
Pyk2 Stabilizes Striatal Medium Spiny Neuron Structure and Striatal-Dependent Action




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Subjects
2.2. Golgi-Cox Staining
2.3. Neuron Imaging
2.4. Plasmids
2.5. Cell Culture, Transfection, and Viral Vector preparation
2.6. Western Blot Analysis of Cell Culture and Brain Samples
2.7. Viral Vector Delivery
2.8. Behavioral Testing
2.8.1. Food-Reinforced Response Training
2.8.2. Test for Behavioral Response Flexibility
2.9. Clozapine N-Oxide (CNO) Administration and Experimental Design
2.10. Locomotor Activity
2.11. Histological Verification of Viral Vector Placement
2.12. Statistical Analysis
3. Results
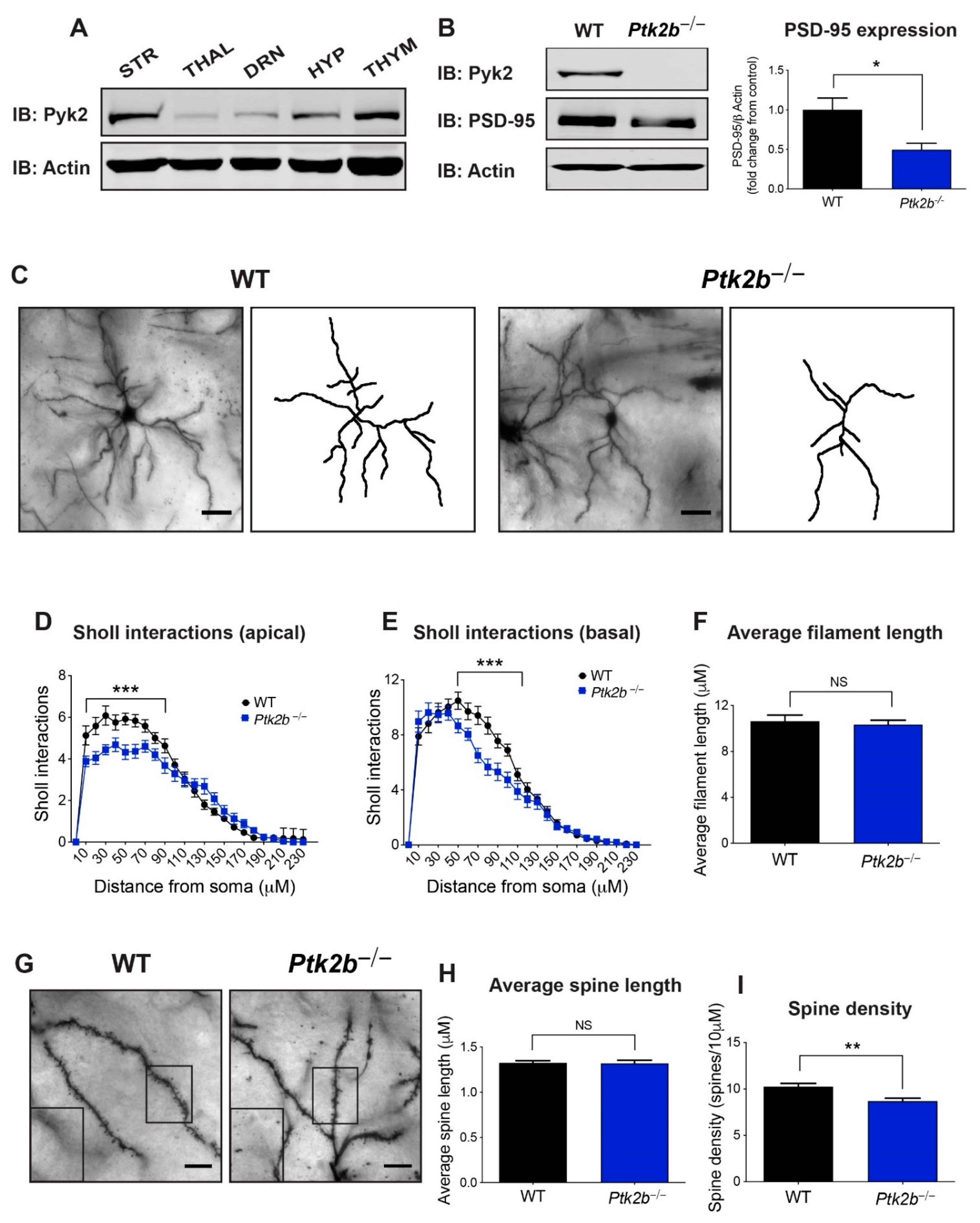
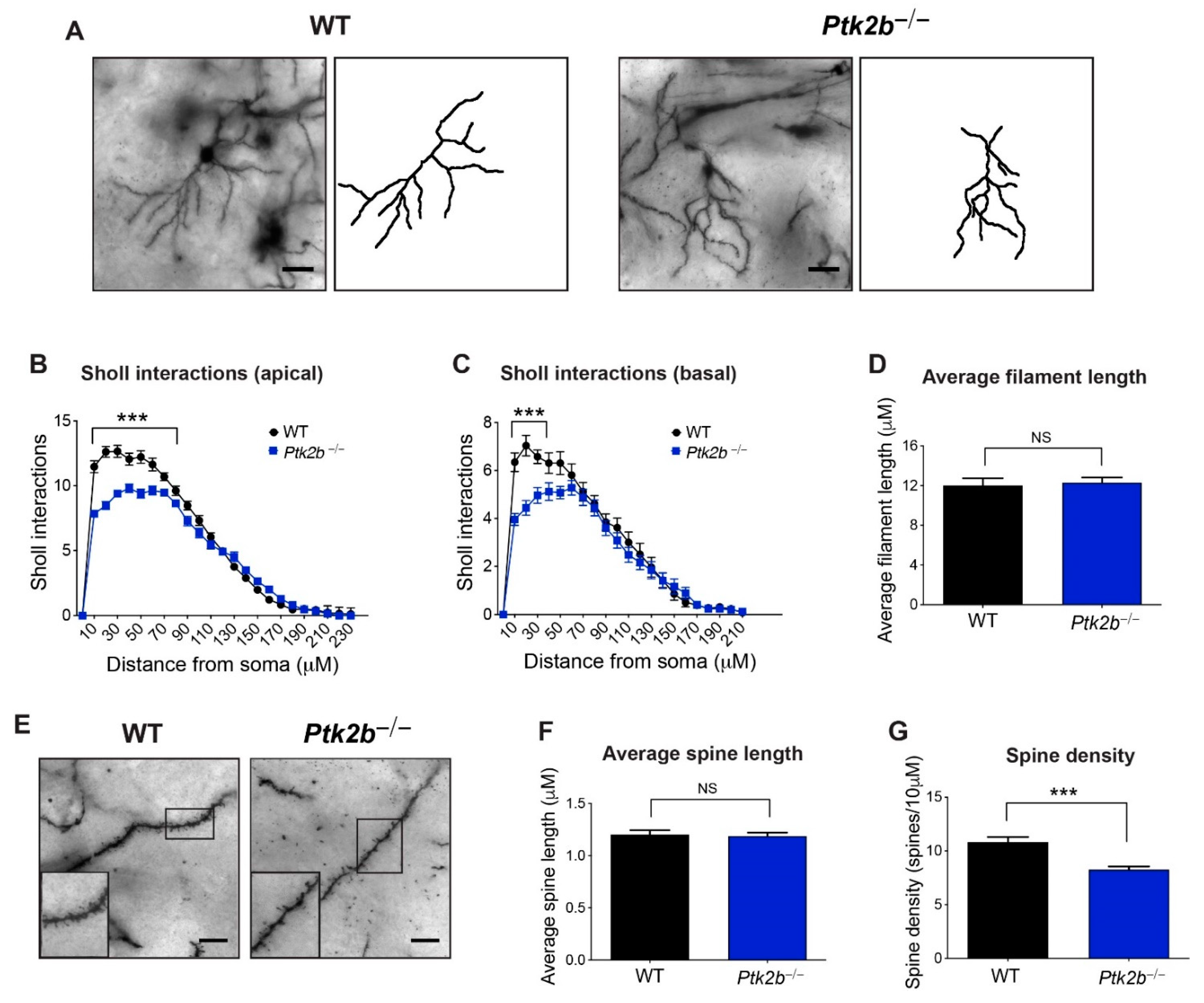
3.1. Pyk2 Is Highly Expressed in the Striatum, Where It Controls Dendrite Arborization and Spine Density on Striatal MSNs
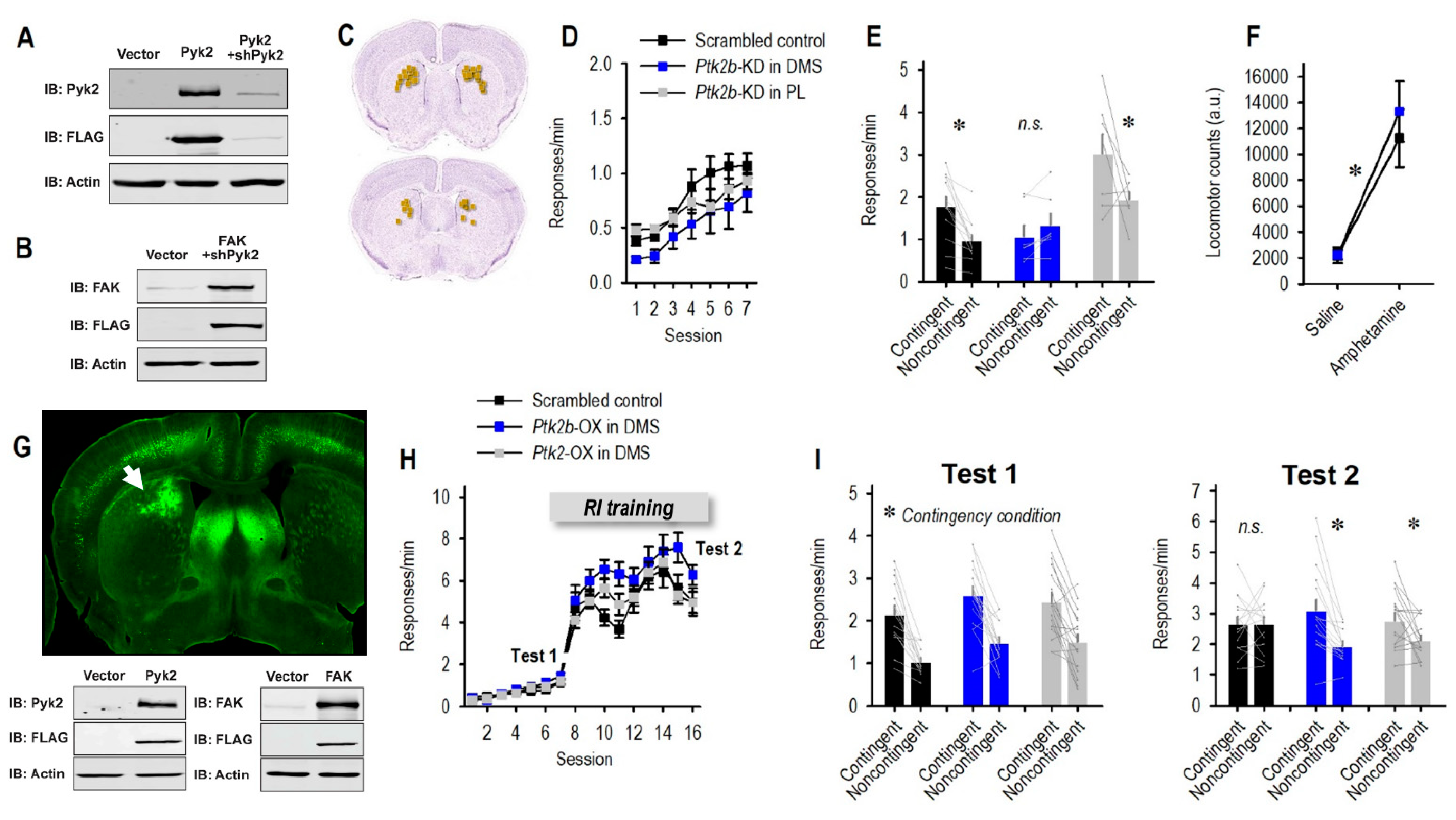
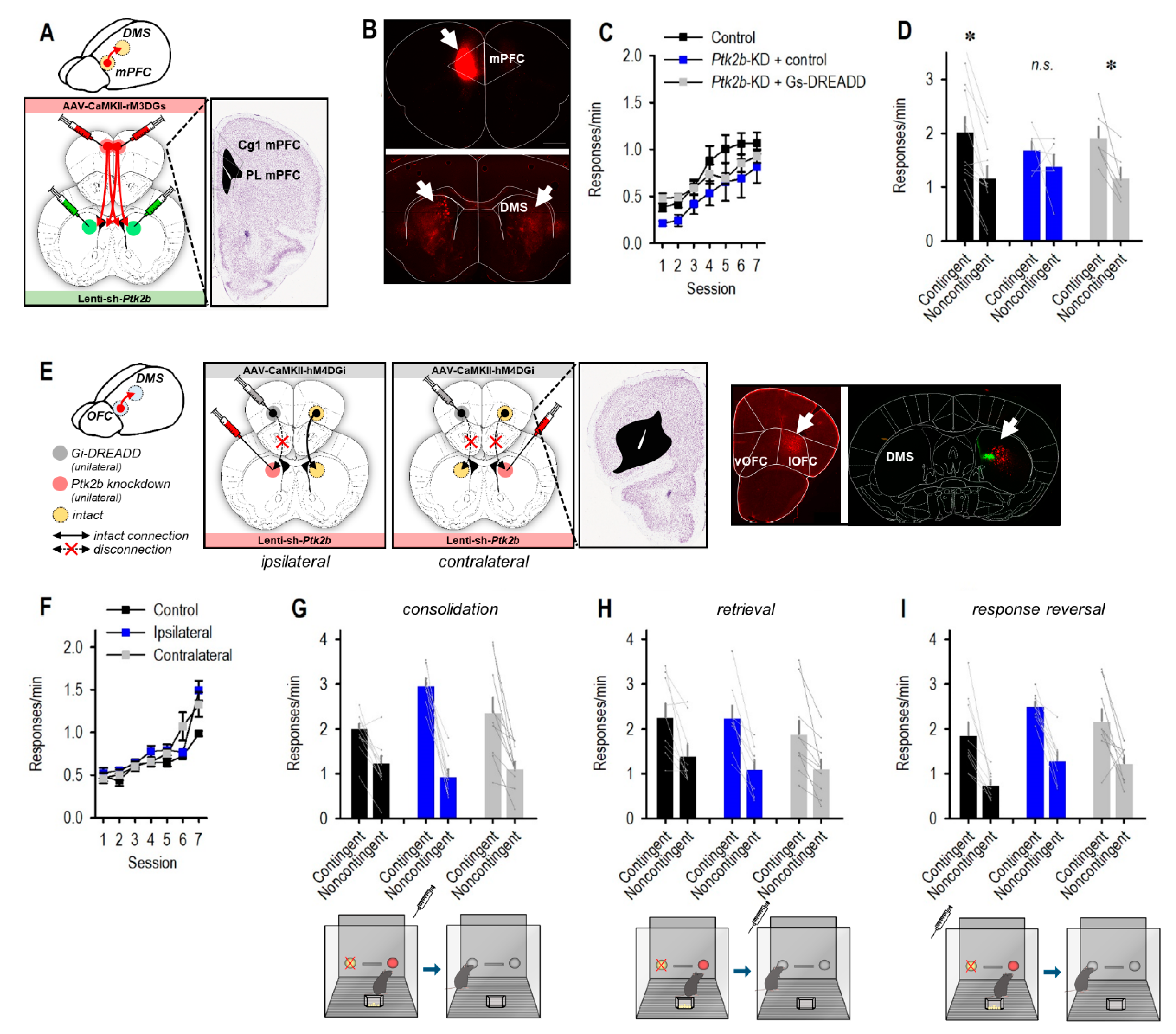
3.2. Pyk2 Supports Behavioral Flexibility
3.3. Enrichment of Striatal Pyk2 or FAK Improves Action Flexibility
3.4. mPFC Stimulation Reinstates Flexible Behavior following Striatal Pyk2 Loss
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Braun, S.; Hauber, W. Striatal dopamine depletion in rats produces variable effects on contingency detection: Task-related influences. Eur. J. Neurosci. 2012, 35, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Pauli, W.M.; Clark, A.D.; Guenther, H.J.; O’Reilly, R.C.; Rudy, J.W. Inhibiting PKMzeta reveals dorsal lateral and dorsal medial striatum store the different memories needed to support adaptive behavior. Learn. Mem. 2012, 19, 307–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradfield, L.A.; Bertran-Gonzalez, J.; Chieng, B.; Balleine, B.W. The thalamostriatal pathway and cholinergic control of goal-directed action: Interlacing new with existing learning in the striatum. Neuron 2013, 79, 153–166. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, A.; Tan, C.; Mettlach, G.; Pozo, K.; Plattner, F.; Bibb, J.A. Cdk5 Modulates Long-Term Synaptic Plasticity and Motor Learning in Dorsolateral Striatum. Sci. Rep. 2016, 6, 29812. [Google Scholar] [CrossRef] [Green Version]
