
Modulatory Impact of Adipose-Derived Mesenchymal Stem Cells of Ankylosing Spondylitis Patients on T Helper Cell Differentiation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Ethics Approval
2.2. ASCs Isolation and Culture
2.3. Cell Co-Cultures
2.4. Flow Cytometry Analysis
2.5. Enzyme-Linked Immunosorbent Assays (ELISA)
2.6. Polymerase Chain Reaction (PCR)
2.7. Statistical Analysis
3. Results
3.1. Patients
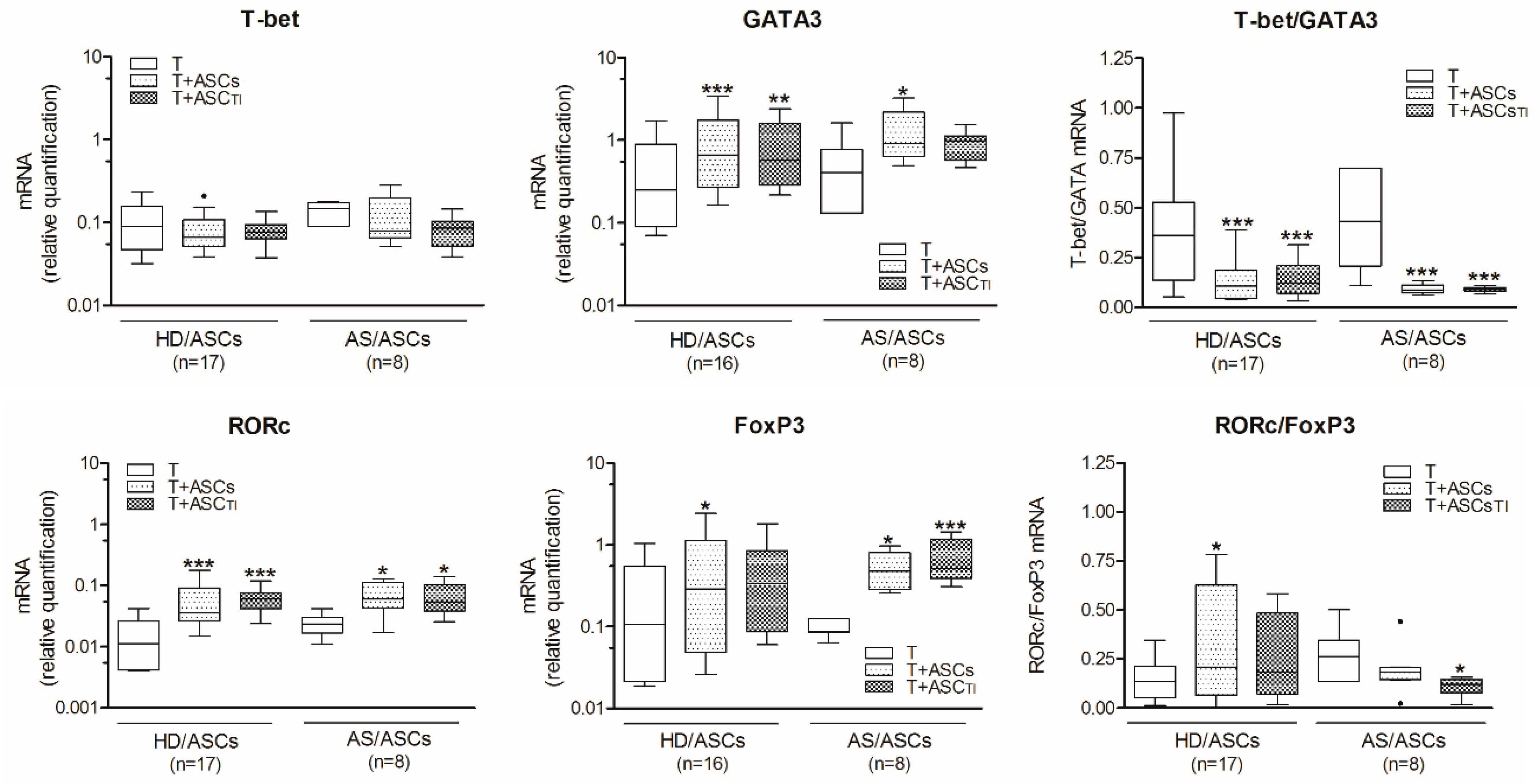
3.2. ASCs Decrease T-Bet/GATA3 and RORc/FoxP3 Ratio in Activated CD4+ T Cells
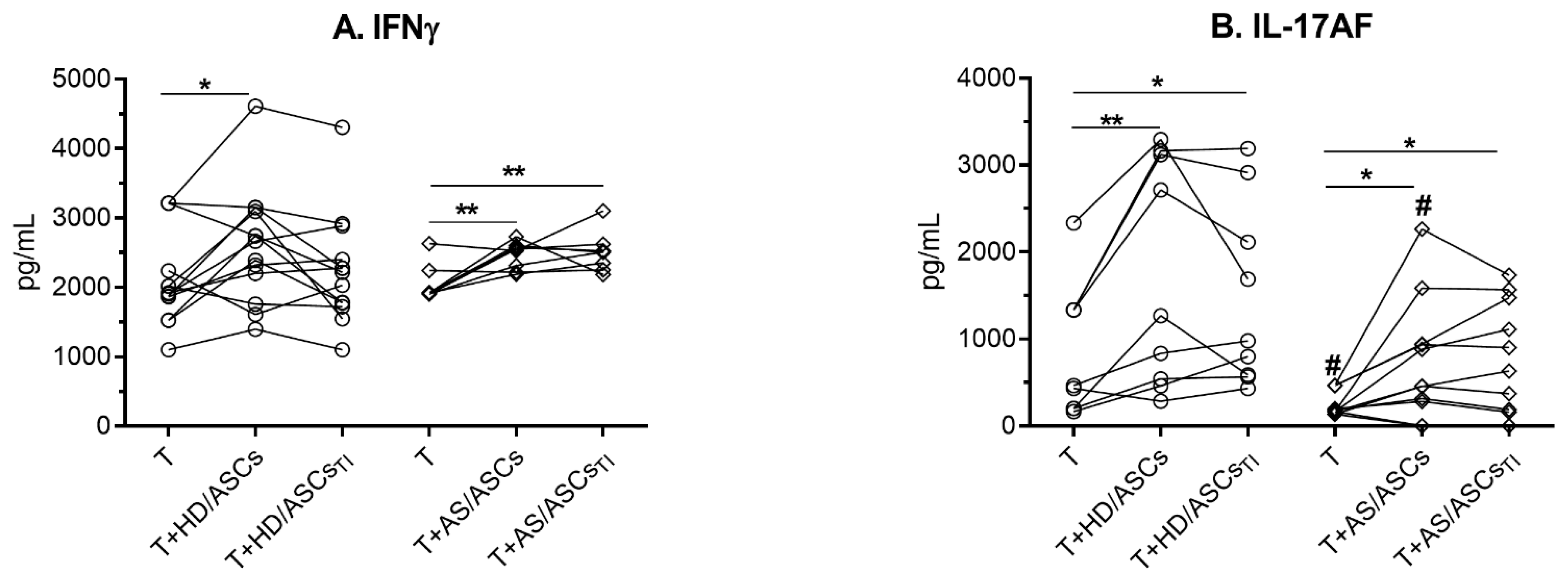
3.3. The Effects of ASCs on the Release of Th1 and Th17 Specific Cytokines
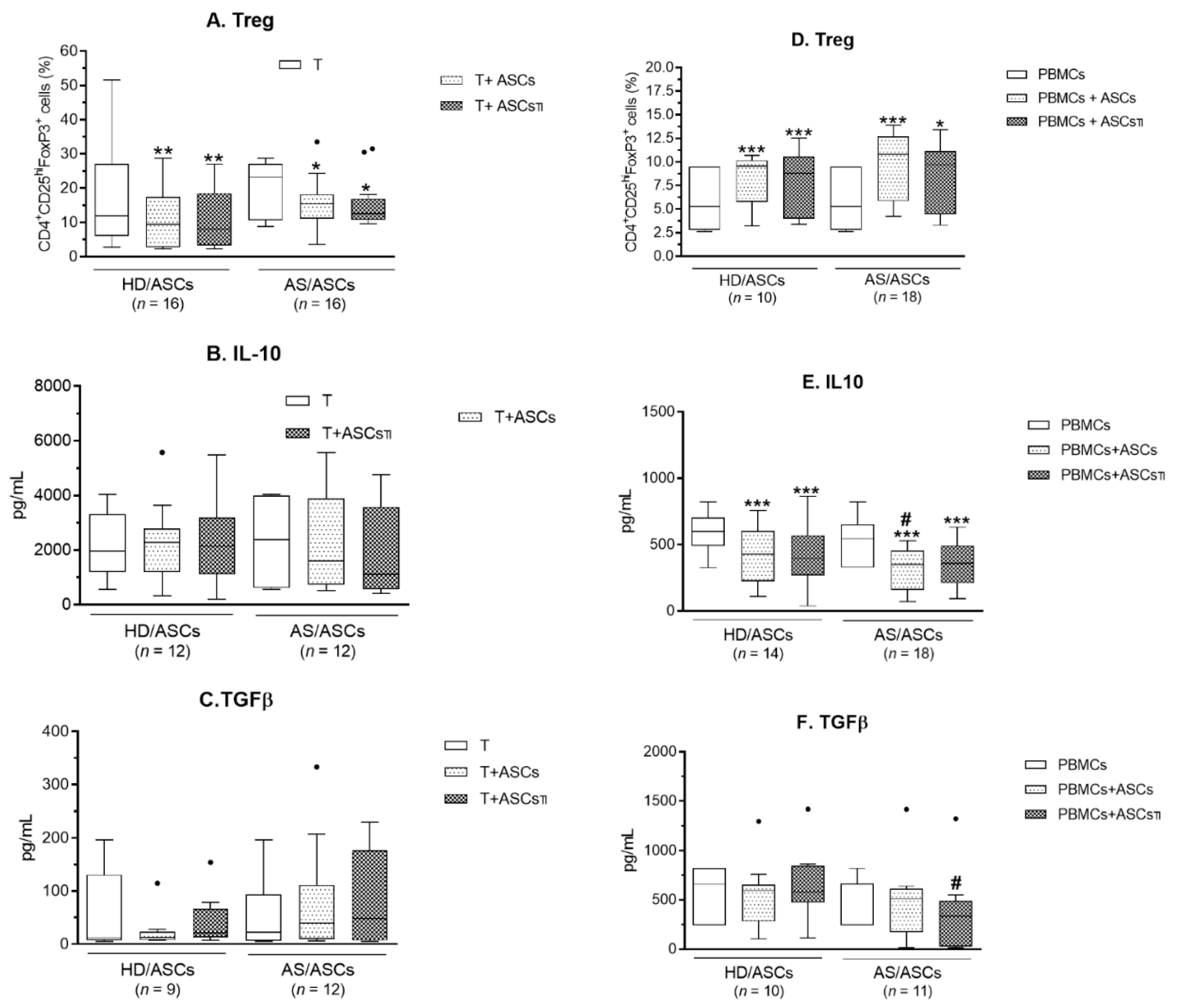
3.4. ASCs Modulate Generation of Classical Treg, But Fail to Up-Regulate Anti-Inflammatory Cytokines (IL-10 and TGFβ)
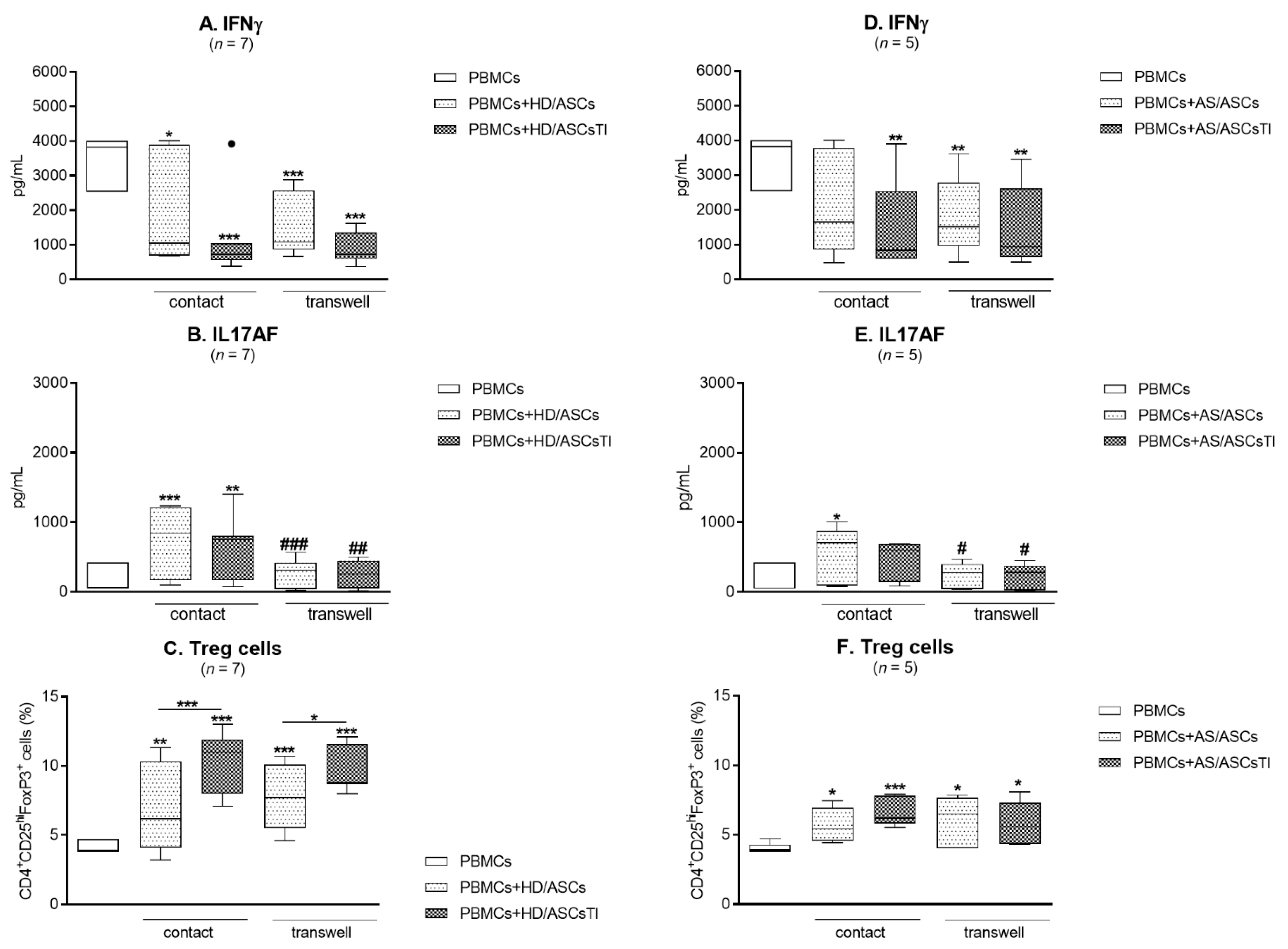
3.5. Contribution of the Cell-to-Cell Contact and Soluble Factors to ASCs Activity
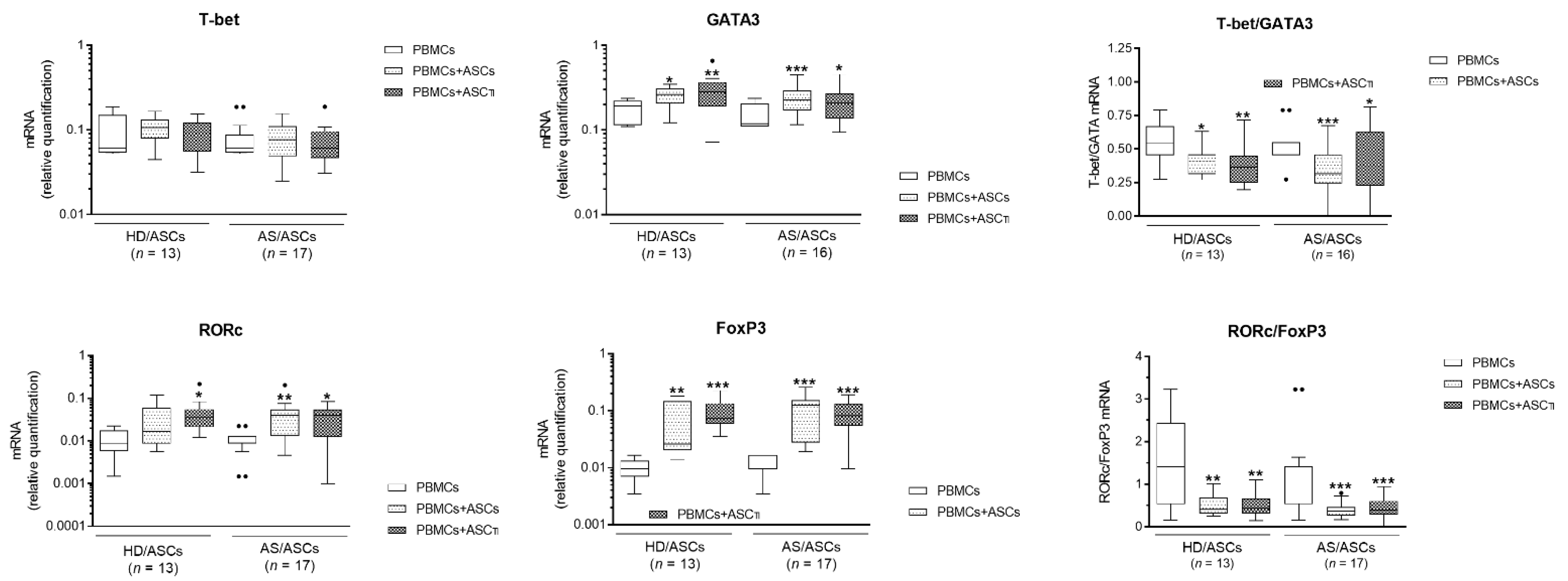
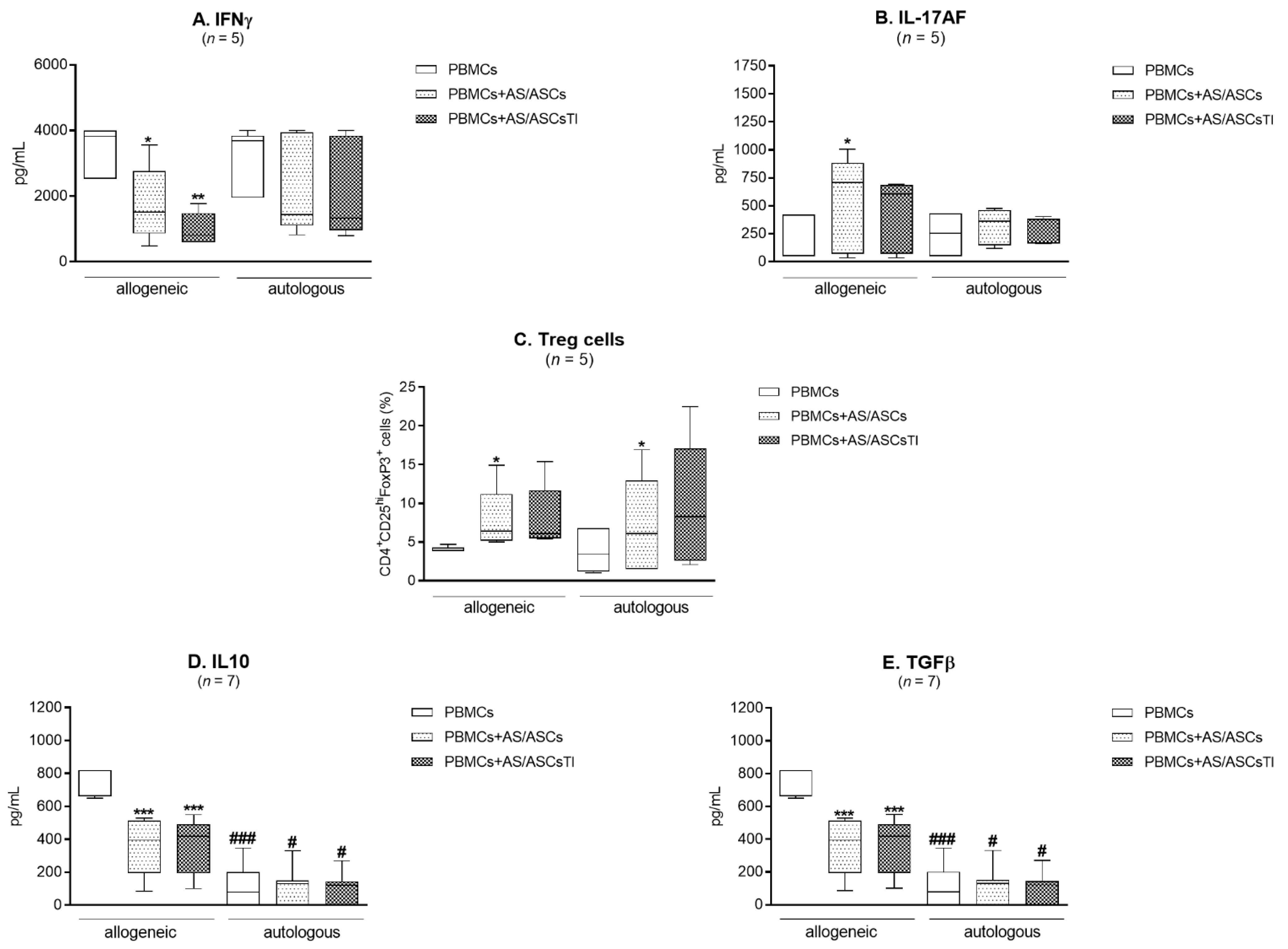
3.6. Comparison of AS/ASCs Effects in Allogeneic and Autologous System
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sieper, J.; Poddubny, D. Axial spondyloarthritis. Lancet 2017, 390, 73–84. [Google Scholar] [CrossRef]
- Cortes, A.; Pulit, S.L.; Leo, P.J.; Pointon, J.J.; Robinson, P.C.; Weisman, M.H.; Ward, M.; Gensler, L.S.; Zhou, X.; Garchon, H.-J.; et al. Major histocompatibility complex associations of ankylosing spondylitis are complex and involve further epistasis with ERAP1. Nat. Commun. 2015, 6, 7146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezaiemanesh, A.; Abdolmaleki, M.; Abdolmohammadi, K.; Aghaei, H.; Pakdel, F.D.; Fatahi, Y.; Soleimanifar, N.; Zavvar, M.; Nicknam, M.H. Immune cells involved in the pathogenesis of ankylosing spondylitis. Biomed. Pharmacother. 2018, 100, 98–204. [Google Scholar] [CrossRef] [PubMed]
- Toussirot, E.; Laheurte, C.; Gaugler, B.; Gabriel, D.; Saas, P. Increased IL-22 and IL-17A-producing mucosal-associated invariant T cells in the peripheral blood of patients with ankylosing spondylitis. Front. Immunol. 2018, 9, 1610. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, A.; Printz, I. Whodunit? The contribution of interleukin(IL)-17/IL-22-producing γδ T cells, αβ T cells, and innate lymphoid cells to the pathogenesis of spondyloarthritis. Front. Immunol. 