
High-Throughput Functional Analysis of CFTR and Other Apically Localized Proteins in iPSC-Derived Human Intestinal Organoids

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. ESC (Embryonic Stem Cell) and iPSC-Derived Human Intestinal Organoids (HIO)
2.2. MDCK and MDCK (Tagged αβγENaC) Cells
2.3. Swelling Assay
2.4. Opened Organoid Cultures
2.5. Apical Chloride Conductance (ACC) Assay for CFTR Function
2.6. Apical Sodium Conductance (ASC) Assay for ENaC Function
2.7. Apical Amino Acid Conductance (AAC) Assay for SLC6A14 Function
2.8. FLIPR Analysis and Heatmap Generation
2.9. Real-Time Quantitative PCR
2.10. Immunofluorescence
2.11. Western Blotting
2.12. Statical Analysis
3. Results
3.1. 3D CF Human Intestinal Organoids (HIO) Exhibit Defective Fluid Secretion, Which Can Be Restored through the Use of CFTR Modulators or Gene Editing
3.2. Opened HIOs Enable Direct Assessment of Apical Wt-CFTR Channel Function in a High-Throughput Format
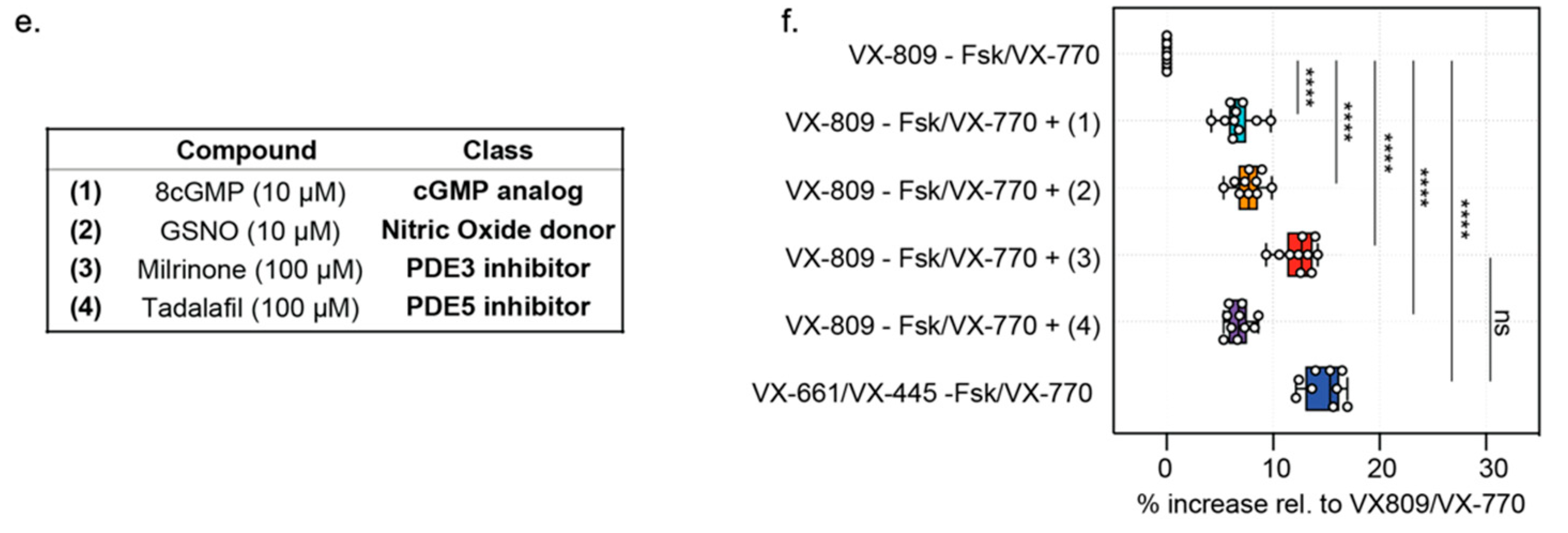
3.3. Opened CF Organoids Can Model Pharmacological Rescue of F508del-CFTR with CF Modulators
3.4. Measurement of ENaC Specific Activity in MDCK Overexpression Cells
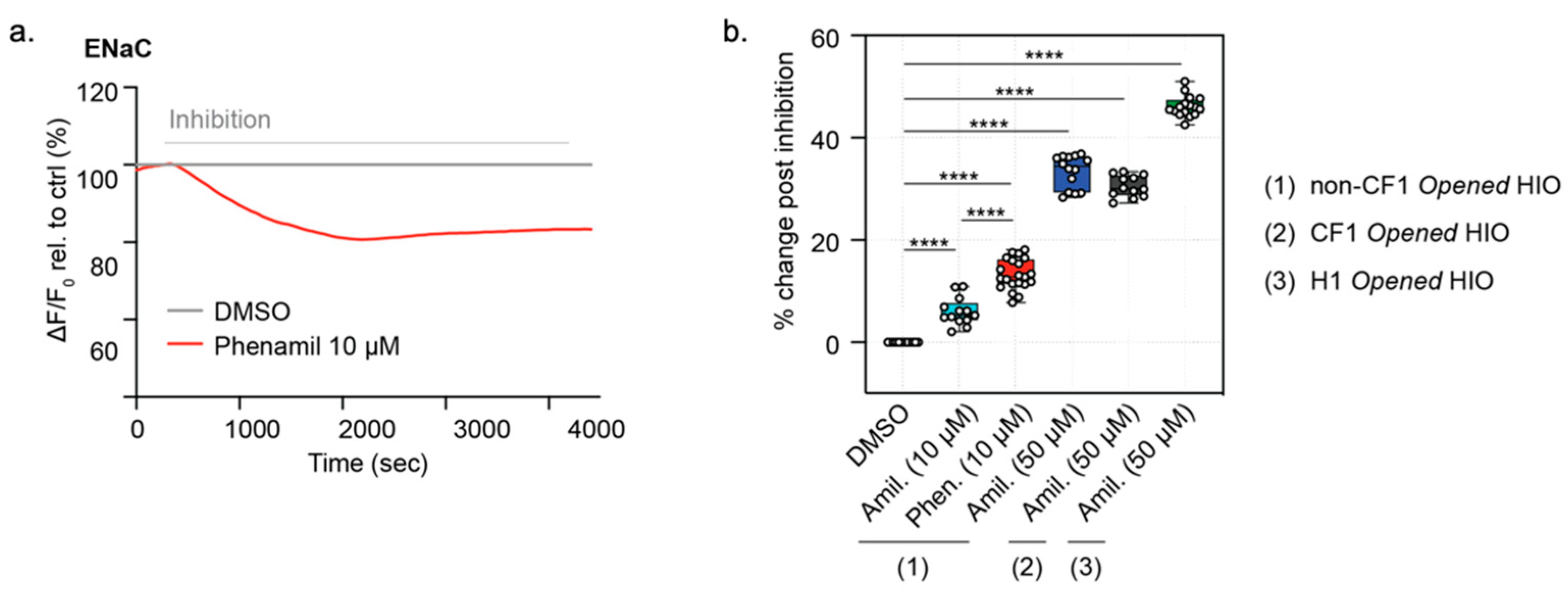
3.5. Function of ENaC Activity Can Be Measured in Both Opened CF HIOs and Non-CF HIOs and HCOs
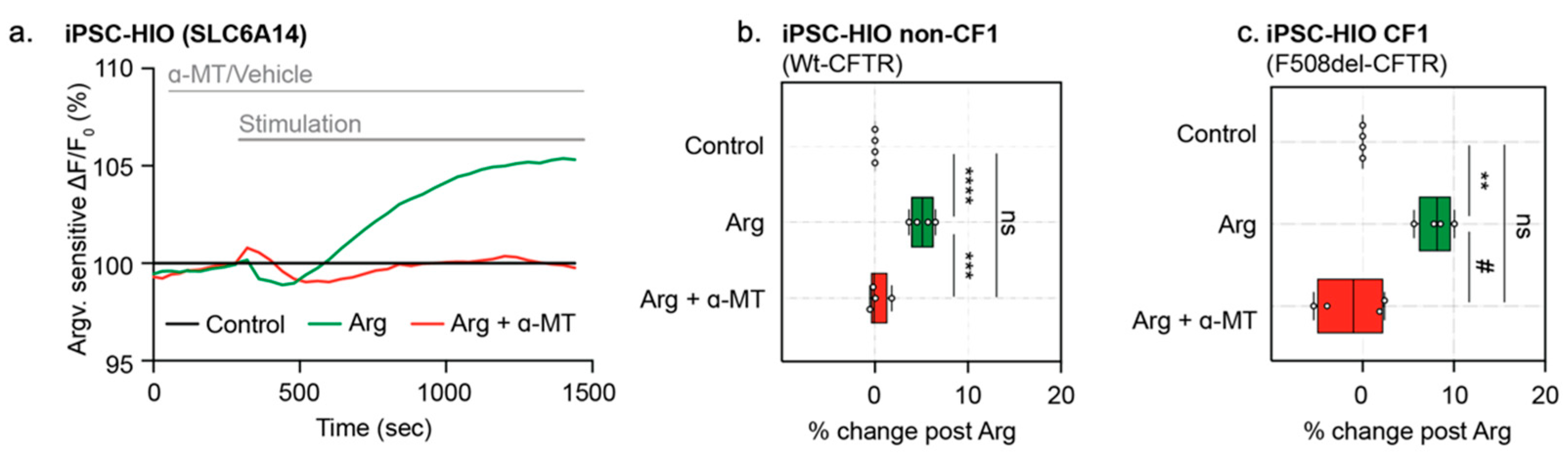
3.6. Specific Activity of Sodium and Chloride-Dependent Amino Acid Transporter, SLC6A14, Can Be Measured in Opened CF and Non-CF HIOs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sato, T.; Vries, R.G.; Snippert, H.J.; Van De Wetering, M.; Barker, N.; Stange, D.E.; Van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J.; et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, J.F.; Wiegerinck, C.L.; De Jonge, H.R.; Bronsveld, I.; Janssens, H.M.; De Winter-de Groot, K.M.; Brandsma, A.M.; de Jong, N.W.; Bijvelds, M.J.; Scholte, B.J.; et al. A functional CFTR assay using primary cystic fibrosis intestinal organoids. Nat. Med. 2013, 19, 939–945. [Google Scholar] [CrossRef]