- Maroteaux, M.; Valjent, E.; Longueville, S.; Topilko, P.; Girault, J.A.; Herve, D. Role of the plasticity-associated transcription factor zif268 in the early phase of instrumental learning. PLoS ONE 2014, 9, e81868. [Google Scholar] [CrossRef]
- Matamales, M.; McGovern, A.E.; Dai Mi, J.; Mazzone, S.B.; Balleine, B.W.; Bertran-Gonzalez, J. Local D2- to D1-neuron transmodulation updates goal-directed learning in the striatum. Science 2020, 367, 549–555. [Google Scholar] [CrossRef]
- Yin, H.H.; Mulcare, S.P.; Hilario, M.R.; Clouse, E.; Holloway, T.; Davis, M.I.; Hansson, A.C.; Lovinger, D.M.; Costa, R.M. Dynamic reorganization of striatal circuits during the acquisition and consolidation of a skill. Nat. Neurosci. 2009, 12, 333–341. [Google Scholar] [CrossRef] [Green Version]
- Hart, G.; Bradfield, L.A.; Fok, S.Y.; Chieng, B.; Balleine, B.W. The Bilateral Prefronto-striatal Pathway Is Necessary for Learning New Goal-Directed Actions. Curr. Biol. 2018, 28, 2218–2229.e2217. [Google Scholar] [CrossRef] [Green Version]
- Gremel, C.M.; Chancey, J.H.; Atwood, B.K.; Luo, G.; Neve, R.; Ramakrishnan, C.; Deisseroth, K.; Lovinger, D.M.; Costa, R.M. Endocannabinoid Modulation of Orbitostriatal Circuits Gates Habit Formation. Neuron 2016, 90, 1312–1324. [Google Scholar] [CrossRef] [Green Version]
- Gremel, C.M.; Costa, R.M. Orbitofrontal and striatal circuitsdynamically encode the shift between goal-directed and habitual actions. Nat. Commun. 2013, 4, 2264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caceda, R.; Nemeroff, C.B.; Harvey, P.D. Toward an Understanding of Decision Making in Severe Mental Illness. J. Neuropsychiatry Clin. Neurosci. 2014, 26, 196–213. [Google Scholar] [CrossRef]
- Giralt, A.; Brito, V.; Chevy, Q.; Simonnet, C.; Otsu, Y.; Cifuentes-Diaz, C.; de Pins, B.; Coura, R.; Alberch, J.; Gines, S.; et al. Pyk2 modulates hippocampal excitatory synapses and contributes to cognitive deficits in a Huntington’s disease model. Nat. Commun. 2017, 8, 15592. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, K.D.; Meirson, T.; Sams, D.S.; Gil-Henn, H. FAK family kinases in brain health and disease. J. Mol. Clin. Med. 2018, 1, 177–190. [Google Scholar] [CrossRef]
