
The Role of BANK1 in B Cell Signaling and Disease


Abstract
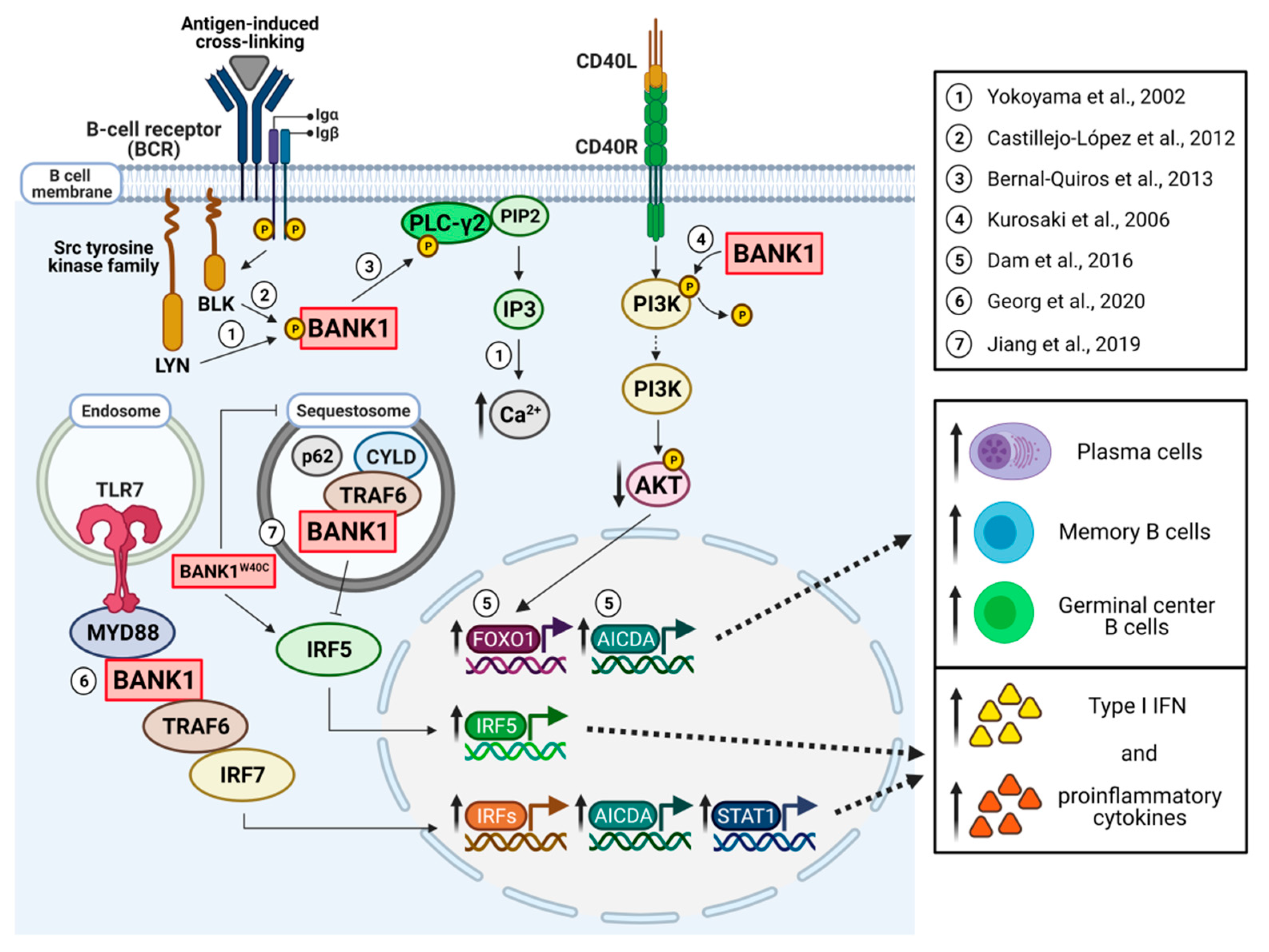
:1. Introduction
2. The Genetic Associations of Variants of BANK1 with Diseases
3. The Role of BANK1 in B Cell Signaling

4. Mechanistic Aspects of the Role of BANK1 in Experimental Disease Model Systems
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yokoyama, K.; Su Ih, I.H.; Tezuka, T.; Yasuda, T.; Mikoshiba, K.; Tarakhovsky, A.; Yamamoto, T. BANK regulates BCR-induced calcium mobilization by promoting tyrosine phosphorylation of IP(3) receptor. Embo J. 2002, 21, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Kozyrev, S.V.; Abelson, A.K.; Wojcik, J.; Zaghlool, A.; Linga Reddy, M.V.; Sanchez, E.; Gunnarsson, I.; Svenungsson, E.; Sturfelt, G.; Jonsen, A.; et al. Functional variants in the B-cell gene BANK1 are associated with systemic lupus erythematosus. Nat. Genet. 2008, 40, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Deshmukh, H.; Lu, R.; Vidal, G.S.; Kelly, J.A.; Kaufman, K.M.; Dominguez, N.; Klein, W.; Kim-Howard, X.; Bruner, G.R.; et al. Replication of the BANK1 genetic association with systemic lupus erythematosus in a European-derived population. Genes Immun. 2009, 10, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Jarvinen, T.M.; Hellquist, A.; Zucchelli, M.; Koskenmies, S.; Panelius, J.; Hasan, T.; Julkunen, H.; D’Amato, M.; Kere, J. Replication of GWAS-identified systemic lupus erythematosus susceptibility genes affirms B-cell receptor pathway signalling and strengthens the role of IRF5 in disease susceptibility in a Northern European population. Rheumatology 2012, 51, 87–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez-Gestal, M.; Calaza, M.; Endreffy, E.; Pullmann, R.; Ordi-Ros, J.; Sebastiani, G.D.; Ruzickova, S.; Jose Santos, M.; Papasteriades, C.; Marchini, M.; et al. Replication of recently identified systemic lupus erythematosus genetic associations: A case-control study. Arthritis Res. Ther. 2009, 11, R69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budarf, M.L.; Goyette, P.; Boucher, G.; Lian, J.; Graham, R.R.; Claudio, J.O.; Hudson, T.; Gladman, D.; Clarke, A.E.; Pope, J.E.; et al. A targeted association study in systemic lupus erythematosus identifies multiple susceptibility alleles. Genes Immun. 2011, 12, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentham, J.; Morris, D.L.; Cunninghame Graham, D.S.; Pinder, C.L.; Tombleson, P.; Behrens, T.W.; Martin, J.; Fairfax, B.P.; Knight, J.C.; Chen, L.; et al. Genetic association analyses implicate aberrant regulation of innate and adaptive immunity genes in the pathogenesis of systemic lupus erythematosus. Nat. Genet. 2015, 47, 1457–1464. [Google Scholar] [CrossRef] [PubMed]
