
Tomato Yellow Leaf Curl Virus (TYLCV) Promotes Plant Tolerance to Drought

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses, Insects and Plants
2.2. Drought Treatment
2.3. Analysis of Metabolites
2.4. Measurements of Quantitative Physiological Traits in iCORE Greenhouse
2.5. Protein Immunodetection
2.6. Drought Recovery Treatment
2.7. Statistical Analyzes
3. Results
3.1. TYLCV Infection of a TYLCV-Resistant Tomato Line Enhanced Plant Survival When Grown under Water Deficit
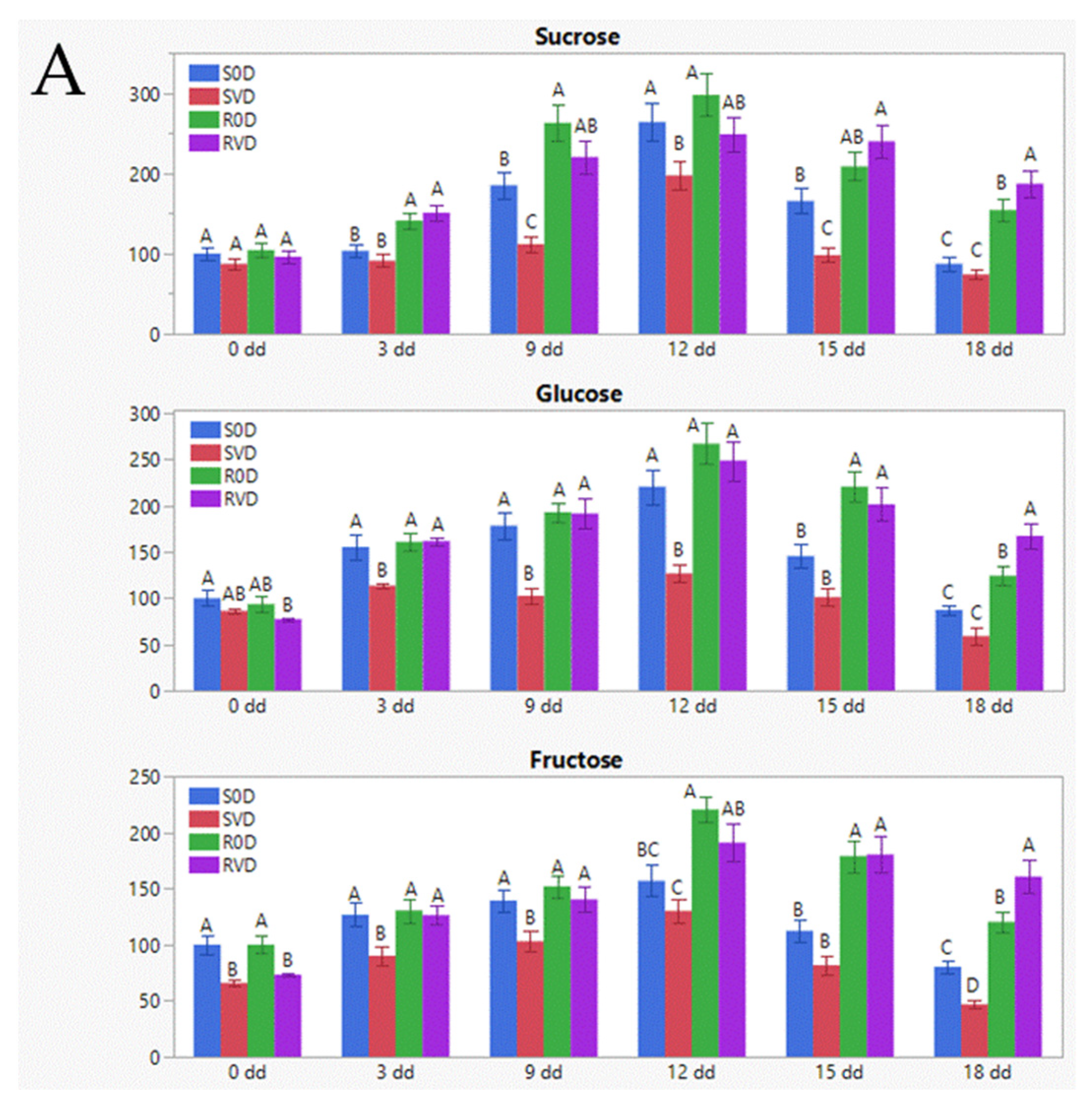
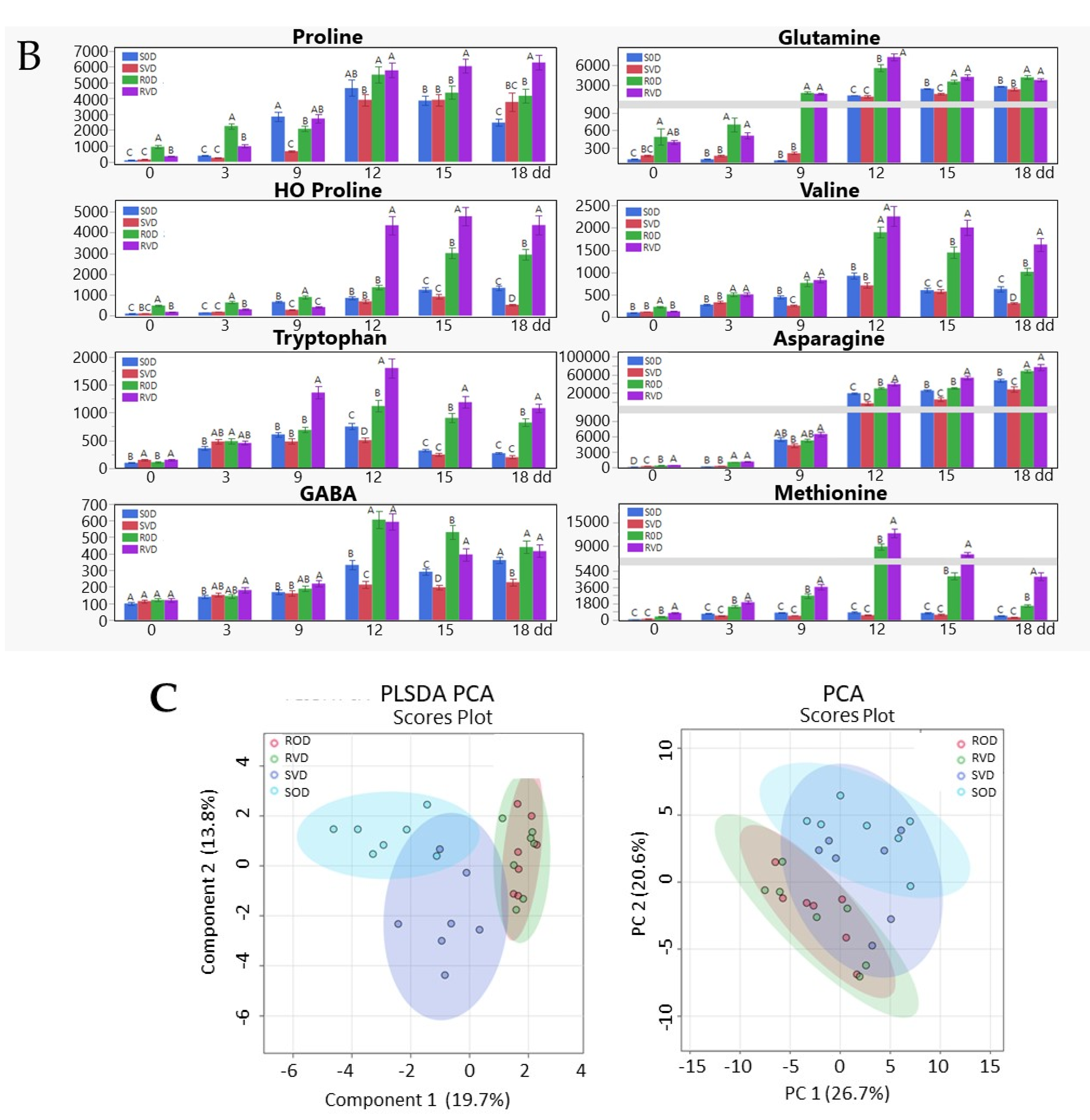
3.2. Osmo-Protectants (Sugars, Amino Acids) Accumulate in TYLCV-Infected Tomatoes Grown in Conditions of Water Deficit
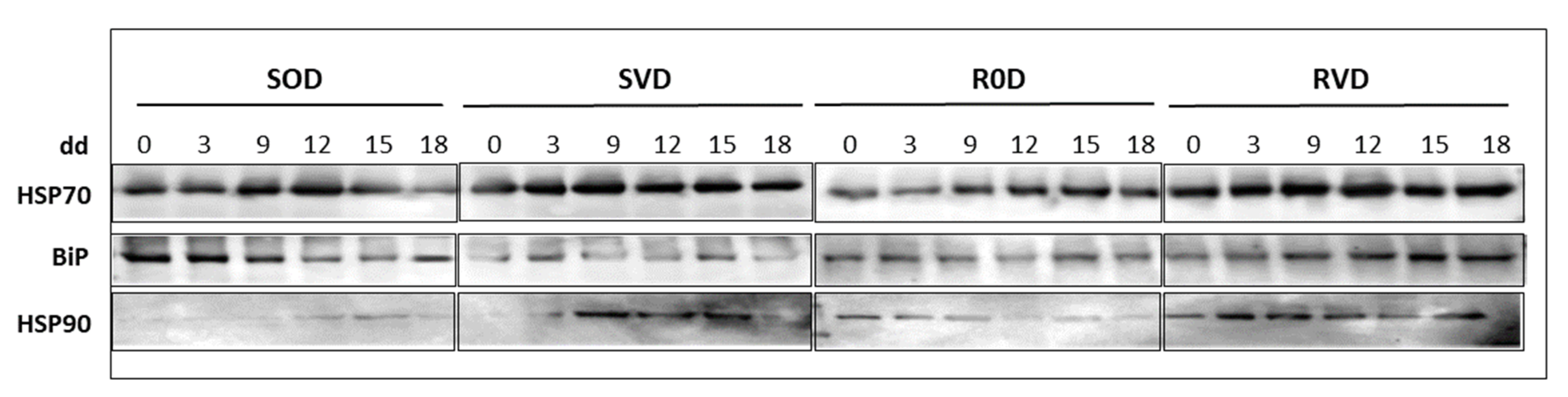
3.3. Enhanced Stabilization of HSPs Patterns in Leaves of Tomatoes Exposed to Prolonged Drought
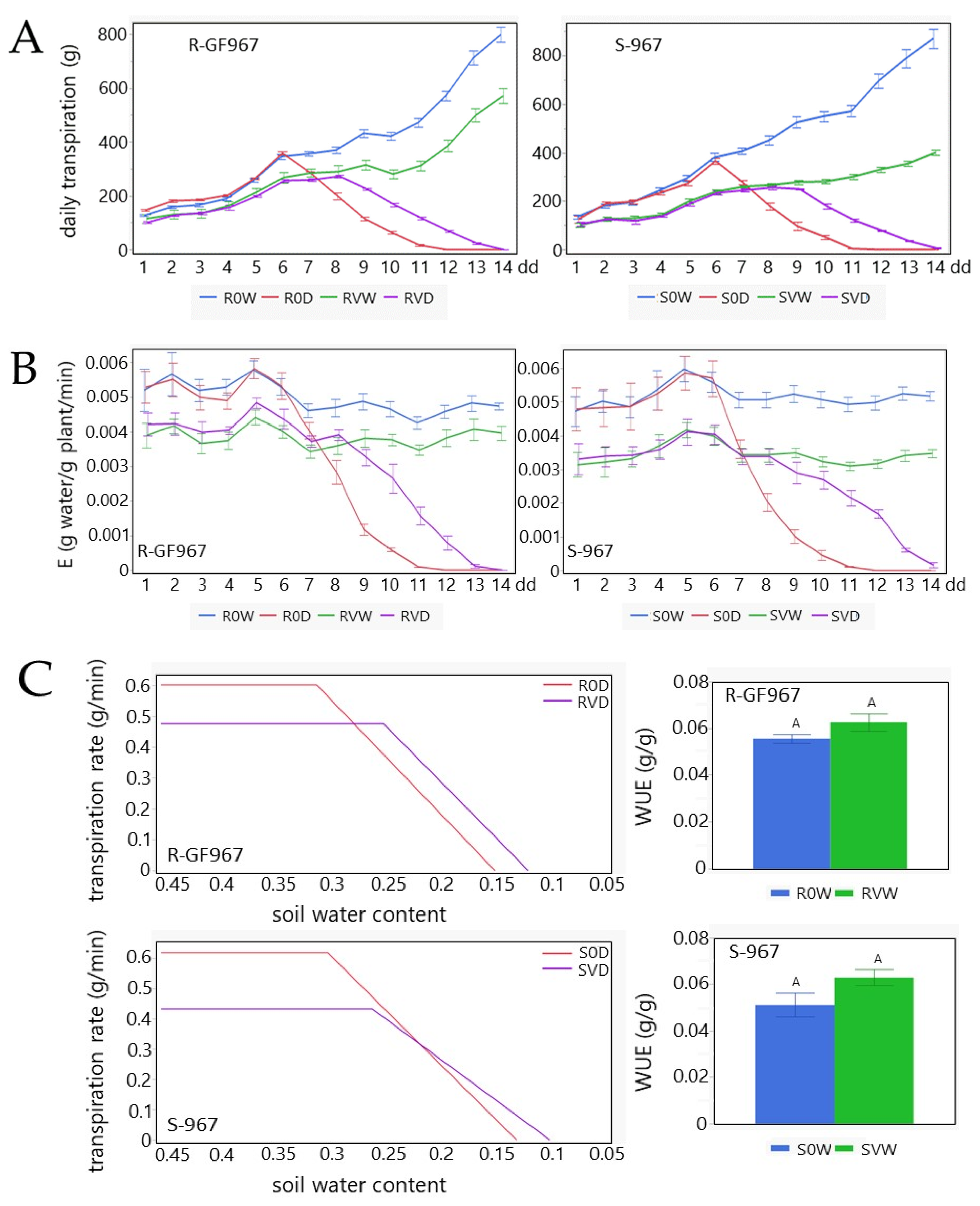
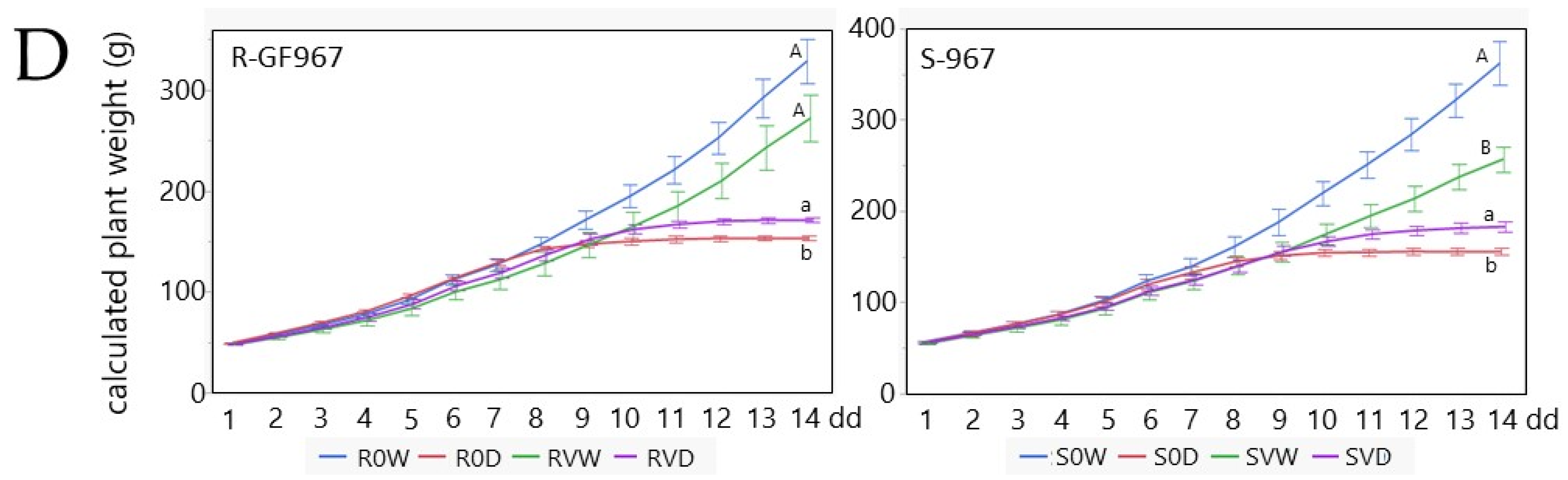
3.4. Transpiration, Whole Canopy Stomatal Conductance and Physiological Drought in TYLVC-Susceptible and Resistant Tomatoes Grown under the Water Deficit
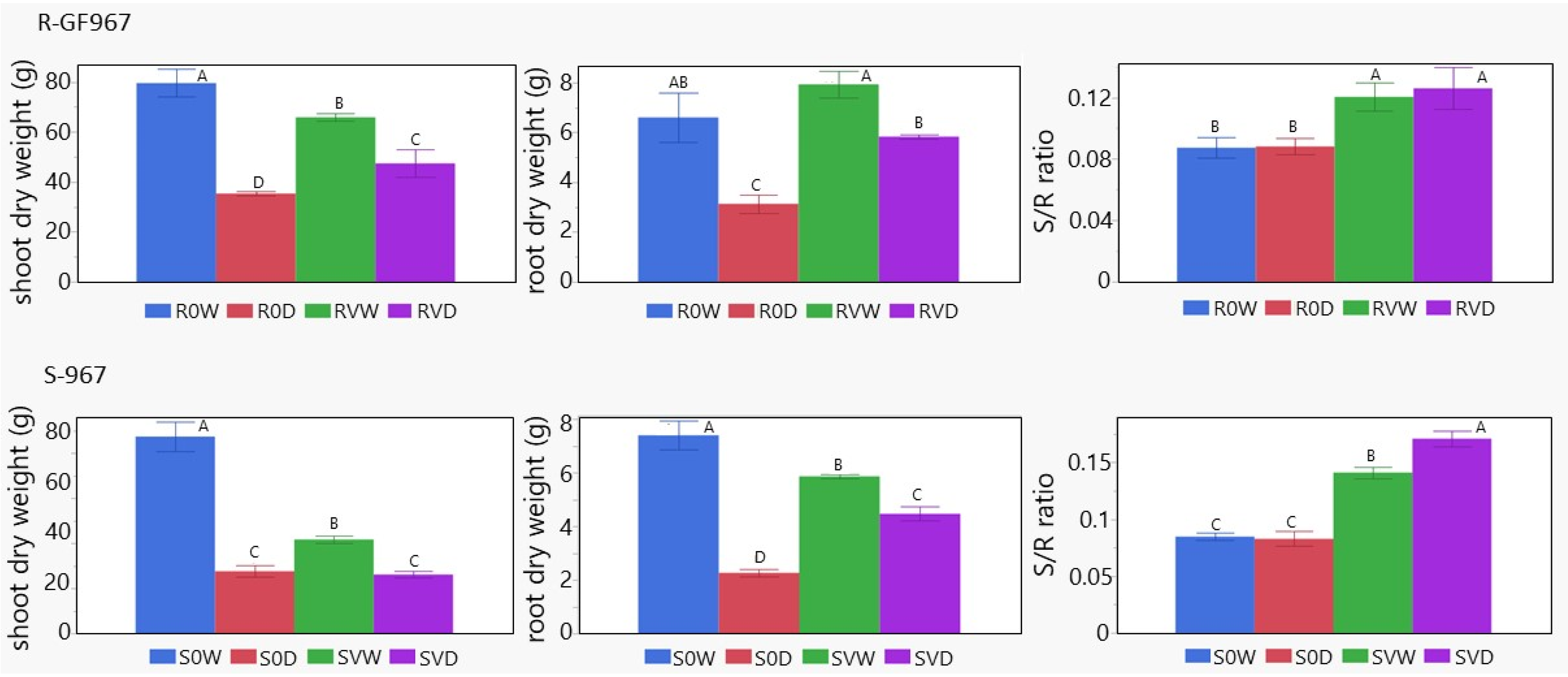
3.5. Shift of Biomass from Shoot to Root Caused by the Presence of TYLCV
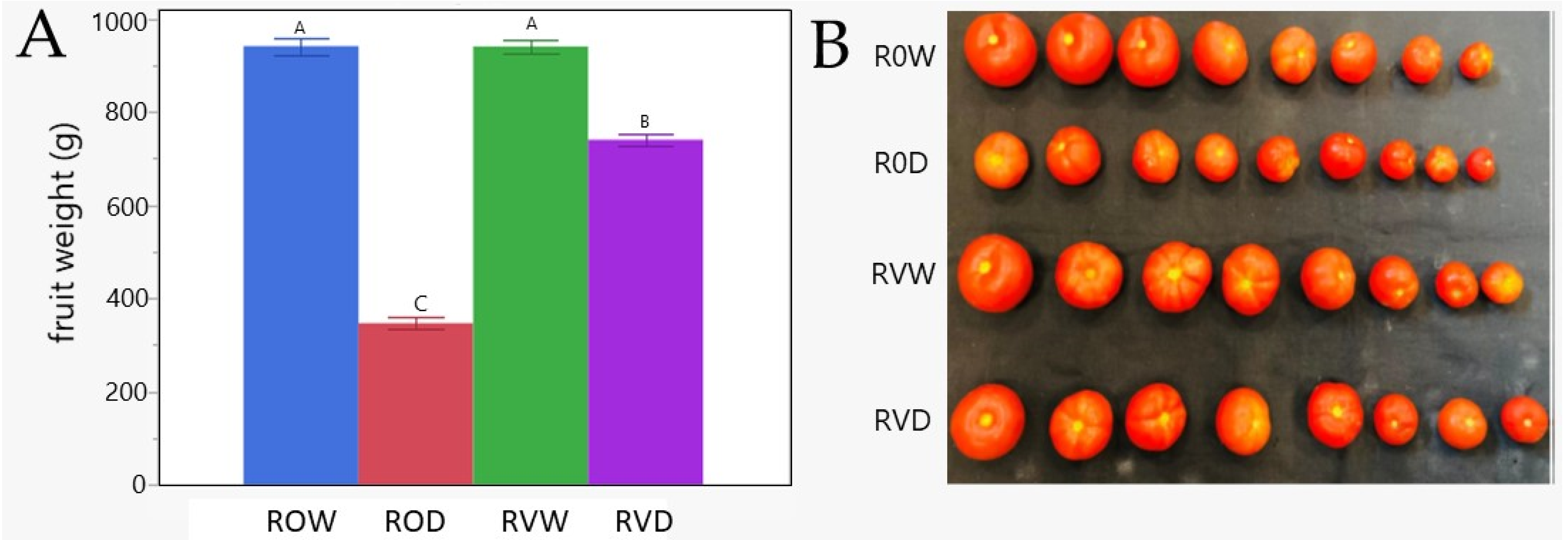
3.6. Infected TYLCV-Resistant Plants Subjected to Drought for 14 Days Were Able to Recover and Produce Fruits
