary cilia in bladder cancer. Cancer Manag. Res. 2018, 10, 3047–3056. [Google Scholar] [CrossRef] [Green Version]
- Nygaard, M.B.; Almstrup, K.; Lindbaek, L.; Christensen, S.T.; Svingen, T. Cell context-specific expression of primary cilia in the human testis and ciliary coordination of Hedgehog signalling in mouse Leydig cells. Sci. Rep. 2015, 5, 10364. [Google Scholar] [CrossRef] [Green Version]
- Dores, C.; Alpaugh, W.; Su, L.; Biernaskie, J.; Dobrinski, I. Primary cilia on porcine testicular somatic cells and their role in hedgehog signaling and tubular morphogenesis in vitro. Cell Tissue Res. 2017, 368, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, J.L.; Schwark, H.D. Neuronal primary cilia: A review. Cell Biol. Int. 2004, 28, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Cao, J.; He, X.; Serra, R.; Qu, J.; Cao, X.; Yang, S. Ciliary IFT80 balances canonical versus non-canonical hedgehog signalling for osteoblast differentiation. Nat. Commun. 2016, 7, 11024. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Alharbi, M.; Graves, D.; Yang, S. IFT80 Is Required for Fracture Healing Through Controlling the Regulation of TGF-beta Signaling in Chondrocyte Differentiation and Function. J. Bone Miner. Res. 2020, 35, 571–582. [Google Scholar] [CrossRef]
- McCann, M.R.; Tamplin, O.J.; Rossant, J.; Seguin, C.A. Tracing notochord-derived cells using a Noto-cre mouse: Implications for intervertebral disc development. Dis. Models Mech. 2012, 5, 73–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirokawa, N.; Tanaka, Y.; Okada, Y.; Takeda, S. Nodal flow and the generation of left-right asymmetry. Cell 2006, 125, 33–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, M.; Shinohara, K.; Wang, J.; Ikeuchi, S.; Yoshiba, S.; Meno, C.; Nonaka, S.; Takada, S.; Hatta, K.; Wynshaw-Boris, A.; et al. Planar polarization of node cells determines the rotational axis of node cilia. Nat. Cell Biol. 2010, 12, 170–176. [Google Scholar] [CrossRef]
- Cai, M.; Li, J.; Yue, R.; Wang, Z.; Sun, Y. Glycosylation of DMP1 maintains cranial sutures in mice. J. Oral Rehabil. 2019. [Google Scholar] [CrossRef]
- Aughsteen, A.A. The ultrastructure of primary cilia in the endocrine and excretory duct cells of the pancreas of mice and rats. Eur. J. Morphol. 2001, 39, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Santos, J.M.; Utrilla, J.C.; Vazquez-Roman, V.; Villar-Rodriguez, J.L.; Gutierrez-Aviles, L.; Martin-Lacave, I. Primary Cilium in the Human Thyrocyte: Changes in Frequency and Length in Relation to the Functional Pathology of the Thyroid Gland. Thyroid 2019, 29, 595–606. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, S.M.; Watson, D.S.; Novoselova, T.V.; Romano, L.E.L.; King, P.J.; Bradshaw, T.Y.; Thompson, C.L.; Knight, M.M.; Sharp, T.V.; Barnes, M.R.; et al. Oncometabolite induced primary cilia loss in pheochromocytoma. Endocr. Relat. Cancer 2019, 26, 165–180. [Google Scholar] [CrossRef]
- Lodh, S.; O’Hare, E.A.; Zaghloul, N.A. Primary cilia in pancreatic development and disease. Birth Defects Res. Part C Embryo Today 2014, 102, 139–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.L.; Malone, T.R. Effects of running on intervertebral disc height. J. Orthop. Sports Phys. Ther. 1990, 12, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yang, S.; Han, L.; Mao, K.; Yang, S. Ciliary IFT80 is essential for intervertebral disc development and maintenance. FASEB J. 2020. [Google Scholar] [CrossRef] [Green Version]