- Langefeld, C.D.; Ainsworth, H.C.; Cunninghame Graham, D.S.; Kelly, J.A.; Comeau, M.E.; Marion, M.C.; Howard, T.D.; Ramos, P.S.; Croker, J.A.; Morris, D.L.; et al. Transancestral mapping and genetic load in systemic lupus erythematosus. Nat. Commun. 2017, 8, 16021. [Google Scholar] [CrossRef]
- Martinez-Bueno, M.; Oparina, N.; Dozmorov, M.G.; Marion, M.C.; Comeau, M.E.; Gilkeson, G.; Kamen, D.; Weisman, M.; Salmon, J.; McCune, J.W.; et al. Trans-Ethnic Mapping of BANK1 Identifies Two Independent SLE-Risk Linkage Groups Enriched for Co-Transcriptional Splicing Marks. Int. J. Mol. Sci. 2018, 19, 2331. [Google Scholar] [CrossRef] [Green Version]
- Chung, S.A.; Tian, C.; Taylor, K.E.; Lee, A.T.; Ortmann, W.A.; Hom, G.; Graham, R.R.; Nititham, J.; Kelly, J.A.; Morrisey, J.; et al. European population substructure is associated with mucocutaneous manifestations and autoantibody production in systemic lupus erythematosus. Arthritis Rheum. 2009, 60, 2448–2456. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.K.; Yang, W.; Zhao, M.; Mok, C.C.; Chan, T.M.; Wong, R.W.; Lee, K.W.; Mok, M.Y.; Wong, S.N.; Ng, I.O.; et al. Association of BANK1 and TNFSF4 with systemic lupus erythematosus in Hong Kong Chinese. Genes Immun. 2009, 10, 414–420. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Shen, N.; Ye, D.Q.; Liu, Q.; Zhang, Y.; Qian, X.X.; Hirankarn, N.; Ying, D.; Pan, H.F.; Mok, C.C.; et al. Genome-wide association study in Asian populations identifies variants in ETS1 and WDFY4 associated with systemic lupus erythematosus. PLoS Genet. 2010, 6, e1000841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, M.; Yu, B.; Wan, J.; Zhang, X.; Wu, Z.; Zhong, Q.; Zhang, W.; Zou, H. Identification of BANK1 polymorphisms by unlabelled probe high resolution melting: Association with systemic lupus erythematosus susceptibility and autoantibody production in Han Chinese. Rheumatology (Oxford) 2011, 50, 473–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez-Bello, J.; Jimenez-Morales, S.; Montufar-Robles, I.; Fragoso, J.M.; Barbosa-Cobos, R.E.; Saavedra, M.A.; Sanchez-Munoz, F. BLK and BANK1 polymorphisms and interactions are associated in Mexican patients with systemic lupus erythematosus. Inflamm. Res. 2019, 68, 705–713. [Google Scholar] [CrossRef]
- Alarcon-Riquelme, M.E.; Ziegler, J.T.; Molineros, J.; Howard, T.D.; Moreno-Estrada, A.; Sanchez-Rodriguez, E.; Ainsworth, H.C.; Ortiz-Tello, P.; Comeau, M.E.; Rasmussen, A.; et al. Genome-Wide Association Study in an Amerindian Ancestry Population Reveals Novel Systemic Lupus Erythematosus Risk Loci and the Role of European Admixture. Arthritis Rheumatol. 