2018, 9, 885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieper, J.; Deodhar, A.; Marzo-Ortega, H.; Aelion, J.A.; Blanco, R.; Jui-Cheng, T.; Anderson, M.; Porter, B.; Richards, H.B. Secukinumab efficacy in anti-TNF-naïve and anti-TNF-experienced subjects with active ankylosing spondylitis; results from the MEASURE 2 study. Ann. Rheum. Dis. 2017, 76, 571–592. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.T.; Zhang, D.H.; Fang, P.F.; Li, M.H.; Wang, R.J.; Li, S.H. Role of Th1/Th2 cytokines in the diagnosis and prognostic evaluation of ankylosing spondylitis. Genet. Mol. Res. 2017, 16. [Google Scholar] [CrossRef]
- Wang, C.; Liao, Q.; Hu, Y.; Zhong, D. T lymphocytes subset imbalances in patients contribute to ankylosing spondylitis. Exp. Ther. Med. 2015, 9, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Duan, Z.; Gui, Y.; Li, C.; Lin, J.; Gober, H.J.; Qin, J.; Li, D.; Wang, L. The immune dysfunction in ankylosing spondylitis patients. Biosci. Trends 2017, 11, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Jandus, C.; Bioley, G.; Rivals, J.P.; Dudler, J.; Speiser, D.; Romero, P. Increased numbers of circulating polyfunctional Th17 memory cells in patients with seronegative spondyloarthritides. Arthritis Rheum. 2008, 58, 2307–2317. [Google Scholar] [CrossRef]
- Miao, J.; Zhu, P. Functional defects of Treg cells: New targets in rheumatic diseases, including ankylosing spondylitis. Curr. Rheumatol. Rep. 2018, 20, 30. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Liu, C.; Bong, A.; Yang, J.; Zhou, X.; Wang, J.; Ji, B. Dysfunction of regulatory T cells in patients with ankylosing spondylitis is associated with a loss of Tim-3. Int. Immunopharmacol. 2018, 59, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Cagliani, J.; Grande, D.; Molmenti, E.P.; Miller, E.J.; Rilo, H.L.R. Immunomodulation by mesenchymal stromal cells and their clinical application. J. Stem Cell Regen. Biol. 2017, 3, 2–24. [Google Scholar] [CrossRef] [Green Version]
- Duffy, M.M.; Ritter, T.; Ceredig, R.; Griffin, M.D. Mesenchymal stem cell effects on T-cell effector pathways. Stem Cell Res. Ther. 2011, 2, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Wang, P.; Xie, Z.; Huang, L.; Yang, R.; Gao, L.; Tang, Y.; Zhang, X.; Ye, J.; Chen, K.; et al. Whole genome expression profiling and signal pathway screening of MSCs in ankylosing spondylitis. Stem Cells Int. 2014, 2014, 913050. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Wang, P.; Li, J.; Li, Y.; Wang, S.; Wu, X.; Sun, S.; Cen, S.; Su, H.; Deng, W.; et al. MCP1 triggers monocyte dysfunctions during abnormal osteogenic differentiation of mesenchymal stem cells in ankylosing spondylitis. J. Mol. Med. 2017, 95, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Ren, M.; Yang, R.; Liang, X.; Ma, Y.; Tang, Y.; Huang, L.; Ye, J.; Chen, K.; Wang, P.; et al. Reduced immunomodulation potential of bone marrow-derived mesenchymal stem cells induced CCR4+CCR6+ Th/Treg cell subset imbalance in ankylosing spondylitis. Arthritis Res. Ther. 2011, 13, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Krajewska-Włodarczyk, M.; Owczarczyk-Saczonek, A.; Placek, W.; Osowski, A.; Engelgardt, P.; Wojtkiewicz, J. Role of stem cells in pathophysiology and therapy of spondyloarthropathies—New therapeutic possibilities? Int. J. Mol. Sci. 2018, 19, 80. [Google Scholar] [CrossRef] [Green Version]
- Bright, B.; Bright, R.; Bright, L.; Limaye, A. Ankylosing spondylitis, chronic fatigue, and depression improve after stromal vascular fraction treatment for osteoarthritis: A case report. J. Med. Case Rep. 2018, 12, 238. [Google Scholar] [CrossRef]
- Kuca-Warnawin, E.; Skalska, U.; Janicka, I.; Musiałowicz, U.; Bonek, K.; Głuszko, P.; Szczęsny, P.; Olesińska, M.; Kontny, E. The phenotype and secretory activity of adipose-derived mesenchymal stem cells (ASCs) of patients with rheumatic diseases. Cells 2019, 8, 1659. [Google Scholar] [CrossRef] [Green Version]
- Kuca-Warnawin, E.; Janicka, I.; Szczęsny, P.; Olesińska, M.; Bonek, K.; Głuszko, P.; Kontny, E. Modulation of T-cell activation markers expression by the adipose tissue-derived mesenchymal stem cells of patients with rheumatic diseases. Cell Transplant. 2020, 29, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Rudwaleit, M.; van der Heijde, D.; Landewé, R.; Listing, J.; Akkoc, N.; Brandt, J.; Braun, J.; Chou, C.T.; Collantes-Estevez, E.; Dougados, M.; et al. The development of Assessment of SpondyloArthritis international Society classification criteria for axial spoandyloarthritis (part II): Validation and final selection. Ann. Rheum. Dis. 2009, 68, 777–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skalska, U.; Kontny, E.; Prochorec-Sobieszek, M.; Maslinski, W. Intra-articular