
Direct Stimulatory Effects of the CB2 Ligand JTE 907 in Human and Mouse Islets



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Isolation of Human and Mouse Islets
2.4. Dynamic Insulin Secretion
2.5. Caspase 3/7 Activities
2.6. Islet β-Cell Proliferation
2.7. IP1 and Cyclic AMP Accumulation
2.8. Single Cell Calcium Microfluorimetry
2.9. Statistical Analyses
3. Results
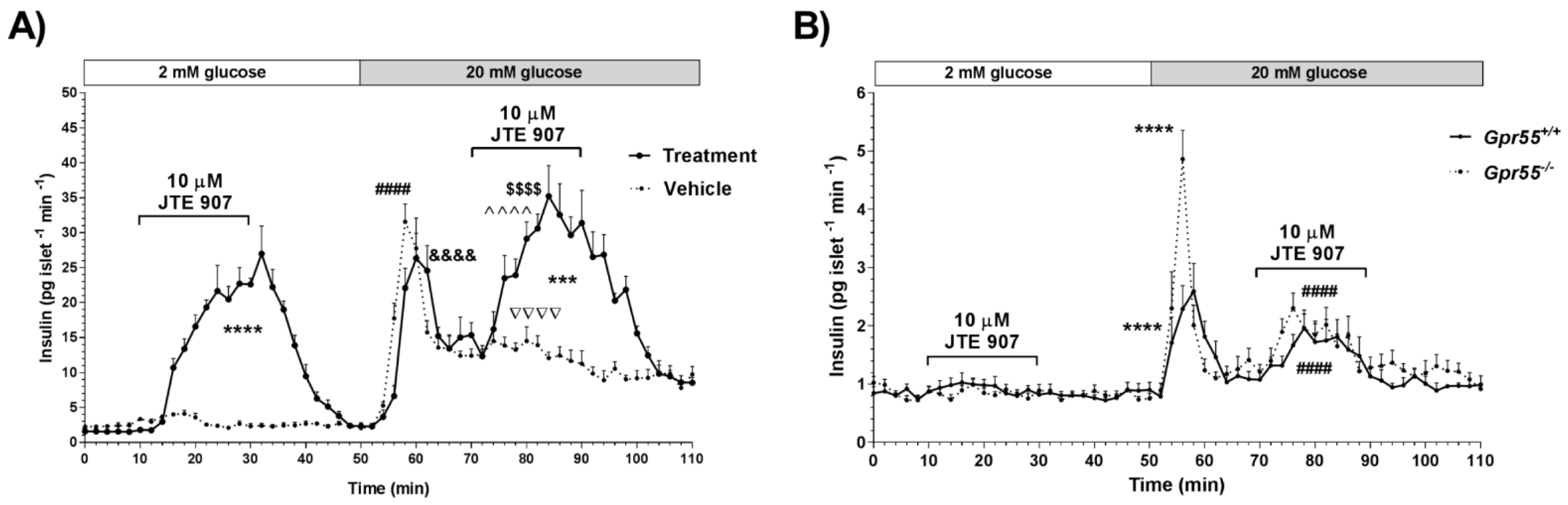
3.1. JTE 907 Stimulates Insulin Secretion in a GPR55-Independent Manner
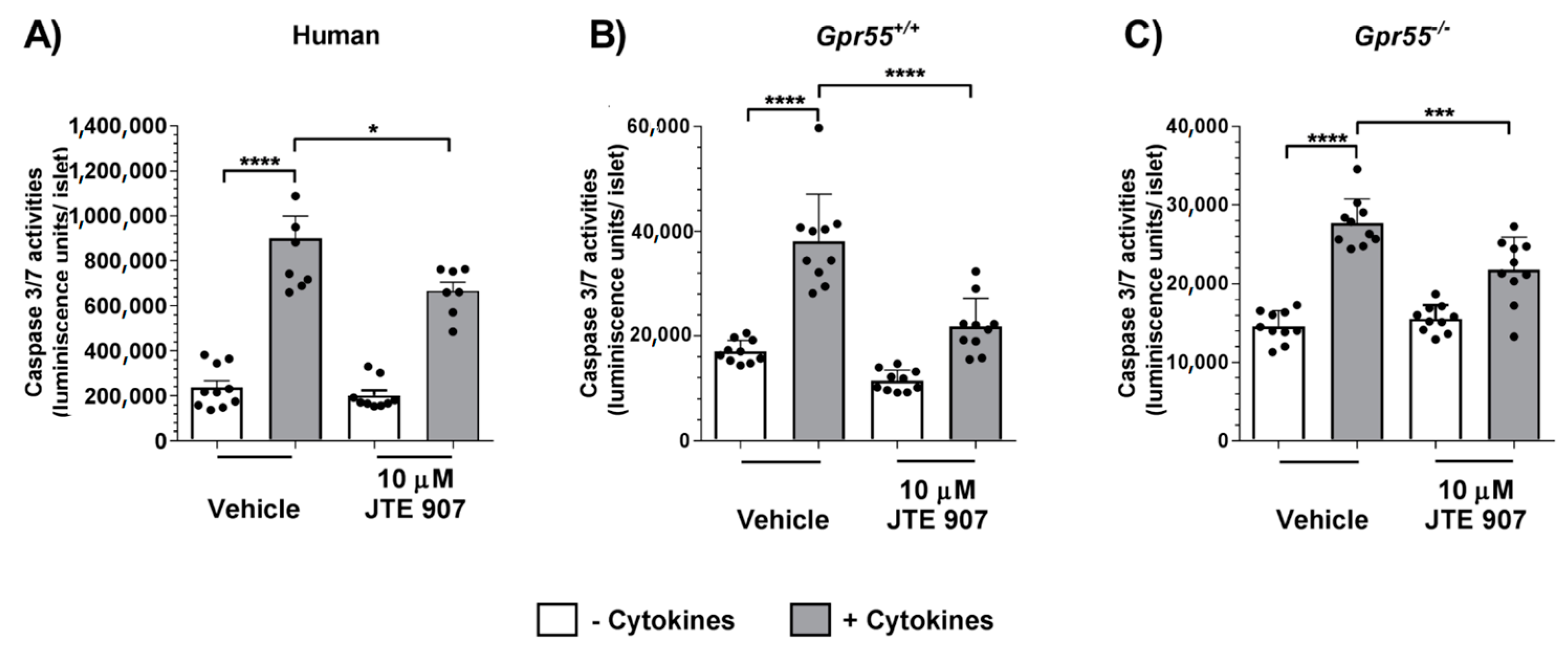
3.2. JTE 907 Protects Human and Mouse Islets from Cytokine-Induced Apoptosis
3.3. JTE 907 Induces Human β-Cell Proliferation
3.4. JTE 907 Is Not Coupled to Gs or Gi Signaling in Human and Mouse Islets
3.5. JTE 907 Increases Islet IP1 Levels in Human and Mouse Islets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BMI | body mass index |
| BSA | bovine serum albumin |
| cAMP | 3′,5′-cyclic adenosine monophosphate |
| Cch | carbachol |
| CB1 | canonical cannabinoid receptor type 1 |
| CB2 | canonical cannabinoid receptor type 2 |
| CMRL | Connaught Medical Research Laboratories |
| DAPI | 4′,6-diamidino-2-phenylindole |
| DMSO | dimethyl sulfoxide |
| ECS | endocannabinoid system |
| EDTA | ethylenediaminetetraacetic acid |
| FBS | fetal bovine serum |
| GLP-1 | glucagon-like peptide-1 |
| GPCR | G protein-coupled receptor |
| GPR55 | G protein-coupled receptor 55 |
| Gq | Gq alpha subunit |
| Gs | Gs alpha subunit |
| HBSS | Hank’s Balanced Salt Solution |
| HEPES | 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid |
| HTRF | Homogeneous Time Resolved Fluorescence |
| IBMX | 3-isobutyl-1-methylxanthine |
| IFNγ | interferon gamma |
| IL-1β | interleukin-1 beta |
| IP1 | inositol-1-phosphate |
| IP3 | inositol trisphosphate |
| JTE 907 | N-(1,3 benzodioxol-5-ylmethyl)-1,2-dihydro-7-methoxy-2-oxo-8-(pentyloxy)-3-quinolinecar boxamide |
| JWH 015 | (2-Methyl-1-propyl-1H-indol-3-yl)-1-naphthalenylmethanone |
| LH-21 | 5-(4-chlorophenyl)-1-(2,4-dichlorophenyl)-3-hexyl-1H-1,2,4-triazole |
| M3 | muscarinic receptors |
| RIA | radioimmunoassay |
| RPMI | Roswell Park Memorial Institute |
| Rimonabant (SR141716A) | 5-(4-chlorophenyl)-1-(2,4-dichloro-phenyl)-4-methyl-N-(piperidin1-yl)-1H-pyrazole-3-carboxamide |
| TNFα | tumor necrosis factor alpha |
| T2D | type 2 diabetes |
References
- Frank, J.A.; Broichhagen, J.; Yushchenko, D.A.; Trauner, D.; Schultz, C.; Hodson, D.J. Optical tools for understanding the complexity of beta-cell signalling and insulin release. Nat. Rev. Endocrinol. 2018, 14, 721–737. [Google Scholar] [CrossRef]
- Persaud, S.J. Islet G-protein coupled receptors: Therapeutic potential for diabetes. Curr. Opin. Pharmacol. 2017, 37, 24–28. [Google Scholar] [CrossRef] [Green Version]