2016, 68, 932–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, E.; Comeau, M.E.; Freedman, B.I.; Kelly, J.A.; Kaufman, K.M.; Langefeld, C.D.; Brown, E.E.; Alarcon, G.S.; Kimberly, R.P.; Edberg, J.C.; et al. Identification of novel genetic susceptibility loci in African American lupus patients in a candidate gene association study. Arthritis Rheum. 2011, 63, 3493–3501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, S.F.; Petri, M.; Bradfield, J.P.; Kim, C.E.; Santa, E.; Annaiah, K.; Frackelton, E.C.; Glessner, J.T.; Otieno, F.G.; Shaner, J.L.; et al. Association of the BANK 1 R61H variant with systemic lupus erythematosus in Americans of European and African ancestry. Appl. Clin. Genet. 2009, 2, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orozco, G.; Eyre, S.; Hinks, A.; Bowes, J.; Morgan, A.W.; Wilson, A.G.; Wordsworth, P.; Steer, S.; Hocking, L.; Thomson, W.; et al. Study of the common genetic background for rheumatoid arthritis and systemic lupus erythematosus. Ann> Rheum. Dis. 2011, 70, 463–468. [Google Scholar] [CrossRef]
- Zhou, X.J.; Qi, Y.Y.; Cheng, F.J.; Zhang, H. Genetic interactions between BANK1 and BLK in Chinese patients with systemic lupus erythematosus. J. Rheumatol. 2013, 40, 1772–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.H.; Athanasopoulos, V.; Ellyard, J.I.; Chuah, A.; Cappello, J.; Cook, A.; Prabhu, S.B.; Cardenas, J.; Gu, J.; Stanley, M.; et al. Functional rare and low frequency variants in BLK and BANK1 contribute to human lupus. Nat. Commun. 2019, 10, 2201. [Google Scholar] [CrossRef] [Green Version]
- Dieude, P.; Wipff, J.; Guedj, M.; Ruiz, B.; Melchers, I.; Hachulla, E.; Riemekasten, G.; Diot, E.; Hunzelmann, N.; Sibilia, J.; et al. BANK1 is a genetic risk factor for diffuse cutaneous systemic sclerosis and has additive effects with IRF5 and STAT4. Arthritis Rheum. 2009, 60, 3447–3454. [Google Scholar] [CrossRef]
- Orozco, G.; Abelson, A.K.; Gonzalez-Gay, M.A.; Balsa, A.; Pascual-Salcedo, D.; Garcia, A.; Fernandez-Gutierrez, B.; Petersson, I.; Pons-Estel, B.; Eimon, A.; et al. Study of functional variants of the BANK1 gene in rheumatoid arthritis. Arthritis Rheum. 2009, 60, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Berndt, S.I.; Camp, N.J.; Skibola, C.F.; Vijai, J.; Wang, Z.; Gu, J.; Nieters, A.; Kelly, R.S.; Smedby, K.E.; Monnereau, A.; et al. Meta-analysis of genome-wide association studies discovers multiple loci for chronic lymphocytic leukemia. Nat. Commun. 2016, 7, 10933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, K.W.; Lyu, J.; Lee, S.H.; Choi, B.Y.; Kim, S.S.; Kim, Y. A nonsynonymous SNP in BANK1 is associated with serum LDL cholesterol levels in three Korean populations. J. Hum. Genet. 2015, 60, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Borghi, M.O.; Delgado-Vega, A.M.; Tincani, A.; Meroni, P.L.; Alarcon-Riquelme, M.E. Association of STAT4 and BLK, but not BANK1 or IRF5, with primary antiphospholipid syndrome. Arthritis Rheum. 2009, 60, 2468–2471. [Google Scholar] [CrossRef] [PubMed]