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anfoka, G.; Moshe, A.; Fridman, L.; Amrani, L.; Rotem, O.; Kolot, M.; Zeidan, M.; Czosnek, H.; Gorovits, R. Tomato yellow leaf curl virus infection mitigates the heat stress response of plants grown at high temperatures. Sci. Rep. 2016, 6, 19715. [Google Scholar] [CrossRef] [Green Version]
- Czosnek, H. Tomato Yellow Leaf Curl Viruses (Geminiviridae). Encyclopedia of Virology, 4th ed.; Bamford, B., Zuckerman, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; p. 5031. [Google Scholar]
- Hanley-Bowdoin, L.; Settlage, S.B.; Robertson, D. Reprogramming plant gene expression: A prerequisite to geminivirus DNA replication. Mol. Plant Path. 2004, 5, 149–156. [Google Scholar] [CrossRef]
- Moshe, A.; Gorovits, R.; Liu, Y.; Czosnek, H. Tomato plant cell death induced by inhibition of HSP90 is alleviated by Tomato yellow leaf curl virus infection. Mol. Plant Path. 2016, 17, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Gorovits, R.; Sobol, I.; Altaleb, M.; Czosnek, H.; Anfoka, G. Taking advantage of a pathogen: Understanding how a virus alleviates plant stress response. Phytopath. Res. 2019, 1, 20. [Google Scholar] [CrossRef]
- Scharf, K.D.; Berberich, T.; Ebersberger, I.; Nover, L. The plant heat stress transcription factor (Hsf) family: Structure, function and evolution. Biochim. Biophys. Acta 2012, 1819, 104–119. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Y.; Xing, D.; Gao, C. Characterization of mitochondrial dynamics and subcellular localization of ROS reveal that HsfA2 alleviates oxidative damage caused by heat stress in Arabidopsis. J. Exp. Bot. 2009, 60, 2073–2091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, R.; Shteinberg, M.; Shkolnik, D.; Anfoka, G.; Czosnek, H.; Gorovits, R. Interplay between abiotic (drought) and biotic (virus) stresses in tomato plants. Plant Mol. Pathol. 2021, 22. in press. [Google Scholar]
- Anfoka, G.; Abhary, M.; Nakhla, M.K. Molecular identification of species of the Tomato yellow leaf curl virus complex in Jordan. J. Plant Pathol. 2005, 87, 65–70. [Google Scholar]
- Navot, N.; Pichersky, E.; Zeidan, M.; Zamir, D.; Czosnek, H. Tomato yellow leaf curl virus: A whitefly-transmitted geminivirus with a single genomic molecule. Virology 1991, 185, 151–161. [Google Scholar] [CrossRef]
- Qaryouti, M.M.; Hurani, O.M.; Mahadeen, A.Y. Susceptibility of Jordanian tomato landraces to Tomato yellow leaf curl virus. Plant Gen. Res. Newslett. 2003, 136, 31–33. [Google Scholar]
- Mejía, L.; Teni, R.E.; Vidavski, F.; Czosnek, H. Evaluation of tomato germplasm and selection of breeding lines for resistance to begomoviruses in Guatemala. Acta Horticul. 2005, 695, 251–256. [Google Scholar] [CrossRef]
- Vidavsky, F.; Czosnek, H. Tomato breeding lines immune and tolerant to tomato yellow leaf curl virus (TYLCV) issued from Lycopersicum hirsutum. Phytopatholy 1998, 88, 910–914. [Google Scholar] [CrossRef] [Green Version]