- Menegon, A.; Burgaya, F.; Baudot, P.; Dunlap, D.D.; Girault, J.-A.; Valtorta, F. FAK+ and PYK2/CAKβ, two related tyrosine kinases highly expressed in the central nervous system: Similarities and differences in the expression pattern. Eur. J. Neurosci. 1999, 11, 3777–3788. [Google Scholar] [CrossRef]
- Okigaki, M.; Davis, C.; Falasca, M.; Harroch, S.; Felsenfeld, D.P.; Sheetz, M.P.; Schlessinger, J. Pyk2 regulates multiple signaling events crucial for macrophage morphology and migration. Proc. Natl. Acad. Sci. USA 2003, 100, 10740–10745. [Google Scholar] [CrossRef] [Green Version]
- Feng, G.; Mellor, R.H.; Bernstein, M.; Keller-Peck, C.; Nguyen, Q.T.; Wallace, M.; Nerbonne, J.M.; Lichtman, J.W.; Sanes, J.R. Imaging Neuronal Subsets in Transgenic Mice Expressing Multiple Spectral Variants of GFP. Neuron 2000, 28, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Urban, D.J.; Roth, B.L. DREADDs (Designer Receptors Exclusively Activated by Designer Drugs): Chemogenetic Tools with Therapeutic Utility. Annu. Rev. Pharmacol. Toxicol. 2015, 55, 399–417. [Google Scholar] [CrossRef] [PubMed]
- Ehrengruber, M.U.; Hennou, S.; Büeler, H.; Naim, H.Y.; Déglon, N.; Lundstrom, K. Gene transfer into neurons from hippocampal slices: Comparison of recombinant Semliki Forest Virus, adenovirus, adeno-associated virus, lentivirus, and measles virus. Mol. Cell. Neurosci. 2001, 17, 855–871. [Google Scholar] [CrossRef]
- Gourley, S.L.; Zimmermann, K.S.; Allen, A.G.; Taylor, J.R. The Medial Orbitofrontal Cortex Regulates Sensitivity to Outcome Value. J. Neurosci. 2016, 36, 4600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whyte, A.J.; Trinoskey-Rice, G.; Davies, R.A.; Woon, E.P.; Foster, S.L.; Shapiro, L.P.; Li, D.C.; Srikanth, K.D.; Gil-Henn, H.; Gourley, S.L. Cell adhesion factors in the orbitofrontal cortex control cue-induced reinstatement of cocaine seeking and amygdala-dependent goal seeking. J. Neurosci. 2021, 41, 5923–5936. [Google Scholar] [CrossRef]