- Speliotes, E.K.; Willer, C.J.; Berndt, S.I.; Monda, K.L.; Thorleifsson, G.; Jackson, A.U.; Lango Allen, H.; Lindgren, C.M.; Luan, J.; Magi, R.; et al. Association analyses of 249,796 individuals reveal 18 new loci associated with body mass index. Nat. Genet. 2010, 42, 937–948. [Google Scholar] [CrossRef] [Green Version]
- Mak, A.C.; Tang, P.L.; Cleveland, C.; Smith, M.H.; Kari Connolly, M.; Katsumoto, T.R.; Wolters, P.J.; Kwok, P.Y.; Criswell, L.A. Brief Report: Whole-Exome Sequencing for Identification of Potential Causal Variants for Diffuse Cutaneous Systemic Sclerosis. Arthritis Rheumatol. 2016, 68, 2257–2262. [Google Scholar] [CrossRef] [Green Version]
- Baranski, T.J.; Kraja, A.T.; Fink, J.L.; Feitosa, M.; Lenzini, P.A.; Borecki, I.B.; Liu, C.T.; Cupples, L.A.; North, K.E.; Province, M.A. A high throughput, functional screen of human Body Mass Index GWAS loci using tissue-specific RNAi Drosophila melanogaster crosses. PLoS Genet. 2018, 14, e1007222. [Google Scholar] [CrossRef] [Green Version]
- Reksten, T.R.; Johnsen, S.J.; Jonsson, M.V.; Omdal, R.; Brun, J.G.; Theander, E.; Eriksson, P.; Wahren-Herlenius, M.; Jonsson, R.; Nordmark, G. Genetic associations to germinal centre formation in primary Sjogren’s syndrome. Ann. Rheum. Dis. 2014, 73, 1253–1258. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Xu, J.; Wu, Z.; Li, P.; Chen, H.; Su, J.; You, X.; Li, M.; Zhao, Y.; Tian, X.; et al. Polymorphisms in the FAM167A-BLK, but not BANK1, are associated with primary Sjogren’s syndrome in a Han Chinese population. Clin. Exp. Rheumatol. 2013, 31, 704–710. [Google Scholar]
- Lessard, C.J.; Li, H.; Adrianto, I.; Ice, J.A.; Rasmussen, A.; Grundahl, K.M.; Kelly, J.A.; Dozmorov, M.G.; Miceli-Richard, C.; Bowman, S.; et al. Variants at multiple loci implicated in both innate and adaptive immune responses are associated with Sjogren’s syndrome. Nat. Genet. 2013, 45, 1284–1292. [Google Scholar] [CrossRef]
- Coustet, B.; Dieude, P.; Guedj, M.; Bouaziz, M.; Avouac, J.; Ruiz, B.; Hachulla, E.; Diot, E.; Kracowski, J.L.; Tiev, K.; et al. C8orf 13/BLK is a genetic risk locus for systemic sclerosis and has additive effect with BANK1: Results from a large French cohort and meta-analysis. Arthritis Rheum. 2011, 63, 2091–2096. [Google Scholar] [CrossRef]
- Castillejo-Lopez, C.; Delgado-Vega, A.M.; Wojcik, J.; Kozyrev, S.V.; Thavathiru, E.; Wu, Y.Y.; Sanchez, E.; Pollmann, D.; Lopez-Egido, J.R.; Fineschi, S.; et al. Genetic and physical interaction of the B-cell systemic lupus erythematosus-associated genes BANK1 and BLK. Ann. Rheum. Dis. 2012, 71, 136–142. [Google Scholar] [CrossRef] [Green Version]