- Despres, J.P.; Golay, A.; Sjostrom, L. Effects of rimonabant on metabolic risk factors in overweight patients with dyslipidemia. N. Engl. J. Med. 2005, 353, 2121–2134. [Google Scholar] [CrossRef] [Green Version]
- Van Gaal, L.F.; Rissanen, A.M.; Scheen, A.J.; Ziegler, O.; Rossner, S.; RIO-Europe Study Group. Effects of the cannabinoid-1 receptor blocker rimonabant on weight reduction and cardiovascular risk factors in overweight patients: 1-year experience from the RIO-Europe study. Lancet 2005, 365, 1389–1397. [Google Scholar] [CrossRef]
- Pi-Sunyer, F.X.; Aronne, L.J.; Heshmati, H.M.; Devin, J.; Rosenstock, J.; RIO-North America Study Group. Effect of rimonabant, a cannabinoid-1 receptor blocker, on weight and cardiometabolic risk factors in overweight or obese patients: RIO-North America: A randomized controlled trial. JAMA 2006, 295, 761–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Jones, P.M.; Persaud, S.J. Role of the endocannabinoid system in food intake, energy homeostasis and regulation of the endocrine pancreas. Pharmacol. Ther. 2011, 129, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Vilches-Flores, A.; Zhao, M.; Amiel, S.A.; Jones, P.M.; Persaud, S.J. Expression and function of monoacylglycerol lipase in mouse beta-cells and human islets of Langerhans. Cell Physiol. Biochem. 2012, 30, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Bowe, J.E.; Jones, P.M.; Persaud, S.J. Expression and function of cannabinoid receptors in mouse islets. Islets 2010, 2, 293–302. [Google Scholar] [CrossRef]
- Ruz-Maldonado, I.; Liu, B.; Atanes, P.; Pingitore, A.; Huang, G.C.; Choudhary, P.; Persaud, S.J. The cannabinoid ligands SR141716A and AM251 enhance human and mouse islet function via GPR55-independent signalling. Cell Mol. Life Sci. 2020, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Bowe, J.E.; Huang, G.C.; Amiel, S.A.; Jones, P.M.; Persaud, S.J. Cannabinoid receptor agonists and antagonists stimulate insulin secretion from isolated human islets of Langerhans. Diabetes Obes. Metab. 2011, 13, 903–910. [Google Scholar] [CrossRef]
- Nakata, M.; Yada, T. Cannabinoids inhibit insulin secretion and cytosolic Ca2+ oscillation in islet beta-cells via CB1 receptors. Regul. Pept. 2008, 145, 49–53. [Google Scholar] [CrossRef]
- Juan-Pico, P.; Fuentes, E.; Bermudez-Silva, F.J.; Javier Diaz-Molina, F.; Ripoll, C.; Rodriguez de Fonseca, F.; Nadal, A. Cannabinoid receptors regulate Ca2+ signals and insulin secretion in pancreatic beta-cell. Cell Calcium 2006, 39, 155–162. [Google Scholar] [CrossRef]