adipose-derived mesenchymal stem cells from rheumatoid arthritis patients maintain the function of chondrogenic differentiation. Rheumatology (Oxford) 2012, 51, 1757–1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Paul, W.E. Peripheral CD4+ T-cell differentiation regulated by networks of cytokines and transcription factors. Immmunol. Rev. 2010, 238, 247–262. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.R. The balance of Th17 versus Treg cells in autoimmunity. Int. J. Mol. Sci. 2018, 19, 730. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Lv, Q.; Wei, Q.; Jiang, Y.; Qi, J.; Xiao, M.; Fang, L.; Xie, Y.; Cao, S.; Lin, Z.; et al. TNF-α inhibitor therapy can improve the immune imbalance of CD4+ T cells and negative regulatory cells but not CD8+ T cells in ankylosing spondylitis. Arthritis Res. Ther. 2020, 22, 149. [Google Scholar] [CrossRef]
- Shen, H.; Goodal, J.C.; Gaston, J.S.H. Frequency and phenotype of peripheral blood Th17 cells in ankylosing spondylitis and rheumatoid arthritis. Arthritis Rheum. 2009, 60, 1647–1656. [Google Scholar] [CrossRef]
- Fattahi, M.J.; Ahmadi, H.; Jafarnezhad-Ansariha, F.; Mortazavi-Jahromi, S.S.; Rehm, B.H.A.; Cuzzocrea, S.; Mastuo, H.; Mirshafiey, A. Oral administration effects of β-D-mannuronic acid (M2000) on Th17 and regulatory T cells in patients with ankylosing spondylitis. Biomed. Pharmacother. 2018, 100, 495–500. [Google Scholar] [CrossRef]
- Li, X.; Lina, C.; Zhenbiao, W.; Qing, H.; Qiang, L.; Zhu, P. Levels of circulating Th17 cells and regulatory T cells in ankylosing spondylitis patients with an inadequate response to anti- TNF –α therapy. J. Clin. Immunol. 2013, 33, 151–161. [Google Scholar]
- Adams, R.; Maroof, A.; Baker, T.; Lawson, A.D.G.; Oliver, R.; Paveley, R.; Rapecki, S.; Shaw, S.; Vajjah, P.; West, S.; et al. Bimekizumab, a novel humanized IgG1 antibody that neutralizes both IL-17A and IL-17F. Front. Immunol. 2020, 11, 1894. [Google Scholar] [CrossRef]
- Bi, Y.; Lin, X.; Liang, H.; Yang, D.; Ke, J.; Xiao, J.; Chen, Z.; Chen, W.; Zhang, X.; Wang, S.; et al. Human adipose tissue-derived mesenchymal stem cells in Parkinson’s Disease: Inhibition of T helper 17 cell differentiation and regulation of immune balance towards a regulatory cell phenotype. Clin. Interv. Aging 2020, 15, 1383–1391. [Google Scholar] [CrossRef] [PubMed]