- Gorovits, R.; Akad, F.; Beery, H.; Vidavsky, F.; Mahadav, A.; Czosnek, H. Expression of stress-response proteins upon whitefly-mediated inoculation of Tomato yellow leaf curl virus (TYLCV) in susceptible and resistant tomato plants. Mol. Plant Microbe Interact. 2007, 20, 1376–1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorovits, R.; Moshe, A.; Kolot, M.; Sobol, I.; Czosnek, H. Progressive aggregation of Tomato yellow leaf curl virus coat protein in systemically infected tomato plants, susceptible and resistant to the virus. Virus Res. 2013, 171, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Lisec, J.; Schauer, N.; Kopka, J.; Willmitzer, L.; Fernie, A.R. Gas chromatography mass spectrometry-based metabolite profiling in plants. Nat. Prot. 2006, 1, 387–396. [Google Scholar] [CrossRef]
- Caldana, C.; Degenkolbe, T.; Cuadros-Inostroza, A.; Klie, S.; Sulpice, R.; Leisse, A.; Steinhauser, D.; Fernie, A.R.; Willmitzer, L.; Hannah, M.A. High-density kinetic of the metabolomic and transcriptomic response of Arabidopsis to eight environmental conditions. Plant J. 2011, 67, 869–884. [Google Scholar] [CrossRef]
- Cuadros-Inostroza, A.; Caldana, C.; Redestig, H.; Kusano, M.; Lisec, J.; Pena-Cortes, H.; Willmitzer, L.; Hannah, M.A. Target Search–A Bioconductor package for the efficient preprocessing of GC-MS metabolite profiling data. BMC Bioinform. 2009, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Halperin, O.; Gebremedhin, A.; Wallach, R.; Moshelion, M. High-throughput physiological phenotyping and screening system for the characterization of plant–environment interactions. Plant J. 2017, 89, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Chi, Y.