
Paracrine Regulation of Alveolar Epithelial Damage and Repair Responses by Human Lung-Resident Mesenchymal Stromal Cells

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. LMSC Isolation and Culture
2.3. Primary Human Lung Cell Isolation and Epithelial Cell Culture
2.4. Culture of Human Lung Epithelial Cell Lines
2.5. Treatment of Epithelial Cell Lines
2.6. Cell Viability by Annexin V/PI Staining
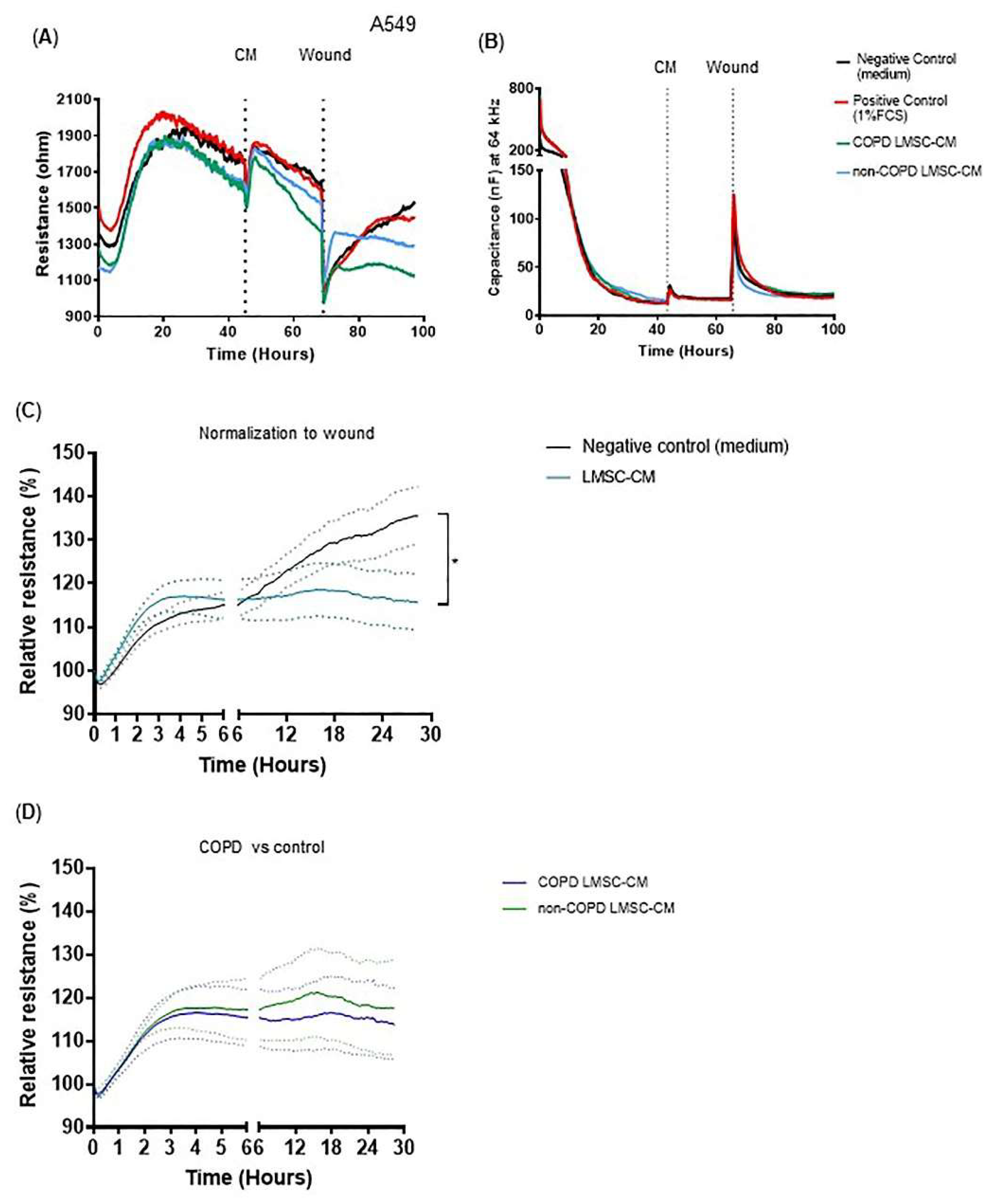
2.7. Electrical Resistance Measurements and Electric Field Injury
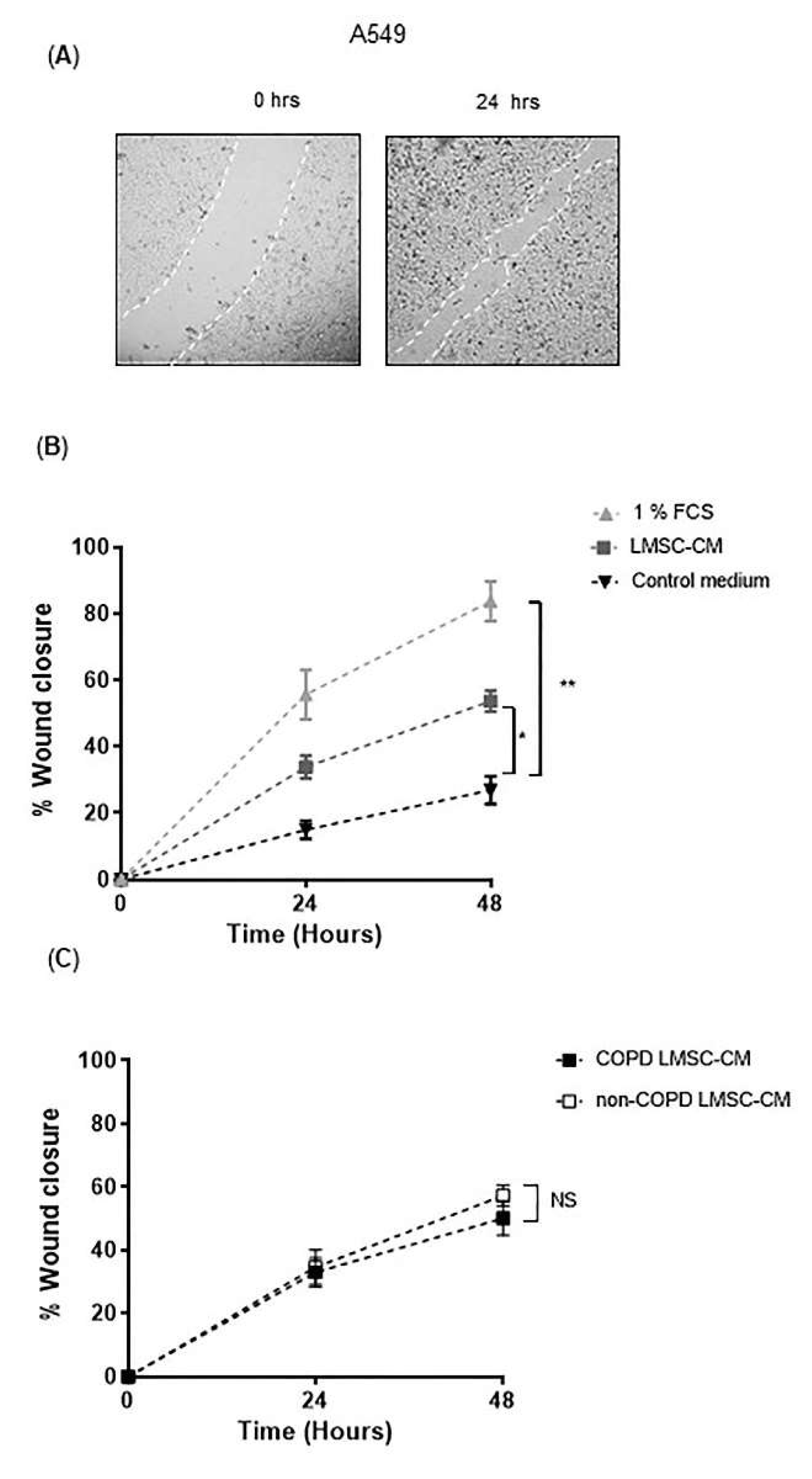
2.8. Scratch Wounding
2.9. qPCR
2.10. MRC-5 and LMSC Culture for Organoids
2.11. Organoid Culture of NCI-H441 and Primary Distal Lung Cells
2.12. Quantification of Organoid Size and Number
2.13. Staining for Type II Pneumocyte Markers SPC and HTII-280
2.14. Statistics
3. Results
3.1. Conditioned-Medium from LMSCs Protects from Oxidative Stress-Induced Cell Death
3.2. Conditioned-Medium from LMSCs Improves Epithelial Repair Migratory and Proliferative Responses upon Injury
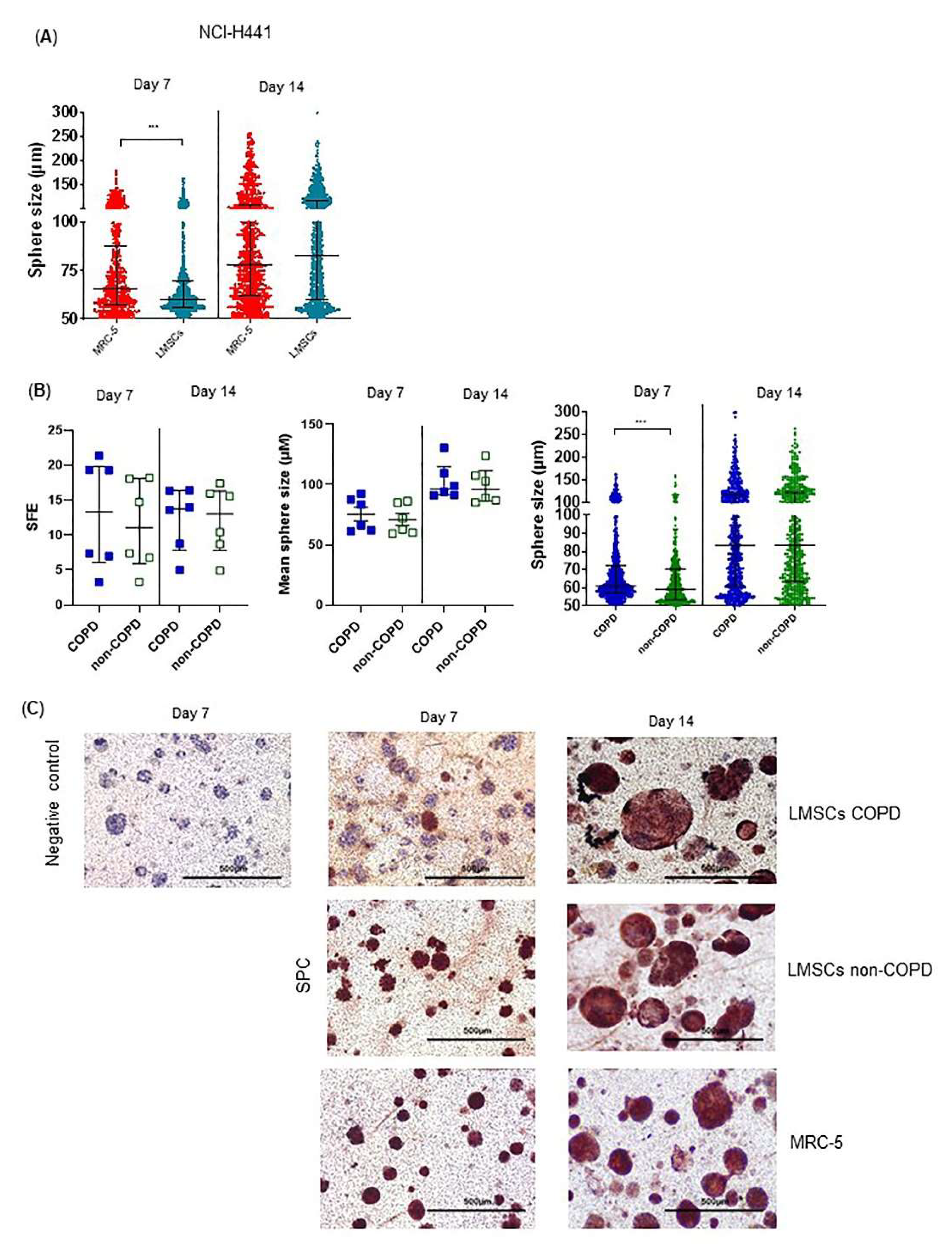
3.3. Regenerative Paracrine Effects of LMSCs in an Organoid Model
3.4. COPD-Derived Lung Cells Form Larger Alveolospheres Than Control-Derived Lung Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tuder, R.M.; Petrache, I. Pathogenesis of chronic obstructive pulmonary disease. J. Clin. Investig. 2012, 122, 2749–2755. [Google Scholar] [CrossRef]
- Kruk, D.M.L.W.; Heijink, I.H.; Slebos, D.J.; Timens, W.; Ten Hacken, N.H. Mesenchymal Stromal Cells to Regenerate Emphysema: On the Horizon? Respiration 2018, 96, 148–158. [Google Scholar] [CrossRef]
- Bernardo, M.E.; Fibbe, W.E. Mesenchymal stromal cells: Sensors and switchers of inflammation. Cell Stem Cell 2013, 13, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Weiss, A.R.R.; Dahlke, M.H. Immunomodulation by Mesenchymal Stem Cells (MSCs): Mechanisms of Action of Living, Apoptotic, and Dead MSCs. Front. Immunol. 2019, 10, 1191. [Google Scholar] [CrossRef] [Green Version]