- Genin, E.; Coustet, B.; Allanore, Y.; Ito, I.; Teruel, M.; Constantin, A.; Schaeverbeke, T.; Ruyssen-Witrand, A.; Tohma, S.; Cantagrel, A.; et al. Epistatic interaction between BANK1 and BLK in rheumatoid arthritis: Results from a large trans-ethnic meta-analysis. PLoS ONE 2013, 8, e61044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, J.; Li, J.; Xin, Q.; Shan, S.; Bian, X.; Yuan, Q.; Liu, N.; Ma, X.; Li, Y.; Liu, Q. Gene-gene interaction of ATG5, ATG7, BLK and BANK1 in systemic lupus erythematosus. Int. J. Rheum. Dis. 2016, 19, 1284–1293. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Barreiro, A.; Bernal-Quiros, M.; Georg, I.; Maranon, C.; Alarcon-Riquelme, M.E.; Castillejo-Lopez, C. The SLE variant Ala71Thr of BLK severely decreases protein abundance and binding to BANK1 through impairment of the SH3 domain function. Genes Immun. 2016, 17, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Georg, I.; Diaz-Barreiro, A.; Morell, M.; Pey, A.L.; Alarcon-Riquelme, M.E. BANK1 interacts with TRAF6 and MyD88 in innate immune signaling in B cells. Cell Mol. Immunol. 2020, 17, 954–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dam, E.M.; Habib, T.; Chen, J.; Funk, A.; Glukhova, V.; Davis-Pickett, M.; Wei, S.; James, R.; Buckner, J.H.; Cerosaletti, K. The BANK1 SLE-risk variants are associated with alterations in peripheral B cell signaling and development in humans. Clin. Immunol. 2016, 173, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Wilbe, M.; Jokinen, P.; Truve, K.; Seppala, E.H.; Karlsson, E.K.; Biagi, T.; Hughes, A.; Bannasch, D.; Andersson, G.; Hansson-Hamlin, H.; et al. Genome-wide association mapping identifies multiple loci for a canine SLE-related disease complex. Nat. Genet. 2010, 42, 250–254. [Google Scholar] [CrossRef]
- Battersby, A.; Csiszar, A.; Leptin, M.; Wilson, R. Isolation of proteins that interact with the signal transduction molecule Dof and identification of a functional domain conserved between Dof and vertebrate BCAP. J. Mol. Biol. 2003, 329, 479–493. [Google Scholar] [CrossRef]
- Troutman, T.D.; Hu, W.; Fulenchek, S.; Yamazaki, T.; Kurosaki, T.; Bazan, J.F.; Pasare, C. Role for B-cell adapter for PI3K (BCAP) as a signaling adapter linking Toll-like receptors (TLRs) to serine/threonine kinases PI3K/Akt. Proc. Natl. Acad. Sci. USA 2012, 109, 273–278. [Google Scholar] [CrossRef] [Green Version]