- Watson, C.L.; Mahe, M.; Múnera, J.; Howell, J.C.; Sundaram, N.; Poling, H.M.; Schweitzer, I.J.; Vallance, J.; Mayhew, C.; Sun, Y.; et al. An in vivo model of human small intestine using pluripotent stem cells. Nat. Med. 2014, 20, 1310–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mithal, A.; Capilla, A.; Heinze, D.; Berical, A.; Villacorta-Martin, C.; Vedaie, M.; Jacob, A.; Abo, K.; Szymaniak, A.; Peasley, M.; et al. Generation of mesenchyme free intestinal organoids from human induced pluripotent stem cells. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Merkert, S.; Schubert, M.; Olmer, R.; Engels, L.; Radetzki, S.; Veltman, M.; Scholte, B.J.; Zöllner, J.; Pedemonte, N.; Galietta, L.J.; et al. High-Throughput Screening for Modulators of CFTR Activity Based on Genetically Engineered Cystic Fibrosis Disease-Specific iPSCs. Stem Cell Rep. 2019, 12, 1389–1403. [Google Scholar] [CrossRef] [Green Version]
- Dekkers, J.F.; Berkers, G.; Kruisselbrink, E.; Vonk, A.; de Jonge, H.R.; Janssens, H.M.; Bronsveld, I.; van de Graaf, E.A.; Nieuwenhuis, E.E.S.; Houwen, R.H.J.; et al. Characterizing responses to CFTR-modulating drugs using rectal organoids derived from subjects with cystic fibrosis. Sci. Transl. Med. 2016, 8, 344ra84. [Google Scholar] [CrossRef] [PubMed]
- Zomer-van Ommen, D.D.; de Poel, E.; Kruisselbrink, E.; Oppelaar, H.; Vonk, A.M.; Janssens, H.M. Comparison of ex vivo and in vitro intestinal cystic fibrosis models to measure CFTR-dependent ion channel activity. J. Cyst. Fibros. 2018, 17, 316–324. [Google Scholar] [CrossRef]
- Mall, M.; Bleich, M.; Kuehr, J.; Brandis, M.; Greger, R.; Kunzelmann, K. CFTR-mediated inhibition of epithelial Na+ conductance in human colon is defective in cystic fibrosis. Am. J. Physiol. Content 1999, 277, G709–G716. [Google Scholar] [CrossRef] [PubMed]