- Bermudez-Silva, F.J.; Suarez, J.; Baixeras, E.; Cobo, N.; Bautista, D.; Cuesta-Munoz, A.L.; Fuentes, E.; Juan-Pico, P.; Castro, M.J.; Milman, G.; et al. Presence of functional cannabinoid receptors in human endocrine pancreas. Diabetologia 2008, 51, 476–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matias, I.; Gonthier, M.P.; Orlando, P.; Martiadis, V.; De Petrocellis, L.; Cervino, C.; Petrosino, S.; Hoareau, L.; Festy, F.; Pasquali, R.; et al. Regulation, function, and dysregulation of endocannabinoids in models of adipose and beta-pancreatic cells and in obesity and hyperglycemia. J. Clin. Endocrinol. Metab. 2006, 91, 3171–3180. [Google Scholar] [CrossRef]
- Ruz-Maldonado, I.; Pingitore, A.; Liu, B.; Atanes, P.; Huang, G.C.; Baker, D.; Alonso, F.J.; Bermudez-Silva, F.J.; Persaud, S.J. LH-21 and abnormal cannabidiol improve beta-cell function in isolated human and mouse islets through GPR55-dependent and -independent signalling. Diabetes Obes. Metab. 2018, 20, 930–942. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Doyle, M.E.; Liu, Z.; Lao, Q.; Shin, Y.K.; Carlson, O.D.; Kim, H.S.; Thomas, S.; Napora, J.K.; Lee, E.K.; et al. Cannabinoids inhibit insulin receptor signaling in pancreatic beta-cells. Diabetes 2011, 60, 1198–1209. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Mariscal, I.; Montoro, R.A.; Doyle, M.E.; Liu, Q.R.; Rouse, M.; O’Connell, J.F.; Santa-Cruz Calvo, S.; Krzysik-Walker, S.M.; Ghosh, S.; Carlson, O.D.; et al. Absence of cannabinoid 1 receptor in beta cells protects against high-fat/high-sugar diet-induced beta cell dysfunction and inflammation in murine islets. Diabetologia 2018, 61, 1470–1483. [Google Scholar] [CrossRef] [Green Version]
- Soethoudt, M.; Grether, U.; Fingerle, J.; Grim, T.W.; Fezza, F.; de Petrocellis, L.; Ullmer, C.; Rothenhausler, B.; Perret, C.; van Gils, N.; et al. Cannabinoid CB2 receptor ligand profiling reveals biased signalling and off-target activity. Nat. Commun. 2017, 8, 13958. [Google Scholar] [CrossRef] [PubMed]
- Morales, P.; Reggio, P.H. An Update on Non-CB1, Non-CB2 Cannabinoid Related G-Protein-Coupled Receptors. Cannabis Cannabinoid Res. 2017, 2, 265–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Zerbo, S.Y.; Ruz-Maldonado, I.; Espinosa-Jimenez, V.; Rafacho, A.; Gomez-Conde, A.I.; Sanchez-Salido, L.; Cobo-Vuilleumier, N.; Gauthier, B.R.; Tinahones, F.J.; Persaud, S.J.; et al. The cannabinoid ligand LH-21 reduces anxiety and improves glucose handling in diet-induced obese pre-diabetic mice. Sci. Rep. 2017, 7, 3946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Song, S.; Jones, P.M.; Persaud, S.J. GPR55: From orphan to metabolic regulator? Pharmacol. Ther. 2015, 145, 35–42. [Google Scholar] [CrossRef]