- Mall, M.A. Role of cilia, mucus, and airway surface liquid in mucociliary dysfunction: Lessons from mouse models. J. Aerosol. Med. Pulm. Drug Deliv. 2008, 21, 13–24. [Google Scholar] [CrossRef]
- Faubel, R.; Westendorf, C.; Bodenschatz, E.; Eichele, G. Cilia-based flow network in the brain ventricles. Science 2016, 353, 176–178. [Google Scholar] [CrossRef] [PubMed]
- Mitchison, H.M.; Valente, E.M. Motile and non-motile cilia in human pathology: From function to phenotypes. J. Pathol. 2017, 241, 294–309. [Google Scholar] [CrossRef] [PubMed]
- Walczak-Sztulpa, J.; Posmyk, R.; Bukowska-Olech, E.M.; Wawrocka, A.; Jamsheer, A.; Oud, M.M.; Schmidts, M.; Arts, H.H.; Latos-Bielenska, A.; Wasilewska, A. Compound heterozygous IFT140 variants in two Polish families with Sensenbrenner syndrome and early onset end-stage renal disease. Orphanet J. Rare Dis. 2020, 15, 36. [Google Scholar] [CrossRef] [PubMed]
- Stinchcombe, J.C.; Salio, M.; Cerundolo, V.; Pende, D.; Arico, M.; Griffiths, G.M. Centriole polarisation to the immunological synapse directs secretion from cytolytic cells of both the innate and adaptive immune systems. BMC Biol. 2011, 9, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stinchcombe, J.C.; Majorovits, E.; Bossi, G.; Fuller, S.; Griffiths, G.M. Centrosome polarization delivers secretory granules to the immunological synapse. Nature 2006, 443, 462–465. [Google Scholar] [CrossRef]
- Larkins, C.E.; Aviles, G.D.; East, M.P.; Kahn, R.A.; Caspary, T. ARL13B regulates ciliogenesis and the dynamic localization of Shh signaling proteins. Mol. Biol. Cell 2011, 22, 4694–4703. [Google Scholar] [CrossRef]
- Bishop, G.A.; Berbari, N.F.; Lewis, J.; Mykytyn, K. Type III adenylyl cyclase localizes to primary cilia throughout the adult mouse brain. J. Comp. Neurol. 2007, 505, 562–571. [Google Scholar] [CrossRef]
- Youn, Y.H.; Han, Y.G. Primary Cilia in Brain Development and Diseases. Am. J. Pathol. 2018, 188, 11–22. [Google Scholar] [CrossRef]
- Mohammed, S.G.; Arjona, F.J.; Verschuren, E.H.J.; Bakey, Z.; Alkema, W.; van Hijum, S.; Schmidts, M.; Bindels, R.J.M.; Hoenderop, J.G.J. Primary cilia-regulated transcriptome in the renal collecting duct. FASEB J. 2018, 32, 3653–3668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volta, F.; Scerbo, M.J.; Seelig, A.; Wagner, R.; O’Brien, N.; Gerst, F.; Fritsche, A.; Haring, H.U.; Zeigerer, A.; Ullrich, S.; et al. Glucose homeostasis is regulated by pancreatic beta-cell cilia via endosomal EphA-processing. Nat. Commun. 2019, 10, 5686. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Allard, B.A.; Pottorf, T.S.; Wang, H.H.; Vivian, J.L.; Tran, P.V. Genetic interaction of mammalian IFT-A paralogs regulates cilia disassembly, ciliary entry of membrane protein, Hedgehog signaling, and embryogenesis. FASEB J. 2020, 34, 6369–6381. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Motile Cilia(%) | Primary Cilia(%) | Ciliary Length (μm) | Orientation | ||
|---|---|---|---|---|---|
| Trachea | Epithelium | 100% | - | 5.0–7.0 | Point to airway |
| Cartilage | - | 72.2 ± 1 | 3.0–5.0 | N/A | |
| Bronchioles | Epithelia | 100% | - | 5.0–7.0 | Pointing to airway |