- Reinders, M.E.; De Fijter, J.W.; Roelofs, H.; Bajema, I.M.; De Vries, D.K.; Schaapherder, A.F.; Claas, F.H.; Van Miert, P.P.; Roelen, D.L.; Van Kooten, C.; et al. Autologous Bone Marrow-Derived Mesenchymal Stromal Cells for the Treatment of Allograft Rejection After Renal Transplantation: Results of a Phase I Study. Stem Cells Transl. Med. 2013, 2, 107–111. [Google Scholar] [CrossRef]
- Rodrigo, S.F.; Van Ramshorst, J.; Hoogslag, G.E.; Boden, H.; Velders, M.A.; Cannegieter, S.C.; Roelofs, H.; Al Younis, I.; Dibbets-Schneider, P.; Fibbe, W.E.; et al. Intramyocardial Injection of Autologous Bone Marrow-Derived Ex Vivo Expanded Mesenchymal Stem Cells in Acute Myocardial Infarction Patients is Feasible and Safe up to 5 Years of Follow-up. J. Cardiovasc. Transl. Res. 2013, 6, 816–825. [Google Scholar] [CrossRef] [Green Version]
- Ingenito, E.P.; Tsai, L.; Murthy, S.; Tyagi, S.; Mazan, M.; Hoffman, A. Autologous Lung-Derived Mesenchymal Stem Cell Transplantation in Experimental Emphysema. Cell Transplant. 2012, 21, 175–189. [Google Scholar] [CrossRef]
- Rolandsson, S.; Sjöland, A.A.; Brune, J.C.; Li, H.; Kassem, M.; Mertens, F.; Westergren, A.; Eriksson, L.; Hansson, L.; Skog, I.; et al. Primary mesenchymal stem cells in human transplanted lungs are CD90/CD105 perivascularly located tissue-resident cells. BMJ Open Respir. Res. 2014, 1, e000027. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Kruk, D.M.L.W.; Wisman, M.; de Bruin, H.G.; Lodewijk, M.E.; Hof, D.J.; Borghuis, T.; Daamen, W.F.; van Kuppevelt, T.H.; Timens, W.; Burgess, J.K.; et al. Abnormalities in reparative function of lung-derived mesenchymal stromal cells in emphysema. Am. J. Physiol. Cell. Mol. Physiol. 2021, 320, L832–L844. [Google Scholar] [CrossRef]
- Weiss, D.J.; Casaburi, R.; Flannery, R.; LeRoux-Williams, M.; Tashkin, D.P. A placebo-controlled, randomized trial of mesenchymal stem cells in COPD. Chest 2013, 143, 1590–1598. [Google Scholar] [CrossRef] [Green Version]