- Liu, B.; Song, S.; Ruz-Maldonado, I.; Pingitore, A.; Huang, G.C.; Baker, D.; Jones, P.M.; Persaud, S.J. GPR55-dependent stimulation of insulin secretion from isolated mouse and human islets of Langerhans. Diabetes Obes. Metab. 2016, 18, 1263–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, B.; Ma, S.; Wang, D.H. TRPV1 mediates glucose-induced insulin secretion through releasing neuropeptides. In Vivo 2019, 33, 1431–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sisay, S.; Pryce, G.; Jackson, S.J.; Tanner, C.; Ross, R.A.; Michael, G.J.; Selwood, D.L.; Giovannoni, G.; Baker, D. Genetic background can result in a marked or minimal effect of gene knockout (GPR55 and CB2 receptor) in experimental autoimmune encephalomyelitis models of multiple sclerosis. PLoS ONE 2013, 8, e76907. [Google Scholar] [CrossRef]
- Huang, G.C.; Zhao, M.; Jones, P.; Persaud, S.; Ramracheya, R.; Lobner, K.; Christie, M.R.; Banga, J.P.; Peakman, M.; Sirinivsan, P.; et al. The development of new density gradient media for purifying human islets and islet-quality assessments. Transplantation 2004, 77, 143–145. [Google Scholar] [CrossRef] [PubMed]
- Atanes, P.; Ruz-Maldonado, I.; Olaniru, O.E.; Persaud, S.J. Assessing mouse islet function. Methods Mol. Biol. 2020, 2128, 241–268. [Google Scholar] [CrossRef]
- Pingitore, A.; Chambers, E.S.; Hill, T.; Maldonado, I.R.; Liu, B.; Bewick, G.; Morrison, D.J.; Preston, T.; Wallis, G.A.; Tedford, C.; et al. The diet-derived short chain fatty acid propionate improves beta-cell function in humans and stimulates insulin secretion from human islets in vitro. Diabetes Obes. Metab. 2017, 19, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Gey, G.O.; Gey, M.K. The maintenance of human normal cells and tumor cells in continuous culture: I. Preliminary report: Cultivation of mesoblastic tumors and normal tissue and notes on methods of cultivation. Am. J. Cancer 1936, 27, 45–76. [Google Scholar] [CrossRef]
- Gilon, P.; Henquin, J.C. Mechanisms and physiological significance of the cholinergic control of pancreatic beta-cell function. Endocr Rev. 2001, 22, 565–604. [Google Scholar] [CrossRef] [Green Version]
- Henstridge, C.M.; Balenga, N.A.; Kargl, J.; Andradas, C.; Brown, A.J.; Irving, A.; Sanchez, C.; Waldhoer, M. Minireview: Recent developments in the physiology and pathology of the lysophosphatidylinositol-sensitive receptor GPR55. Mol. Endocrinol. 2011, 25, 1835–1848. [Google Scholar] [CrossRef] [Green Version]
- Cabrera, O.; Berman, D.M.; Kenyon, N.S.; Ricordi, C.; Berggren, P.O.; Caicedo, A. The unique cytoarchitecture of human pancreatic islets has implications for islet cell function. Proc. Natl. Acad. Sci. USA 2006, 103, 2334–2339. [Google Scholar] [CrossRef] [Green Version]
- Dolensek, J.; Rupnik, M.S.; Stozer, A. Structural similarities and differences between the human and the mouse pancreas. Islets 2015, 7, e1024405. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Xavier, G.; Rutter, G.A. Metabolic and functional heterogeneity in pancreatic beta cells. J. Mol. Biol. 2020, 432, 1395–1406. [Google Scholar] [CrossRef]