| Smooth muscle | - | 24.7 ± 1.6 | 3.0–5.0 | N/A | |
| Heart | - | 8.7 ± 0.6 | 1.0–3.0 | N/A | |
| Arteries | Endothelium | - | 6 ± 0.1 | 0.5–2.0 | The long axis of epithelia nuclear |
| Immune system | Spleen | - | 13.4 ± 0.9 | 3.0–6.0 | N/A |
| Thymus | - | 20.3 ± 0.9 | 3.0–6.0 | N/A | |
| Liver | Hepatocytes | - | 8.2 ± 0.3 | 0.5–2.0 | N/A |
| Cholangiocytes | - | 43.5 ± 2 | 5.0–7.0 | N/A | |
| Intestine | - | 9.8 ± 0.6 | 3.0–5.0 | N/A | |
| Gallbladder | Epithelia | - | 16.1 ± 0.8 | 1.5–3.0 | N/A |
| Smooth muscle | - | 9.1 ± 0.4 | 3.0–5.0 | The long axis of nuclear | |
| Stomach | Mucosa | - | 5.9 ± 0.2 | 0.5–3.0 | N/A |
| Submucosa | - | 14 ± 0.7 | 3.0–5.0 | N/A | |
| Muscularis externa | - | 9.1 ± 0.3 | 0.5–3.0 | N/A | |
| Serosa | - | 21.2 ± 0.6 | 3.0–5.0 | The long axis of nuclear | |
| Esophagus | Squamous epithelium | - | 7 ± 0.1 | 3.0–5.0 | The long axis of epithelia nuclear |
| Submucosa | - | 7.4 ± 0.1 | 0.5–3.0 | The long axis of nuclear | |
| Muscularis externa | - | 9 ± 0.2 | 0.5–3.0 | N/A | |
| Kidneys | Glomeruli in kidney | - | 9.6 ± 0.4 | 1.5–3.0 | N/A |
| Proximal convoluted tubules | - | 23.9 ± 0.6 | 1.5–3.0 | Toward the lumen of the tubules | |
| Distal convoluted tubules | - | 12.6 ± 0.2 | 1.5–3.0 | N/A | |
| Collecting tubules | - | 54.5 ± 3.8 | 10.0–5.0 | Same direction as fluid | |
| Bladder | Connective tissues | - | 31.8 ± 0.8 | 5.0–7.0 | N/A |
| Testes | Leyding cells | - | 23.0 ± 1.3 | 3.0–10.0 | N/A |
| Spermatogenic cells | - | 10.9 ± 1 | 0.5–2.0 | N/A | |
| Bone | Osteoblasts | - | 86.8 ± 3 | 3.0–3.5 | Long axis of nuclear |
| Osteocytes | - | 81 ± 2.9 | 2.5–3.0 | Long axis of nuclear | |
| Cartilage | Growth plate | - | 68.9 ± 4.1 | 2.0–4 | Long axis of the growth plate chondrocyte |
| Artcular cartilage | - | 49.6 ± 1.5 | 2.0–4.0 | Long axis of the artcular cartilage chondrocyte | |
| Intervertebral discs | Nucleus pulposus | - | 33.2 ± 0.4 | 0.5–15.0 | N/A |
| Annulus fibrosus | - | 36.1 ± 1.9 | 0.5–3.5 | Aligned parallel to the long axis of the aunulus fibrosus cells | |
| Skin | Hair follicles | - | 33.4 ± 1.1 | 1.5–5.0 | N/A |
| Reticular dermis | 52.8 ± 0.9 | 1.5–5.0 | N/A | ||
| Adipose tissue | - | 18.7 ± 0.5 | 3.0–6.0 | N/A | |
| Sutures | - | 85.5 ± 1.1 | 5.0–7.0 | Long axis of the nuclear | |
| Brain | Cerebellum cortex | - | 19.3 ± 0.8 | 0.5–3.0 | N/A |
| Cerebellum | - | 73.9 ± 3.7 | 0.5–3.0 | N/A | |
| Midbrain | - | 9.6 ± 0.1 | 0.5–3.0 | N/A | |
| Cerebral cortex | - | 91.7 ± 3.8 | 0.5–3.0 | Pointing to centre of brain | |
| Endocrine system | Thyroid | - | 23.9 ± 2.2 | 0.5–2.5 | N/A |
| Adrenal gland | - | 7.3 ± 1.2 | 3.0–7.0 | N/A | |
| Islet of Langerhans | - | 21.1 ± 1.6 | 5.5–15.0 | N/A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Yang, S.; Deepak, V.; Chinipardaz, Z.; Yang, S. Identification of Cilia in Different Mouse Tissues. Cells 2021, 10, 1623. https://doi.org/10.3390/cells10071623
Li X, Yang S, Deepak V, Chinipardaz Z, Yang S. Identification of Cilia in Different Mouse Tissues. Cells. 2021; 10(7):1623. https://doi.org/10.3390/cells10071623
Chicago/Turabian StyleLi, Xinhua, Shuting Yang, Vishwa Deepak, Zahra Chinipardaz, and Shuying Yang. 2021. "Identification of Cilia in Different Mouse Tissues" Cells 10, no. 7: 1623. https://doi.org/10.3390/cells10071623