- Stolk, J.; Broekman, W.; Mauad, T.; Zwaginga, J.; Roelofs, H.; Fibbe, W.; Oostendorp, J.; Bajema, I.; Versteegh, M.; Taube, C.; et al. A phase I study for intravenous autologous mesenchymal stromal cell administration to patients with severe emphysema. Qjm: Int. J. Med. 2016, 109, 331–336. [Google Scholar] [CrossRef] [Green Version]
- Weiss, D.J.; Segal, K.; Casaburi, R.; Hayes, J.; Tashkin, D. Effect of mesenchymal stromal cell infusions on lung function in COPD patients with high CRP levels. Respir. Res. 2021, 22, 142. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.W.; Tamatam, C.R.; Cho, I.S.; Toth, P.T.; Bargi, R.; Belvitch, P.; Lee, J.C.; Rehman, J.; Reddy, S.P.; Shin, J.-W. Inhibition of aberrant tissue remodelling by mesenchymal stromal cells singly coated with soft gels presenting defined chemomechanical cues. Nat. Biomed. Eng. 2021. [Google Scholar] [CrossRef] [PubMed]
- Enes, S.R.; Sjöland, A.A.; Skog, I.; Hansson, L.; Larsson, H.; Le Blanc, K.; Eriksson, L.; Bjermer, L.; Scheding, S.; Westergren-Thorsson, G. MSC from fetal and adult lungs possess lung-specific properties compared to bone marrow-derived MSC. Sci. Rep. 2016, 6, 29160. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, A.M.; Paxson, J.A.; Mazan, M.R.; Davis, A.M.; Tyagi, S.; Murthy, S.; Ingenito, E.P. Lung-Derived Mesenchymal Stromal Cell Post-Transplantation Survival, Persistence, Paracrine Expression, and Repair of Elastase-Injured Lung. Stem Cells Dev. 2011, 20, 1779–1792. [Google Scholar] [CrossRef] [PubMed]
- Kennelly, H.; Mahon, B.P.; English, K. Human mesenchymal stromal cells exert HGF dependent cytoprotective effects in a human relevant pre-clinical model of COPD. Sci. Rep. 2016, 6, 38207. [Google Scholar] [CrossRef]
- Wang, C.; De Mochel, N.S.R.; Christenson, S.A.; Cassandras, M.; Moon, R.; Brumwell, A.N.; Byrnes, L.E.; Li, A.; Yokosaki, Y.; Shan, P.; et al. Expansion of hedgehog disrupts mesenchymal identity and induces emphysema phenotype. J. Clin. Investig. 2018, 128, 4343–4358. [Google Scholar] [CrossRef]