-H.; Koo, S.S.; Oh, H.T.; Lee, E.S.; Park, J.H.; Phan, K.A.T.; Wi, D.; Bae, S.B.; Paeng, S.K.; Chae, H.B.; et al. The physiological functions of universal stress proteins and their molecular mechanism to protect plants from environmental stresses. Front. Plant Sci. 2019, 10, 750. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.-K. Abiotic cell signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Fàbregas, N.; Fernie, A.R. The metabolic response to drought. J. Exp. Bot. 2019, 70, 1077–1085. [Google Scholar] [CrossRef] [Green Version]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obata, T.; Witt, S.; Lisec, J.; Palacios-Rojas, N.; Florez-Sarasa, I.; Yousfi, S.; Araus, J.L.; Cairns, J.E.; Fernie, A.R. Metabolite profiles of maize leaves in drought, heat, and combined stress field trials reveal the relationship between metabolism and grain yield. Plant Physiol. 2015, 169, 2665–2683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urano, K.; Maruyama, K.; Ogata, Y.; Morishita, Y.; Takeda, M.; Sakurai, N.; Suzuki, H.; Saito, K.; Shibata, D.; Kobayashi, M. Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics. Plant J. 2009, 57, 1065–1078. [Google Scholar] [CrossRef] [PubMed]
- Batista-Silva, W.; Heinemann, B.; Rugen, N.; Nunes-Nesi, A.; Araújo, W.L.; Braun, H.P. The role of amino acid metabolism during abiotic stress release. Plant Cell Environ. 2019, 42, 1630–1644. [Google Scholar] [CrossRef] [Green Version]