- Kozyrev, S.V.; Bernal-Quiros, M.; Alarcon-Riquelme, M.E.; Castillejo-Lopez, C. The dual effect of the lupus-associated polymorphism rs10516487 on BANK1 gene expression and protein localization. Genes Immun. 2012, 13, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Feng, J.; Wen, R.; Marine, J.C.; Sangster, M.Y.; Parganas, E.; Hoffmeyer, A.; Jackson, C.W.; Cleveland, J.L.; Murray, P.J.; et al. Phospholipase Cgamma2 is essential in the functions of B cell and several Fc receptors. Immunity 2000, 13, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Bernal-Quiros, M.; Wu, Y.Y.; Alarcon-Riquelme, M.E.; Castillejo-Lopez, C. BANK1 and BLK Act through Phospholipase C Gamma 2 in B-Cell Signaling. PLoS ONE 2013, 8, e59842. [Google Scholar] [CrossRef] [Green Version]
- Aiba, Y.; Yamazaki, T.; Okada, T.; Gotoh, K.; Sanjo, H.; Ogata, M.; Kurosaki, T. BANK negatively regulates Akt activation and subsequent B cell responses. Immunity 2006, 24, 259–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, T.; Sato, S.; Ishii, K.J.; Coban, C.; Hemmi, H.; Yamamoto, M.; Terai, K.; Matsuda, M.; Inoue, J.; Uematsu, S.; et al. Interferon-alpha induction through Toll-like receptors involves a direct interaction of IRF7 with MyD88 and TRAF6. Nat. Immunol. 2004, 5, 1061–1068. [Google Scholar] [CrossRef]
- Kim, J.Y.; Ozato, K. The sequestosome 1/p62 attenuates cytokine gene expression in activated macrophages by inhibiting IFN regulatory factor 8 and TNF receptor-associated factor 6/NF-kappaB activity. J. Immunol. 2009, 182, 2131–2140. [Google Scholar] [CrossRef]
- Han, B.; Wu, J. DcR3 protects islet beta cells from apoptosis through modulating Adcyap1 and Bank1 expression. J. Immunol. 2009, 183, 8157–8166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fei, H.; Zhao, B.; Zhao, S.; Wang, Q. Requirements of calcium fluxes and ERK kinase activation for glucose- and interleukin-1beta-induced beta-cell apoptosis. Mol. Cell Biochem. 2008, 315, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Christensen, S.R.; Kashgarian, M.; Alexopoulou, L.; Flavell, R.A.; Akira, S.; Shlomchik, M.J. Toll-like receptor 9 controls anti-DNA autoantibody production in murine lupus. J. Exp. Med. 2005, 202, 321–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.Y.; Kumar, R.; Haque, M.S.; Castillejo-Lopez, C.; Alarcon-Riquelme, M.E. BANK1 Controls CpG-Induced IL-6 Secretion via a p38 and MNK1/2/eIF4E Translation Initiation Pathway. J. Immunol. 2013, 191, 6110–6116. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.Y.; Kumar, R.; Iida, R.; Bagavant, H.; Alarcon-Riquelme, M.E. BANK1 Regulates IgG Production in a Lupus Model by Controlling TLR7-Dependent STAT1 Activation. PLoS ONE 2016, 11, e0156302. [Google Scholar] [CrossRef] [PubMed]
- Morel, L.; Blenman, K.R.; Croker, B.P.; Wakeland, E.K. The major murine systemic lupus erythematosus susceptibility locus, Sle1, is a cluster of functionally related genes. Proc. Natl Acad. Sci. USA 2001, 98, 1787–1792. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, S.; Tus, K.; Li, Q.Z.; Wang, A.; Tian, X.H.; Zhou, J.; Liang, C.; Bartov, G.; McDaniel, L.D.; Zhou, X.J.; et al. A Tlr7 translocation accelerates systemic autoimmunity in murine lupus. Proc. Natl. Acad. Sci. USA 2006, 103, 9970–9975. [Google Scholar] [CrossRef] [Green Version]