- Volckaert, T.; Yuan, T.; Chao, C.-M.; Bell, H.; Sitaula, A.; Szimmtenings, L.; El Agha, E.; Chanda, D.; Majka, S.; Bellusci, S.; et al. Fgf10-Hippo Epithelial-Mesenchymal Crosstalk Maintains and Recruits Lung Basal Stem Cells. Dev. Cell 2017, 43, 48–59.e5. [Google Scholar] [CrossRef] [Green Version]
- Ramasamy, S.K.; Mailleux, A.A.; Gupte, V.V.; Mata, F.; Sala, F.G.; Veltmaat, J.M.; Del Moral, P.M.; De Langhe, S.; Parsa, S.; Kelly, L.K.; et al. Fgf10 dosage is critical for the amplification of epithelial cell progenitors and for the formation of multiple mesenchymal lineages during lung development. Dev. Biol. 2007, 307, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Ng-Blichfeldt, J.-P.; Schrik, A.; Kortekaas, R.K.; Noordhoek, J.A.; Heijink, I.H.; Hiemstra, P.S.; Stolk, J.; Königshoff, M.; Gosens, R. Retinoic acid signaling balances adult distal lung epithelial progenitor cell growth and differentiation. EBio Med. 2018, 36, 461–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heijink, I.H.; Brandenburg, S.M.; Noordhoek, J.A.; Postma, D.S.; Slebos, D.-J.; van Oosterhout, A.J.M. Characterisation of cell adhesion in airway epithelial cell types using electric cell-substrate impedance sensing. Eur. Respir. J. 2010, 35, 894–903. [Google Scholar] [CrossRef] [Green Version]
- Salomon, J.J.; Muchitsch, V.E.; Gausterer, J.C.; Schwagerus, E.; Huwer, H.; Daum, N.; Lehr, C.-M.; Ehrhardt, C. The Cell Line NCl-H441 Is a Useful in Vitro Model for Transport Studies of Human Distal Lung Epithelial Barrier. Mol. Pharm. 2014, 11, 995–1006. [Google Scholar] [CrossRef]
- Barkauskas, C.E.; Chung, M.-I.; Fioret, B.; Gao, X.; Katsura, H.; Hogan, B.L.M. Lung organoids: Current uses and future promise. Development 2017, 144, 986–997. [Google Scholar] [CrossRef] [Green Version]
- Chanda, D.; Rehan, M.; Smith, S.R.; Dsouza, K.G.; Wang, Y.; Bernard, K.; Kurundkar, D.; Memula, V.; Kyoko, K.; Mobley, J.A.; et al. Author response: Mesenchymal stromal cell aging impairs the self-organizing capacity of lung alveolar epithelial stem cells. Elife 2021, 10, e68049. [Google Scholar] [CrossRef]