- Swindell, W.R.; Huebner, M.; Weber, A.P. Transcriptional profiling of Arabidopsis heat shock proteins and transcription factors reveals extensive overlap between heat and non-heat stress response pathways. BMC Genom. 2007, 8, 125. [Google Scholar] [CrossRef] [Green Version]
- Malmstrom, C.M.; Melcher, U.; Bosque-Perez, N.A. The expanding field of plant virus ecology: Historical foundations, knowledge gaps, and research directions. Virus Res. 2011, 159, 84–94. [Google Scholar] [CrossRef]
- Roossinck, M.J. Plant virus metagenomics: Biodiversity and ecology. Ann. Rev. Gen. 2012, 46, 359–369. [Google Scholar] [CrossRef]
- Xu, P.; Chen, F.; Manna, J.P.; Feldman, T.; Sumner, L.W.; Roossinck, M.J. Virus infection improves drought tolerance. New Phytol. 2008, 180, 911–921. [Google Scholar] [CrossRef]
- Westwood, J.H.; McCann, L.; Naish, M.; Dixon, H.; Murphy, A.M.; Stancombe, M.A.; Bennett, M.H.; Powell, G.; Webb, A.A.; Carr, J.P. A viral RNA silencing suppressor interferes with abscisic acid-mediated signalling and induces drought tolerance in Arabidopsis thaliana. Mol. Plant Pathol. 2013, 14, 158–170. [Google Scholar] [CrossRef]
- van Munster, M.; Yvon, M.; Vile, D.; Dader, B.; Fereres, A.; Blanc, S. Water deficit enhances the transmission of plant viruses by insect vectors. PLoS ONE 2017, 12, e0174398. [Google Scholar] [CrossRef]