- Gourley, S.L.; Swanson, A.M.; Jacobs, A.M.; Howell, J.L.; Mo, M.; DiLeone, R.J.; Koleske, A.J.; Taylor, J.R. Action control is mediated by prefrontal BDNF and glucocorticoid receptor binding. Proc. Natl. Acad. Sci. USA 2012, 109, 20714–20719. [Google Scholar] [CrossRef] [Green Version]
- Barker, J.; Torregrossa, M.; Taylor, J. Bidirectional modulation of infralimbic dopamine D1 and D2 receptor activity regulates flexible reward seeking. Front. Neurosci. 2013, 7, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson, A.M.; DePoy, L.M.; Gourley, S.L. Inhibiting Rho kinase promotes goal-directed decision making and blocks habitual responding for cocaine. Nat. Commun. 2017, 8, 1861. [Google Scholar] [CrossRef] [Green Version]
- Whyte, A.J.; Kietzman, H.W.; Swanson, A.M.; Butkovich, L.M.; Barbee, B.R.; Bassell, G.J.; Gross, C.; Gourley, S.L. Reward-Related Expectations Trigger Dendritic Spine Plasticity in the Mouse Ventrolateral Orbitofrontal Cortex. J. Neurosci. 2019, 39, 4595–4605. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, K.S.; Li, C.C.; Rainnie, D.G.; Ressler, K.J.; Gourley, S.L. Memory Retention Involves the Ventrolateral Orbitofrontal Cortex: Comparison with the Basolateral Amygdala. Neuropsychopharmacology 2018, 43, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, K.S.; Yamin, J.A.; Rainnie, D.G.; Ressler, K.J.; Gourley, S.L. Connections of the Mouse Orbitofrontal Cortex and Regulation of Goal-Directed Action Selection by Brain-Derived Neurotrophic Factor. Biol. Psychiatry 2017, 81, 366–377. [Google Scholar] [CrossRef] [Green Version]
- Gomez, J.L.; Bonaventura, J.; Lesniak, W.; Mathews, W.B.; Sysa-Shah, P.; Rodriguez, L.A.; Ellis, R.J.; Richie, C.T.; Harvey, B.K.; Dannals, R.F.; et al. Chemogenetics revealed: DREADD occupancy and activation via converted clozapine. Science 2017, 357, 503–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, J.M.; Bryant, K.G.; Chandler, L.J. Inactivation of ventral hippocampus projections promotes sensitivity to changes in contingency. Learn. Mem. 2019, 26, 1–8. [Google Scholar] [CrossRef] [Green Version]