- Pingitore, A.; Ruz-Maldonado, I.; Liu, B.; Huang, G.C.; Choudhary, P.; Persaud, S.J. Dynamic profiling of insulin secretion and ATP generation in isolated human and mouse islets reveals differential glucose sensitivity. Cell Physiol. Biochem. 2017, 44, 1352–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linnemann, A.K.; Baan, M.; Davis, D.B. Pancreatic beta-cell proliferation in obesity. Adv. Nutr. 2014, 5, 278–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwamura, H.; Suzuki, H.; Ueda, Y.; Kaya, T.; Inaba, T. In Vitro and in vivo pharmacological characterization of JTE-907, a novel selective ligand for cannabinoid CB2 receptor. J. Pharmacol. Exp. Ther. 2001, 296, 420–425. [Google Scholar]
- Sassmann, A.; Gier, B.; Grone, H.J.; Drews, G.; Offermanns, S.; Wettschureck, N. The Gq/G11-mediated signaling pathway is critical for autocrine potentiation of insulin secretion in mice. J. Clin. Investig. 2010, 120, 2184–2193. [Google Scholar] [CrossRef] [Green Version]
- Console-Bram, L.; Brailoiu, E.; Brailoiu, G.C.; Sharir, H.; Abood, M.E. Activation of GPR18 by cannabinoid compounds: A tale of biased agonism. Br. J. Pharmacol. 2014, 171, 3908–3917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertwee, R.G.; Howlett, A.C.; Abood, M.E.; Alexander, S.P.; Di Marzo, V.; Elphick, M.R.; Greasley, P.J.; Hansen, H.S.; Kunos, G.; Mackie, K.; et al. International union of basic and clinical pharmacology. LXXIX. Cannabinoid receptors and their ligands: Beyond CB(1) and CB(2). Pharmacol. Rev. 2010, 62, 588–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amisten, S.; Atanes, P.; Hawkes, R.; Ruz-Maldonado, I.; Liu, B.; Parandeh, F.; Zhao, M.; Huang, G.C.; Salehi, A.; Persaud, S.J. A comparative analysis of human and mouse islet G-protein coupled receptor expression. Sci. Rep. 2017, 7, 46600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.W.; Rivera, R.; Gardell, S.; Dubin, A.E.; Chun, J. GPR92 as a new G12/13- and Gq-coupled lysophosphatidic acid receptor that increases cAMP, LPA5. J. Biol. Chem. 2006, 281, 23589–23597. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruz-Maldonado, I.; Atanes, P.; Huang, G.C.; Liu, B.; Persaud, S.J. Direct Stimulatory Effects of the CB2 Ligand JTE 907 in Human and Mouse Islets. Cells 2021, 10, 700. https://doi.org/10.3390/cells10030700
Ruz-Maldonado I, Atanes P, Huang GC, Liu B, Persaud SJ. Direct Stimulatory Effects of the CB2 Ligand JTE 907 in Human and Mouse Islets. Cells. 2021; 10(3):700. https://doi.org/10.3390/cells10030700
Chicago/Turabian StyleRuz-Maldonado, Inmaculada, Patricio Atanes, Guo Cai Huang, Bo Liu, and Shanta J Persaud. 2021. "Direct Stimulatory Effects of the CB2 Ligand JTE 907 in Human and Mouse Islets" Cells 10, no. 3: 700. https://doi.org/10.3390/cells10030700