- González, R.; Butković, A.; Escaray, F.J.; Martínez-Latorre, J.; Melero, Í.; Pérez-Parets, E.; Gómez-Cadenas, A.; Carrasco, P.; Elena, S.F. Plant virus evolution under strong drought conditions results in a transition from parasitism to mutualism. Proc. Natl. Acad. Sci. USA 2021, 118, e2020990118. [Google Scholar] [CrossRef]
- Aguilar, E.; Cutrona, C.; Del Toro, F.J.; Vallarino, J.G.; Osorio, S.; Perez-Bueno, M.L.; Baron, M.; Ching, B.N.; Tenllado, F. Virulence determines beneficial trade-offs in the response of virus-infected plants to drought via induction of salicylic acid. Plant Cell Environ. 2017, 40, 2909–2930. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Calvino, L.; Osorio, S.; Hernandez, M.L.; Hamada, I.B.; del Toro, F.J.; Donaire, L.; Yu, A.; Bustos, R.; Fernie, A.R.; Martínez-Rivas, J.M.; et al. Virus induced alterations in primary metabolism modulate susceptibility to tobacco rattle virus in Arabidopsis. Plant Physiol. 2014, 166, 1821–1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shabala, S.; Baekgaard, L.; Shabala, L.; Fuglsang, A.; Babourina, O.; Palmgren, M.G.; Cuin, T.A.; Rengel, Z.; Nemchinov, L.G. Plasma membrane Ca2+ transporters mediate virus-induced acquired resistance to oxidative stress: Calcium efflux and oxidative stress tolerance. Plant Cell Environ. 2011, 34, 406–417. [Google Scholar] [CrossRef] [PubMed]