- Santiago-Raber, M.L.; Dunand-Sauthier, I.; Wu, T.; Li, Q.Z.; Uematsu, S.; Akira, S.; Reith, W.; Mohan, C.; Kotzin, B.L.; Izui, S. Critical role of TLR7 in the acceleration of systemic lupus erythematosus in TLR9-deficient mice. J. Autoimmun. 2010, 34, 339–348. [Google Scholar] [CrossRef]
- Fairhurst, A.M.; Hwang, S.H.; Wang, A.; Tian, X.H.; Boudreaux, C.; Zhou, X.J.; Casco, J.; Li, Q.Z.; Connolly, J.E.; Wakeland, E.K. Yaa autoimmune phenotypes are conferred by overexpression of TLR7. Eur. J. Immunol. 2008, 38, 1971–1978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiratsuka, T.; Takei, Y.; Ohmori, R.; Imai, Y.; Ozeki, M.; Tamaki, K.; Haga, H.; Nakamura, T.; Tsuruyama, T. ZFP521 contributes to pre-B-cell lymphomagenesis through modulation of the pre-B-cell receptor signaling pathway. Oncogene 2016, 35, 3227–3238. [Google Scholar] [CrossRef] [PubMed]
- Fugmann, T.; Sofron, A.; Ritz, D.; Bootz, F.; Neri, D. The MHC Class II Immunopeptidome of Lymph Nodes in Health and in Chemically Induced Colitis. J. Immunol. 2017, 198, 1357–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Ren, J.; Yang, Y.; Sun, J.; Zhou, X.; Zheng, S.; Xuan, D.; Xue, Y.; Fan, H.; Zhang, J.; et al. BANK1 alters B cell responses and influences the interactions between B cells and induced T regulatory cells in mice with collagen-induced arthritis. Arthritis Res. Ther. 2018, 20, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
| Disease | SNPs 1 | Location in Protein | Study |
|---|---|---|---|
| SLE | rs10516487 | TIR domain | [2] |
| SLE | rs17266594 | -- | [2] |
| SLE | rs3733197 | Ankyrin repeats | [2] |
| SLE | rs17266594 | --- | [3] |
| SLE | rs10028805 | --- | [4,5,6,7,8,9,12,13,14,16,17,18,19] |
| SLE | W40C | TIR domain | [20] |
| Diffuse SSC | rs10516487 | TIR domain | [21] |
| Diffuse SSC | rs3733197 | Ankyrin repeats | [21] |
| RA | rs10516487 | TIR domain | [18,22] |
| Chronic Lymphocytic Leukemia | rs10028805 | --- | [23] |
| LDL Cholesterol | rs3733197 | Ankyrin repeats | [24] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez Hernández, G.; Morell, M.; Alarcón-Riquelme, M.E. The Role of BANK1 in B Cell Signaling and Disease. Cells 2021, 10, 1184. https://doi.org/10.3390/cells10051184
Gómez Hernández G, Morell M, Alarcón-Riquelme ME. The Role of BANK1 in B Cell Signaling and Disease. Cells. 2021; 10(5):1184. https://doi.org/10.3390/cells10051184
Chicago/Turabian StyleGómez Hernández, Gonzalo, María Morell, and Marta E. Alarcón-Riquelme. 2021. "The Role of BANK1 in B Cell Signaling and Disease" Cells 10, no. 5: 1184. https://doi.org/10.3390/cells10051184