- Fasching, P.; Stadner, M.; Graninger, W.; Dejaco, C.; Fessler, J. Therapeutic potential of targeting the Th17/Treg axis in autoimmune disorders. Molecules 2017, 22, 134. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.M.; Jenner, R.G. Transcriptional factor interplay in T helper cell differentiation. Brief. Funct. Genom. 2013, 12, 499–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.R. Transcriptional regulation of T helper type 2 differentiation. Immunology 2013, 141, 498–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Lin, F.; Gao, Y.; Li, Z.; Zhang, J.; Xing, Y.; Deng, Z.; Yao, Z.; Tsun, A.; Li, B. FoxP3 and RORγτ: Transcriptional regulation of Treg and Th17. Int. Immunopharmacol. 2011, 11, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, P.; Charbonnier, L.M.; Chatila, T.A. Regulatory T cells: The many faces of FoxP3. J. Clin. Immunol. 2019, 39, 623–640. [Google Scholar] [CrossRef]
- Seo, Y.; Shin, T.H.; Kim, H.S. Current strategies to enhance adipose stem cell function: An update. Int. J. Mol. Sci. 2019, 20, 3827. [Google Scholar] [CrossRef] [Green Version]
- Najar, M.; Lombard, C.A.; Fayyad-Kazan, H.; Faour, W.H.; Merimi, M.; Sokal, E.M.; Lagneaux, L.; Fahmi, H. Th17 immune response to adipose tissue-derived mesenchymal stromal cells. J. Cell Physiol. 2019, 234, 21145–21152. [Google Scholar] [CrossRef] [Green Version]
- Najar, M.; Fayyad-Kazan, H.; Faour, W.H.; Merimi, M.; Sokal, E.M.; Lombard, C.A.; Fahmi, H. Immunological modulation following bone marrow-derived mesenchymal stromal cells and Th17 lymphocyte co-cultures. Inflamm. Res. 2019, 68, 203–213. [Google Scholar] [CrossRef]
- Maseda, D.; Johnson, E.M.; Nyhoff, L.E.; Baron, E.; Kojima, F.; Wilhelm, A.J.; Ward, M.R.; Woodward, J.G.; Brand, D.D.; Crofford, L.J. mPGES1-dependent prostaglandin E2 (PGE2) controls antigen-specific Th17 and Th1 responses by regulating T autocrine and paracrine PGE2 production. J. Immunol. 2018, 200, 725–736. [Google Scholar] [CrossRef] [Green Version]
- Mahnke, J.; Schumacher, V.; Ahrens, S.; Käding, N.; Feldhoff, L.M.; Huber, M.; Rupp, J.; Raczkowski, F.; Mittrücker, H.W. Interferon regulatory factor 4 controls Th1 cell effector function and metabolism. Sci. Rep. 2016, 6, 35521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.; Zheng, C.; Chen, Z.; Gu, D.; Du, W.; Ge, J.; Ha, Z.; Yang, R. Fetal BM-derived mesenchymal stem cells promote the expansion of human Th17 cells, but inhibit the production of Th1 cells. Eur. J. Immunol. 2009, 39, 2840–2849. [Google Scholar] [CrossRef] [PubMed]