- Moore, P.J.; Tarran, R. The epithelial sodium channel (ENaC) as a therapeutic target for cystic fibrosis lung disease. Expert Opin. Ther. Targets 2018, 22, 687–701. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, S.; Xia, S.; Wu, Y.-S.; Di Paola, M.; Kissoon, R.; Luk, C.; Lin, F.; Du, K.; Rommens, J.; E Bear, C. SLC6A14, an amino acid transporter, modifies the primary CF defect in fluid secretion. eLife 2018, 7, e37963. [Google Scholar] [CrossRef]
- Mall, M.A. ENaC inhibition in cystic fibrosis: Potential role in the new era of CFTR modulator therapies. Eur. Respir. J. 2020, 56, 2000946. [Google Scholar] [CrossRef] [PubMed]
- Eckford, P.D.; McCormack, J.; Munsie, L.; He, G.; Stanojevic, S.; Pereira, S.L.; Ho, K.; Avolio, J.; Bartlett, C.; Yang, J.Y.; et al. The CF Canada-Sick Kids Program in individual CF therapy: A resource for the advancement of personalized medicine in CF. J. Cyst. Fibros. 2019, 18, 35–43. [Google Scholar] [CrossRef] [Green Version]
- McCracken, K.W.; Howell, J.C.; Wells, J.M.; Spence, J.R. Generating human intestinal tissue from pluripotent stem cells in vitro. Nat. Protoc. 2011, 6, 1920–1928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spence, J.R.; Mayhew, C.N.; Rankin, S.A.; Kuhar, M.F.; Vallance, J.E.; Tolle, K. Directed differentiation of human pluripotent stem cells into intestinal tissue in vitro. Nature 2011, 470, 105–109. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Pribanic, S.; Debonneville, A.; Jiang, C.; Rotin, D. The PY Motif of ENaC, Mutated in Liddle Syndrome, Regulates Channel Internalization, Sorting and Mobilization from Subapical Pool. Traffic 2007, 8, 1246–1264. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, S.; Bozoky, Z.; Di Paola, M.; Xia, S.; Li, C.; Wong, A.; Wellhauser, L.; Molinski, S.; Ip, W.; Ouyang, H.; et al. Phenotypic profiling of CFTR modulators in patient-derived respiratory epithelia. NPJ Genom. Med. 2017, 2, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.X.; Gatfield, K.; Ward, E.; Downie, D.; Sneddon, H.F.; Walsh, S.; Powell, A.; Laine, D.; Carr, M.; Trezise, D. Validation and Optimization of Novel High-Throughput Assays for Human Epithelial Sodium Channels. J. Biomol. Screen. 