- Woodrow, P.; Ciarmiello, L.F.; Annunziata, M.G.; Pacifico, S.; Iannuzzi, F.; Mirto, A.; D’Amelia, L.; Dell’Aversana, E.; Piccolella, S.; Fuggi, A.; et al. Durum wheat seedling responses to simultaneous high light and salinity involve a fine reconfiguration of amino acids and carbohydrate metabolism. Physiol. Plantar. 2017, 159, 290–312. [Google Scholar] [CrossRef]
- Gorovits, R.; Czosnek, H. Expression of stress gene networks in tomato lines susceptible and resistant to Tomato yellow leaf curl virus in response to abiotic stresses. Plant Physiol. Biochem. 2008, 46, 482–492. [Google Scholar] [CrossRef]
- Wilson, J.B. A review of evidence on the control of shoot: Root ratio, in relation to models. Ann. Bot. 1988, 61, 433–449. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shteinberg, M.; Mishra, R.; Anfoka, G.; Altaleb, M.; Brotman, Y.; Moshelion, M.; Gorovits, R.; Czosnek, H. Tomato Yellow Leaf Curl Virus (TYLCV) Promotes Plant Tolerance to Drought. Cells 2021, 10, 2875. https://doi.org/10.3390/cells10112875
Shteinberg M, Mishra R, Anfoka G, Altaleb M, Brotman Y, Moshelion M, Gorovits R, Czosnek H. Tomato Yellow Leaf Curl Virus (TYLCV) Promotes Plant Tolerance to Drought. Cells. 2021; 10(11):2875. https://doi.org/10.3390/cells10112875
Chicago/Turabian StyleShteinberg, Moshik, Ritesh Mishra, Ghandi Anfoka, Miassar Altaleb, Yariv Brotman, Menachem Moshelion, Rena Gorovits, and Henryk Czosnek. 2021. "Tomato Yellow Leaf Curl Virus (TYLCV) Promotes Plant Tolerance to Drought" Cells 10, no. 11: 2875. https://doi.org/10.3390/cells10112875