- Leeman, K.T.; Pessina, P.; Lee, J.H.; Kim, C.F. Mesenchymal Stem Cells Increase Alveolar Differentiation in Lung Progenitor Organoid Cultures. Sci. Rep. 2019, 9, 6479. [Google Scholar] [CrossRef] [PubMed]
- Akram, K.M.; Samad, S.; Spiteri, M.A.; Forsyth, N.R. Mesenchymal stem cells promote alveolar epithelial cell wound repair in vitro through distinct migratory and paracrine mechanisms. Respir. Res. 2013, 14, 9. [Google Scholar] [CrossRef] [Green Version]
- Curley, G.; Hayes, M.; Ansari, B.; Shaw, G.; Ryan, A.; Barry, F.; O’Brien, T.; O′Toole, D.; Laffey, J.G. Mesenchymal stem cells enhance recovery and repair following ventilator-induced lung injury in the rat. Thorax 2012, 67, 496–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comer, D.M.; Kidney, J.C.; Ennis, M.; Elborn, J.S. Airway epithelial cell apoptosis and inflammation in COPD, smokers and nonsmokers. Eur. Respir. J. 2013, 41, 1058–1067. [Google Scholar] [CrossRef] [Green Version]
- Nita, I.; Hostettler, K.; Tamò, L.; Medova, M.; Bombaci, G.; Zhong, J.; Allam, R.; Zimmer, Y.; Roth, M.; Geiser, T.; et al. Hepatocyte growth factor secreted by bone marrow stem cell reduce ER stress and improves repair in alveolar epithelial II cells. Sci. Rep. 2017, 7, 41901. [Google Scholar] [CrossRef]
- Kanazawa, H.; Tochino, Y.; Asai, K.; Hirata, K. Simultaneous assessment of hepatocyte growth factor and vascular endothelial growth factor in epithelial lining fluid from patients with COPD. Chest 2014, 146, 1159–1165. [Google Scholar] [CrossRef]
- Maumus, M.; Jorgensen, C.; Noel, D. Mesenchymal stem cells in regenerative medicine applied to rheumatic diseases: Role of secretome and exosomes. Biochimie 2013, 95, 2229–2234. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, M.C.; Gorantla, V.; Makena, P.S.; Luellen, C.; Sinclair, S.E.; Schwingshackl, A.; Waters, C.M. Insulin-like growth factor-I stimulates differentiation of ATII cells to ATI-like cells through activation of Wnt5a. Am. J. Physiol. Cell. Mol. Physiol. 2013, 305, L222–L228. [Google Scholar] [CrossRef] [Green Version]