2015, 20, 242–253. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.-S.; Jiang, J.; Ahmadi, S.; Lew, A.; Laselva, O.; Xia, S.; Bartlett, C.; Ip, W.; Wellhauser, L.; Ouyang, H.; et al. ORKAMBI-Mediated Rescue of Mucociliary Clearance in Cystic Fibrosis Primary Respiratory Cultures Is Enhanced by Arginine Uptake, Arginase Inhibition, and Promotion of Nitric Oxide Signaling to the Cystic Fibrosis Transmembrane Conductance Regulator Channel. Mol. Pharmacol. 2019, 96, 515–525. [Google Scholar] [CrossRef]
- Di Paola, M.; Park, A.J.; Ahmadi, S.; Roach, E.J.; Wu, Y.S.; Struder-Kypke, M. SLC6A14 Is a Genetic Modifier of Cystic Fibrosis That Regulates Pseudomonas aeruginosa Attachment to Human Bronchial Epithelial Cells. mBio 2017, 8, e02073-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karunakaran, S.; Umapathy, N.S.; Thangaraju, M.; Hatanaka, T.; Itagaki, S.; Munn, D.; Prasad, P.D.; Ganapathy, V. Interaction of tryptophan derivatives with SLC6A14 (ATB0,+) reveals the potential of the transporter as a drug target for cancer chemotherapy. Biochem. J. 2008, 414, 343–355. [Google Scholar] [CrossRef]
- Laselva, O.; Stone, T.A.; Bear, C.E.; Deber, C.M. Anti-Infectives Restore ORKAMBI((R)) Rescue of F508del-CFTR Function in Human Bronchial Epithelial Cells Infected with Clinical Strains of P. aeruginosa. Biomolecules 2020, 10, 334. [Google Scholar] [CrossRef] [Green Version]
- Molinski, S.V.; Ahmadi, S.; Ip, W.; Ouyang, H.; Villella, A.; Miller, J.P. Orkambi(R) and amplifier co-therapy improves function from a rare CFTR mutation in gene-edited cells and patient tissue. EMBO Mol. Med. 2017, 9, 1224–1243. [Google Scholar] [CrossRef] [PubMed]
- Kopeikin, Z.; Yuksek, Z.; Yang, H.-Y.; Bompadre, S. Combined effects of VX-770 and VX-809 on several functional abnormalities of F508del-CFTR channels. J. Cyst. Fibros. 2014, 13, 508–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Middleton, P.G.; Mall, M.A.; Dřevínek, P.; Lands, L.C.; McKone, E.F.; Polineni, D.; Ramsey, B.W.; Taylor-Cousar, J.L.; Tullis, E.; Vermeulen, F.; et al. Elexacaftor–Tezacaftor–Ivacaftor for Cystic Fibrosis with a Single Phe508del Allele. N. Engl. J. Med. 2019, 381, 1809–1819. [Google Scholar] [CrossRef]