- De Pins, B.; Montalban, E.; Vanhoutte, P.; Giralt, A.; Girault, J.A. Pyk2 in D1 receptor-expressing neurons of the nucleus accumbens modulates the acute locomotor effects of cocaine. BioRxiv 2019. [Google Scholar] [CrossRef]
- Brunzell, D.H.; Russell, D.S.; Picciotto, M.R. In vivo nicotine treatment regulates mesocorticolimbic CREB and ERK signaling in C57Bl/6J mice. J. Neurochem. 2003, 84, 1431–1441. [Google Scholar] [CrossRef]
- Sheehan, T.P.; Neve, R.L.; Duman, R.S.; Russell, D.S. Antidepressant effect of the calcium-activated tyrosine kinase Pyk2 in the lateral septum. Biol. Psychiatry 2003, 54, 540–551. [Google Scholar] [CrossRef]
- De Pins, B.; Montalban, E.; Vanhoutte, P.; Giralt, A.; Girault, J.-A. The non-receptor tyrosine kinase Pyk2 modulates acute locomotor effects of cocaine in D1 receptor-expressing neurons of the nucleus accumbens. Sci. Rep. 2020, 10, 6619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Wit, S.; Dickinson, A. Associative theories of goal-directed behaviour: A case for animal-human translational models. Psychol. Res. 2009, 73, 463–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwabe, L.; Wolf, O.T. Stress and multiple memory systems: From ‘thinking’ to ‘doing’. Trends Cogn. Sci. 2013, 17, 60–68. [Google Scholar] [CrossRef]
- Gourley, S.L.; Taylor, J.R. Going and stopping: Dichotomies in behavioral control by the prefrontal cortex. Nat. Neurosci. 2016, 19, 656–664. [Google Scholar] [CrossRef] [Green Version]
- Everitt, B.J.; Robbins, T.W. Drug Addiction: Updating Actions to Habits to Compulsions Ten Years On. Annu. Rev. Psychol. 2016, 67, 23–50. [Google Scholar] [CrossRef]
- Huang, Y.; Lu, W.; Ali, D.W.; Pelkey, K.A.; Pitcher, G.M.; Lu, Y.M.; Aoto, H.; Roder, J.C.; Sasaki, T.; Salter, M.W.; et al. CAKbeta/Pyk2 kinase is a signaling link for induction of long-term potentiation in CA1 hippocampus. Neuron 2001, 29, 485–496. [Google Scholar] [CrossRef] [Green Version]