- Nabhan, A.N.; Brownfield, D.G.; Harbury, P.B.; Krasnow, M.A.; Desai, T.J. Single-cell Wnt signaling niches maintain stemness of alveolar type 2 cells. Science 2018, 359, 1118–1123. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Van Dijk, E.M.; Ng-Blichfeldt, J.-P.; Bos, I.S.T.; Ciminieri, C.; Königshoff, M.; Kistemaker, L.E.; Gosens, R. Mesenchymal WNT-5A/5B Signaling Represses Lung Alveolar Epithelial Progenitors. Cells 2019, 8, 1147. [Google Scholar] [CrossRef] [Green Version]
- Ng-Blichfeldt, J.P.; de Jong, T.; Kortekaas, R.K.; Wu, X.; Lindner, M.; Guryev, V.; Hiemstra, P.S.; Stolk, J.; Königshoff, M.; Gosens, R. TGF-beta activation impairs fibroblast ability to support adult lung epithelial progenitor cell organoid formation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 317, L14–L28. [Google Scholar] [CrossRef]
- Broekman, W.; Amatngalim, G.M.; de Mooji-Eijk, Y.; Oostendorp, J.; Roelofs, H.; Taube, C.; Stolk, J.; Hiemstra, P.S. TNF-alpha and IL-1beta-activated human mesenchymal stromal cells increase airway epithelial wound healing in vitro via activation of the epidermal growth factor receptor. Respir. Res. 2016, 17, 3. [Google Scholar] [CrossRef] [Green Version]
- Pouwels, S.D.; Hesse, L.; Wu, X.; Allam, V.S.R.R.; van Oldeniel, D.; Bhiekharie, L.J.; Phipps, S.; Oliver, B.G.G.; Gosens, R.; Sukkar, M.; et al. LL-37 and HMGB1 induce alveolar damage and reduce lung tissue regeneration via RAGE. Am. J. Physiol. Cell. Mol. Physiol. 2021, 321, L641–L652. [Google Scholar] [CrossRef]
- Ohmichi, H.; Matsumoto, K.; Nakamura, T. In vivo mitogenic action of HGF on lung epithelial cells: Pulmotrophic role in lung regeneration. Am. J. Physiol. 1996, 270, L1031–L1039. [Google Scholar] [CrossRef]
- Min, H.; Danilenko, D.; Scully, S.A.; Bolon, B.; Ring, B.D.; Tarpley, J.E.; De Rose, M.; Simonet, W.S. Fgf-10 is required for both limb and lung development and exhibits striking functional similarity to Drosophila branchless. Genes Dev. 1998, 12, 3156–3161. [Google Scholar] [CrossRef] [Green Version]