- Carriòn, F.; Nova, E.; Luz, P.; Apablaza, F.; Figueroa, F. Opposing effect of mesenchymal stem cells on Th1 and Th17 cell polarization according to the state of CD4+ T cell activation. Immunol. Lett. 2011, 135, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Darlington, P.J.; Boivin, M.N.; Renoux, C.; Franҫois, M.; Galipeau, J.; Freedman, M.S.; Atkins, H.L.; Cohen, J.A.; Solchaga, L.; Bar-Or, A. Reciprocal Th1 and Th17 regulation by mesenchymal stem cells: Implication for multiple sclerosis. Ann. Neurol. 2010, 68, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Rozenberg, A.; Rezk, A.; Boivin, M.N.; Darlington, P.J.; Nyirenda, M.; Li, R.; Jalili, F.; Winer, R.; Artsy, E.A.; Uccelli, A. Human mesenchymal stem cells impact Th17 and Th1 responses through a prostaglandin E2 and myeloid-dependent mechanism. Stem Cells Transl. Med. 2016, 5, 1506–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melief, S.M.; Schrama, E.; Brugman, M.H.; Tiemessen, M.M.; Hoogduijn, M.J.; Fibbe, W.E.; Roelofs, H. Multipotent stromal cells induce human regulatory T cells through a novel pathway involving skewing of monocytes toward anti-inflammatory macrophages. Stem Cells 2013, 31, 1980–1991. [Google Scholar] [CrossRef]
- Luz-Crawford, P.; Kurte, M.; Bravo-Alegria, J.; Contreras, R.; Nova-Lamperti, E.; Tejedor, G.; Noël, D.; Jorgensen, C.; Figueroa, F.; Djouad, F. Mesenchymal stem cells generate a CD4+CD25+ Foxp3+ regulatory T cell population during the differentiation process of Th1 and Th17 cells. Stem Cell Res. Ther. 2013, 4, 65. [Google Scholar] [CrossRef] [Green Version]
- Luz-Crawford, P.; Noël, D.; Fernandez, X.; Khoury, M.; Figueroa, F.; Carriòn, F.; Jorgense, C.; Djouad, F. Mesenchymal stem cells repress Th17 molecular program through the PD-1 pathway. PLoS ONE 2012, 7, e45272. [Google Scholar] [CrossRef]