- Polis, B.; Gil-Henn, H. Commentary on Giralt et al.: PTK2B/Pyk2 overexpression improves a mouse model of Alzheimer’s disease. Exp. Neurol. 2019, 311, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Salazar, S.V.; Cox, T.O.; Strittmatter, S.M. Pyk2 Signaling through Graf1 and RhoA GTPase Is Required for Amyloid-β Oligomer-Triggered Synapse Loss. J. Neurosci. 2019, 39, 1910–1929. [Google Scholar] [CrossRef] [PubMed]
- Salazar, S.V.; Cox, T.O.; Lee, S.; Brody, A.H.; Chyung, A.S.; Haas, L.T.; Strittmatter, S.M. Alzheimer’s Disease Risk Factor Pyk2 Mediates Amyloid-β-Induced Synaptic Dysfunction and Loss. J. Neurosci. 2019, 39, 758–772. [Google Scholar] [CrossRef] [Green Version]
- Pitts, E.G.; Li, D.C.; Gourley, S.L. Bidirectional coordination of actions and habits by TrkB in mice. Sci. Rep. 2018, 8, 4495. [Google Scholar] [CrossRef] [Green Version]
- Gourley, S.L.; Olevska, A.; Gordon, J.; Taylor, J.R. Cytoskeletal Determinants of Stimulus-Response Habits. J. Neurosci. 2013, 33, 11811–11816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monje, F.J.; Kim, E.J.; Pollak, D.D.; Cabatic, M.; Li, L.; Baston, A.; Lubec, G. Focal Adhesion Kinase Regulates Neuronal Growth, Synaptic Plasticity and Hippocampus-Dependent Spatial Learning and Memory. Neurosignals 2012, 20, 1–14. [Google Scholar] [CrossRef]
- Woon, E.P.; Sequeira, M.K.; Barbee, B.R.; Gourley, S.L. Involvement of the rodent prelimbic and medial orbitofrontal cortices in goal-directed action: A brief review. J. Neurosci. Res. 2020, 98, 1020–1030. [Google Scholar] [CrossRef] [PubMed]
- Stalnaker, T.A.; Cooch, N.K.; Schoenbaum, G. What the orbitofrontal cortex does not do. Nat. Neurosci. 2015, 18, 620–627. [Google Scholar] [CrossRef]
- Jackson, S.A.; Horst, N.K.; Pears, A.; Robbins, T.W.; Roberts, A.C. Role of the Perigenual Anterior Cingulate and Orbitofrontal Cortex in Contingency Learning in the Marmoset. Cereb. Cortex 2016, 26, 3273–3284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, L.Y.; Horst, N.K.; Cranmore, S.A.W.; Horiguchi, N.; Cardinal, R.N.; Roberts, A.C.; Robbins, T.W. Controlling one’s world: Identification of sub-regions of primate PFC underlying goal-directed behavior. Neuron 2021, 109, 2485–2498.e2485. [Google Scholar] [CrossRef] [PubMed]