- McClure, M.L.; Barnes, S.; Brodsky, J.L.; Sorscher, E.J. Trafficking and function of the cystic fibrosis transmembrane conductance regulator: A complex network of posttranslational modifications. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 311, L719–L733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pankow, S.; Bamberger, C.; Yates, J.R. A posttranslational modification code for CFTR maturation is altered in cystic fibrosis. Sci. Signal. 2019, 12, eaan7984. [Google Scholar] [CrossRef]
- Hobbs, C.A.; Da Tan, C.; Tarran, R. Does epithelial sodium channel hyperactivity contribute to cystic fibrosis lung disease? J. Physiol. 2013, 591, 4377–4387. [Google Scholar] [CrossRef]
- Mall, M.A.; Galietta, L.J. Targeting ion channels in cystic fibrosis. J. Cyst. Fibros. 2015, 14, 561–570. [Google Scholar] [CrossRef] [Green Version]
- Scott, D.W.; Walker, M.P.; Sesma, J.; Wu, B.; Stuhlmiller, T.J.; Sabater, J.R.; Abraham, W.M.; Crowder, T.M.; Christensen, D.J.; Tarran, R. SPX-101 Is a Novel Epithelial Sodium Channel–targeted Therapeutic for Cystic Fibrosis That Restores Mucus Transport. Am. J. Respir. Crit. Care Med. 2017, 196, 734–744. [Google Scholar] [CrossRef] [PubMed]
- Bhalla, V.; Hallows, K.R. Mechanisms of ENaC Regulation and Clinical Implications. J. Am. Soc. Nephrol. 2008, 19, 1845–1854. [Google Scholar] [CrossRef] [Green Version]
- Repishti, M.; Hogan, D.L.; Pratha, V.; Davydova, L.; Donowitz, M.; Tse, C.M. Human duodenal mucosal brush border Na(+)/H(+) exchangers NHE2 and NHE3 alter net bicarbonate movement. Am. J. Physiol. Gastrointest Liver Physiol. 2001, 281, G159–G163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirsh, A.J.; Sabater, J.R.; Zamurs, A.; Smith, R.T.; Paradiso, A.M.; Hopkins, S. Evaluation of second generation amiloride analogs as therapy for cystic fibrosis lung disease. J. Pharmacol. Exp. Ther. 2004, 311, 929–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ESC and iPSC Lines | CFTR Genotype | Description and Source |