- Laranjeira, P.; Pedrosa, M.; Pedreiro, S.; Gomes, J.; Martinho, A.; Antunes, B.; Ribeiro, T.; Santos, F.; Trindade, H.; Paiva, A. Effect of human bone marrow mesenchymal stromal cells on cytokine production by peripheral blood naïve, memory, and effector T cells. Stem Cell Res. Ther. 2015, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Ghannam, S.; Péne, J.; Moquet-Torcy, G.; Jorgensen, C.; Yssel, H. Mesenchymal stem cells inhibit human Th17 cell differentiation and function and induce a T regulatory cell phenotype. J. Immunol. 2010, 185, 302–312. [Google Scholar] [CrossRef] [Green Version]
- Duffy, M.M.; Pindjakova, J.; Hanley, S.A.; McCarthy, C.; Weidhofer, G.A.; Sweeney, E.M.; English, K.; Shaw, G.; Murphy, J.M.; Barry, F.P. Mesenchymal stem cell inhibition of T-helper 17 cell-differentiation is triggered by cell-cell contact and mediated by prostaglandin E2 via EP4 receptor. Eur. J. Immunol. 2011, 41, 2840–2851. [Google Scholar] [CrossRef] [PubMed]
- Tatara, R.; Ozaki, K.; Kikuchi, Y.; Hatanaka, K.; Oh, I.; Meguro, A.; Matsu, H.; Sat, K.; Ozawa, K. Mesenchymal stromal cells inhibit Th17 but not regulatory T-cell differentiation. Cytotherapy 2011, 13, 686–694. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Zhang, J.; Gu, Q.; Huang, M.; Zhang, W.; Guo, J.; Zhou, X. Reciprocal expression of IL-35 and IL-10 defines two distinct effector Treg subsets that are required for maintenance of immune tolerance. Cell Rep. 2017, 21, 1853–1869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roncarolo, M.G.; Gregori, S.; Bacchetta, R.; Battaglia, M.; Gagliani, N. The biology of T regulatory type 1 cells and their therapeutic application in immune-mediated diseases. Immunity 2018, 49, 1004–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova-Todorova, E.; Bochev, I.; Dimitrov, R.; Bolemezova, K.; Mourdjeva, M.; Kyurkchiev, S.; Kinov, P.; Altankova, I.; Kyurkchiev, D. Conditioned medium from adipose tissue-derived mesenchymal stem cells induces CD4+FOXP3+ cells and increases IL-10 secretion. J. Biomed. Biotechnol. 2012, 2012, 295167. [Google Scholar] [CrossRef] [PubMed]
- Engela, A.U.; Hoogduijn, M.J.; Boer, K.; Litjens, N.H.R.; Betjes, M.G.H.; Weimar, W.; Baan, C.C. Human adipose-tissue derived mesenchymal stem cells induce functional de novo regulatory T cells with methylated FOXP3 gene DNA. Clin. Exp. Immunol. 2013, 173, 343–354. [Google Scholar] [CrossRef]
- Hsu, S.C.; Wang, L.T.; Yao, C.L.; Lai, H.Y.; Chan, K.Y.; Liu, B.S.; Chong, P.; Lee, O.K.S.; Chen, H.W. Mesenchymal stem cells promote neutrophil activation by inducing IL-17 production in CD4+ CD45RO+ T cells. Immunobiology 2013, 218, 90–95. [Google Scholar] [CrossRef]