- Burguière, E.; Monteiro, P.; Feng, G.; Graybiel, A.M. Optogenetic stimulation of lateral orbitofronto-striatal pathway suppresses compulsive behaviors. Science 2013, 340, 1243–1246. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.J.M.; Deutch, A.Y. Organization of afferents to the orbitofrontal cortex in the rat. J. Comp. Neurol. 2018, 526, 1498–1526. [Google Scholar] [CrossRef] [PubMed]
- Gourley, S.L.; Olevska, A.; Zimmermann, K.S.; Ressler, K.J.; Dileone, R.J.; Taylor, J.R. The orbitofrontal cortex regulates outcome-based decision-making via the lateral striatum. Eur. J. Neurosci. 2013, 38, 2382–2388. [Google Scholar] [CrossRef]
- Puighermanal, E.; Castell, L.; Esteve-Codina, A.; Melser, S.; Kaganovsky, K.; Zussy, C.; Boubaker-Vitre, J.; Gut, M.; Rialle, S.; Kellendonk, C.; et al. Functional and molecular heterogeneity of D2R neurons along dorsal ventral axis in the striatum. Nat. Commun. 2020, 11, 1957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Cheng, Y.; Xie, X.; Woodson, K.; Bonifacio, J.; Disney, E.; Barbee, B.; Wang, X.; Zaidi, M.; Wang, J. Whole-Brain Mapping of Direct Inputs to Dopamine D1 and D2 Receptor-Expressing Medium Spiny Neurons in the Posterior Dorsomedial Striatum. eNeuro 2021, 8. [Google Scholar] [CrossRef]
- Freeman, W.M.; Nader, M.A.; Nader, S.H.; Robertson, D.J.; Gioia, L.; Mitchell, S.M.; Daunais, J.B.; Porrino, L.J.; Friedman, D.P.; Vrana, K.E. Chronic cocaine-mediated changes in non-human primate nucleus accumbens gene expression. J. Neurochem. 2001, 77, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Amaya, K.A.; Smith, K.S. Neurobiology of habit formation. Curr. Opin. Behav. Sci. 2018, 20, 145–152. [Google Scholar] [CrossRef]
- Jedynak, J.P.; Uslaner, J.M.; Esteban, J.A.; Robinson, T.E. Methamphetamine-induced structural plasticity in the dorsal striatum. Eur. J. Neurosci. 2007, 25, 847–853. [Google Scholar] [CrossRef] [Green Version]