|---|---|---|
| CF 1 | F508del/F508del | iPSC / CFIT Program 1 |
| Non-CF 1 | Wt/Wt | iPSC / CFIT Program 1 |
| Non-CF 2 | Wt/Wt | iPSC / CFIT Program 1 |
| H1 | Wt/Wt | ESC / Cincinnati Children’s Hospital 2 |
| Primer Sequence | |
|---|---|
| CFTR | Fwd: 5′-CGGAGTGATAACACAGAAAGT-3′ |
| Rev: 5′-CAGGAAACTGCTCTATTACAGAC-3′ | |
| αENaC | Fwd: 5′-TTGACGTCTCCAACTCACCG-3′ |
| Rev: 5′-GGCAGAGGAGGACAAAGGTC-3′ | |
| SLC6A14 | Fwd: 5′-GCTTGCTGGTTTGTCATCACTCC-3′ |
| Rev: 5′-TACACCAGCCAAGAGCAACTCC-3′ | |
| EpCAM | Fwd: 5′-AACACAAGACGACGTGGACA-3′ |
| Rev: 5′-GCTCTCCGTTCACTCTCAGG-3′ | |
| Villin | Fwd: 5′-CTGTGATGTCCAGCGACTGT-3′ |
| Rev: 5′-CTCTCTGGCCCATTCCACTG-3′ | |
| MUC2 | Fwd: 5′-GCTGCTATGTCGAGGACACC-3′ |
| Rev: 5′-GGGAGGAGTTGGTACACACG-3′ | |
| TBP | Fwd: 5′-CAAACCCAGAATTGTTCTCCTT-3′ |
| Rev: 5′-ATGTGGTCTTCCTGAATCCCT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xia, S.; Bozóky, Z.; Di Paola, M.; Laselva, O.; Ahmadi, S.; Jiang, J.X.; Pitstick, A.L.; Jiang, C.; Rotin, D.; Mayhew, C.N.; et al. High-Throughput Functional Analysis of CFTR and Other Apically Localized Proteins in iPSC-Derived Human Intestinal Organoids. Cells 2021, 10, 3419. https://doi.org/10.3390/cells10123419
Xia S, Bozóky Z, Di Paola M, Laselva O, Ahmadi S, Jiang JX, Pitstick AL, Jiang C, Rotin D, Mayhew CN, et al. High-Throughput Functional Analysis of CFTR and Other Apically Localized Proteins in iPSC-Derived Human Intestinal Organoids. Cells. 2021; 10(12):3419. https://doi.org/10.3390/cells10123419
Chicago/Turabian StyleXia, Sunny, Zoltán Bozóky, Michelle Di Paola, Onofrio Laselva, Saumel Ahmadi, Jia Xin Jiang, Amy L. Pitstick, Chong Jiang, Daniela Rotin, Christopher N. Mayhew, and et al. 2021. "High-Throughput Functional Analysis of CFTR and Other Apically Localized Proteins in iPSC-Derived Human Intestinal Organoids" Cells 10, no. 12: 3419. https://doi.org/10.3390/cells10123419