- Seddiki, N.; Santner-Nanan, B.; Martison, J.; Zaunders, J.; Sasson, S.; Launday, A.; Solomon, M.; Selby, W.; Alexander, S.J.; Nanan, R. Expression of interleukin (IL)-2 and IL-7 receptors discriminates between human regulatory and activated T cells. J. Exp. Med. 2006, 203, 1693–1700. [Google Scholar] [CrossRef]
- Kastrianaki, M.C.; Sidiropoulos, P.; Roche, S.; Ringe, J.; Lehmann, S.; Kritikos, H.; Vlaha, V.M.; Delorme, B.; Eliopoulos, G.D.; Jorgensen, C. Functional, molecular and proteomic characterisation of bone marrow mesenchymal stem cells in rheumatoid arthritis. Ann. Rheum. Dis. 2008, 67, 741–749. [Google Scholar] [CrossRef] [Green Version]
- Müller, M.; Raabe, O.; Addicks, K.; Wenisch, S.; Arnhold, S. Effects of non-steroidal anti-inflammatory drugs on proliferation, differentiation and migration in equine mesenchymal stem cells. Cell Biol. Int. 2011, 35, 235–248. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Ankylosing Spondylitis (AS) (n = 21) |
|---|---|
| Demographics | |
| Age, years | 43.5 ± 2.79 |
| Sex, female (F)/male (M), n | 9 F/12 M |
| Disease duration, years | 7.53 ± 0.98 |
| Clinical data | |
| BASDAI, score | 5.6 ± 0.48 |
| ASDASCRP, score | 3.42 ± 0.22 |
| BASFI, score | 4.72 ± 0.61 |
| BASMI, score | 3.99 ± 0.43 |
| HAQ, score | 1.1 ± 0.17 |
| Laboratory values | |
| CRP, mg/L | 13.25 ± 3.38 |
| ESR, mm/h | 20.8 ± 4.13 |
| Medications, % | |
| NSAIDs | 90.0 |
| Non-biologic DMARDs | 25.0 |
| Glucocorticosteroids | 15.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuca-Warnawin, E.; Janicka, I.; Bonek, K.; Kontny, E. Modulatory Impact of Adipose-Derived Mesenchymal Stem Cells of Ankylosing Spondylitis Patients on T Helper Cell Differentiation. Cells 2021, 10, 280. https://doi.org/10.3390/cells10020280
Kuca-Warnawin E, Janicka I, Bonek K, Kontny E. Modulatory Impact of Adipose-Derived Mesenchymal Stem Cells of Ankylosing Spondylitis Patients on T Helper Cell Differentiation. Cells. 2021; 10(2):280. https://doi.org/10.3390/cells10020280
Chicago/Turabian StyleKuca-Warnawin, Ewa, Iwona Janicka, Krzysztof Bonek, and Ewa Kontny. 2021. "Modulatory Impact of Adipose-Derived Mesenchymal Stem Cells of Ankylosing Spondylitis Patients on T Helper Cell Differentiation" Cells 10, no. 2: 280. https://doi.org/10.3390/cells10020280