- Montalban, E.; Al-Massadi, O.; Sancho-Balsells, A.; Brito, V.; de Pins, B.; Alberch, J.; Ginés, S.; Girault, J.-A.; Giralt, A. Pyk2 in the amygdala modulates chronic stress sequelae via PSD-95-related micro-structural changes. Transl. Psychiatry 2019, 9, 3. [Google Scholar] [CrossRef] [Green Version]
- Wemm, S.E.; Sinha, R. Drug-induced stress responses and addiction risk and relapse. Neurobiol. Stress 2019, 10, 100148. [Google Scholar] [CrossRef]
- DePoy, L.M.; Zimmermann, K.S.; Marvar, P.J.; Gourley, S.L. Induction and Blockade of Adolescent Cocaine-Induced Habits. Biol. Psychiatry 2017, 81, 595–605. [Google Scholar] [CrossRef] [Green Version]
- Spencer, S.; Garcia-Keller, C.; Roberts-Wolfe, D.; Heinsbroek, J.A.; Mulvaney, M.; Sorrell, A.; Kalivas, P.W. Cocaine Use Reverses Striatal Plasticity Produced During Cocaine Seeking. Biol. Psychiatry 2017, 81, 616–624. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Keller, C.; Smiley, C.; Monforton, C.; Melton, S.; Kalivas, P.W.; Gass, J. N-Acetylcysteine treatment during acute stress prevents stress-induced augmentation of addictive drug use and relapse. Addict. Biol. 2020, 25, e12798. [Google Scholar] [CrossRef]
- Garcia-Keller, C.; Neuhofer, D.; Bobadilla, A.-C.; Spencer, S.; Chioma, V.C.; Monforton, C.; Kalivas, P.W. Extracellular Matrix Signaling Through β3 Integrin Mediates Cocaine Cue–Induced Transient Synaptic Plasticity and Relapse. Biol. Psychiatry 2019, 86, 377–387. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gourley, S.L.; Srikanth, K.D.; Woon, E.P.; Gil-Henn, H. Pyk2 Stabilizes Striatal Medium Spiny Neuron Structure and Striatal-Dependent Action. Cells 2021, 10, 3442. https://doi.org/10.3390/cells10123442
Gourley SL, Srikanth KD, Woon EP, Gil-Henn H. Pyk2 Stabilizes Striatal Medium Spiny Neuron Structure and Striatal-Dependent Action. Cells. 2021; 10(12):3442. https://doi.org/10.3390/cells10123442
Chicago/Turabian StyleGourley, Shannon L., Kolluru D. Srikanth, Ellen P. Woon, and Hava Gil-Henn. 2021. "Pyk2 Stabilizes Striatal Medium Spiny Neuron Structure and Striatal-Dependent Action" Cells 10, no. 12: 3442. https://doi.org/10.3390/cells10123442