- Rabata, A.; Fedr, R.; Soucek, K.; Hampl, A.; Koledova, Z. 3D Cell Culture Models Demonstrate a Role for FGF and WNT Signaling in Regulation of Lung Epithelial Cell Fate and Morphogenesis. Front. Cell. Dev. Biol. 2020, 8, 574. [Google Scholar] [CrossRef]
- Ghosh, M.; Miller, Y.E.; Nakachi, I.; Kwon, J.B.; Barón, A.E.; Brantley, A.E.; Merrick, D.T.; Franklin, W.A.; Keith, R.L.; Vandivier, R.W. Exhaustion of Airway Basal Progenitor Cells in Early and Established Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2018, 197, 885–896. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-COPD Control (n = 12) | COPD GOLD III–IV (n = 12) | |
|---|---|---|
| Sex (M/F/NA) | 3/7/2 | 3/9/0 |
| Smoking (current/ex/never) | 2/5/3 | 0/12/0 |
| Age (years-range) | 66.2 (53–79) | 58.7 (51–68) |
| FEV1%Pred | 106.4 (80–131) | 25.0 (12–63) |
| FEV1/FVC | 75.6 (65.0–89.9) | 38.8 (21.9–89.2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kruk, D.M.L.W.; Wisman, M.; Noordhoek, J.A.; Nizamoglu, M.; Jonker, M.R.; de Bruin, H.G.; Arevalo Gomez, K.; ten Hacken, N.H.T.; Pouwels, S.D.; Heijink, I.H. Paracrine Regulation of Alveolar Epithelial Damage and Repair Responses by Human Lung-Resident Mesenchymal Stromal Cells. Cells 2021, 10, 2860. https://doi.org/10.3390/cells10112860
Kruk DMLW, Wisman M, Noordhoek JA, Nizamoglu M, Jonker MR, de Bruin HG, Arevalo Gomez K, ten Hacken NHT, Pouwels SD, Heijink IH. Paracrine Regulation of Alveolar Epithelial Damage and Repair Responses by Human Lung-Resident Mesenchymal Stromal Cells. Cells. 2021; 10(11):2860. https://doi.org/10.3390/cells10112860
Chicago/Turabian StyleKruk, Dennis M. L. W., Marissa Wisman, Jacobien A. Noordhoek, Mehmet Nizamoglu, Marnix R. Jonker, Harold G. de Bruin, Karla Arevalo Gomez, Nick H. T. ten Hacken, Simon D. Pouwels, and Irene H. Heijink. 2021. "Paracrine Regulation of Alveolar Epithelial Damage and Repair Responses by Human Lung-Resident Mesenchymal Stromal Cells" Cells 10, no. 11: 2860. https://doi.org/10.3390/cells10112860